
Subfamily Anischiinae (Coleoptera: Eucnemidae) in Early Cretaceous of Northeast China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
- Systematic palaeontology
- Class: Insecta Linnaeus, 1758;
- Order: Coleoptera Linnaeus, 1758;
- Superfamily: Elateroidea Leach, 1815;
- Family: Eucnemidae Eschscholtz, 1829;
- Subfamily: Anischiinae Fleutiaux, 1936;
3.1. Diagnosis
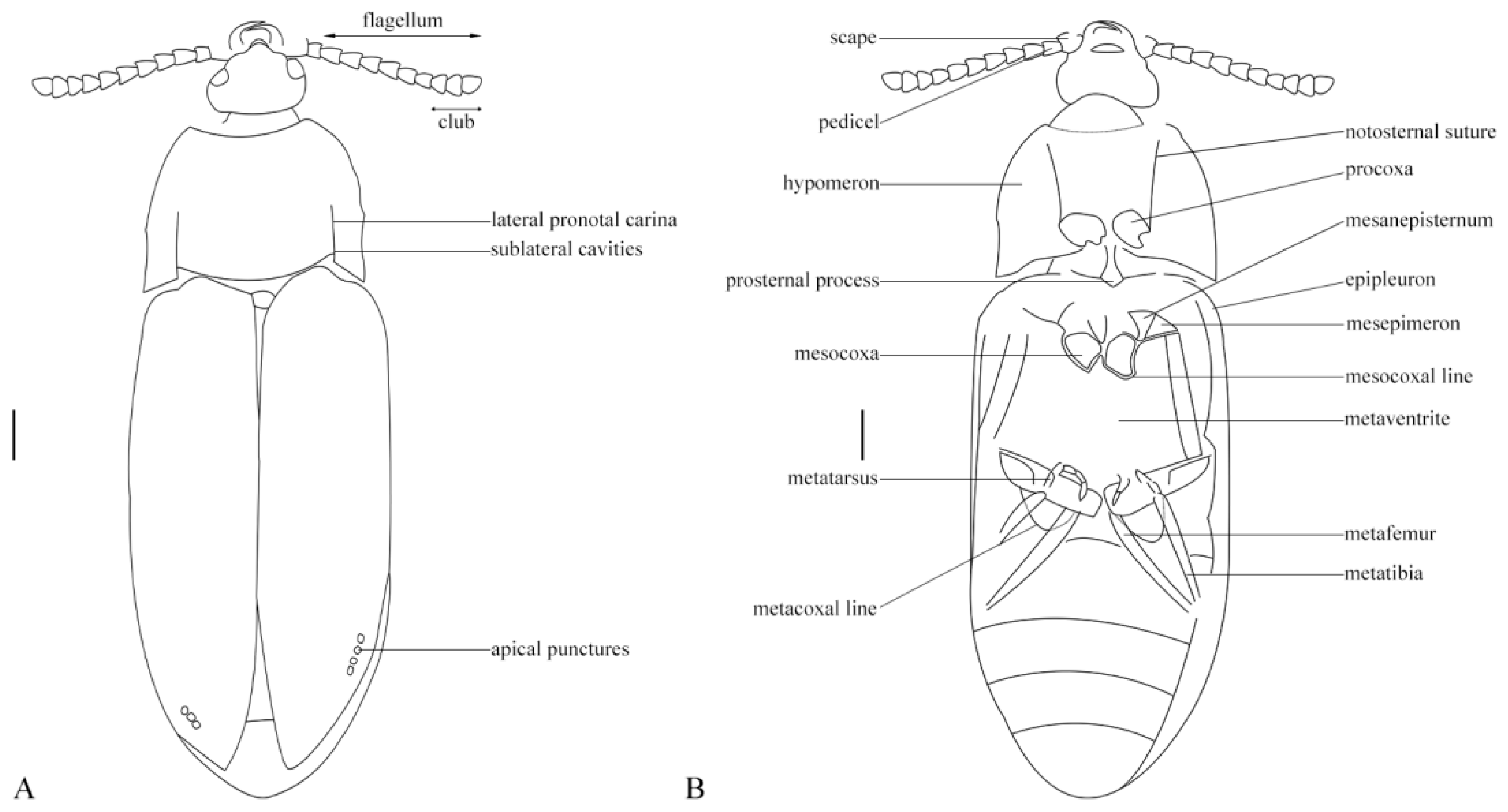
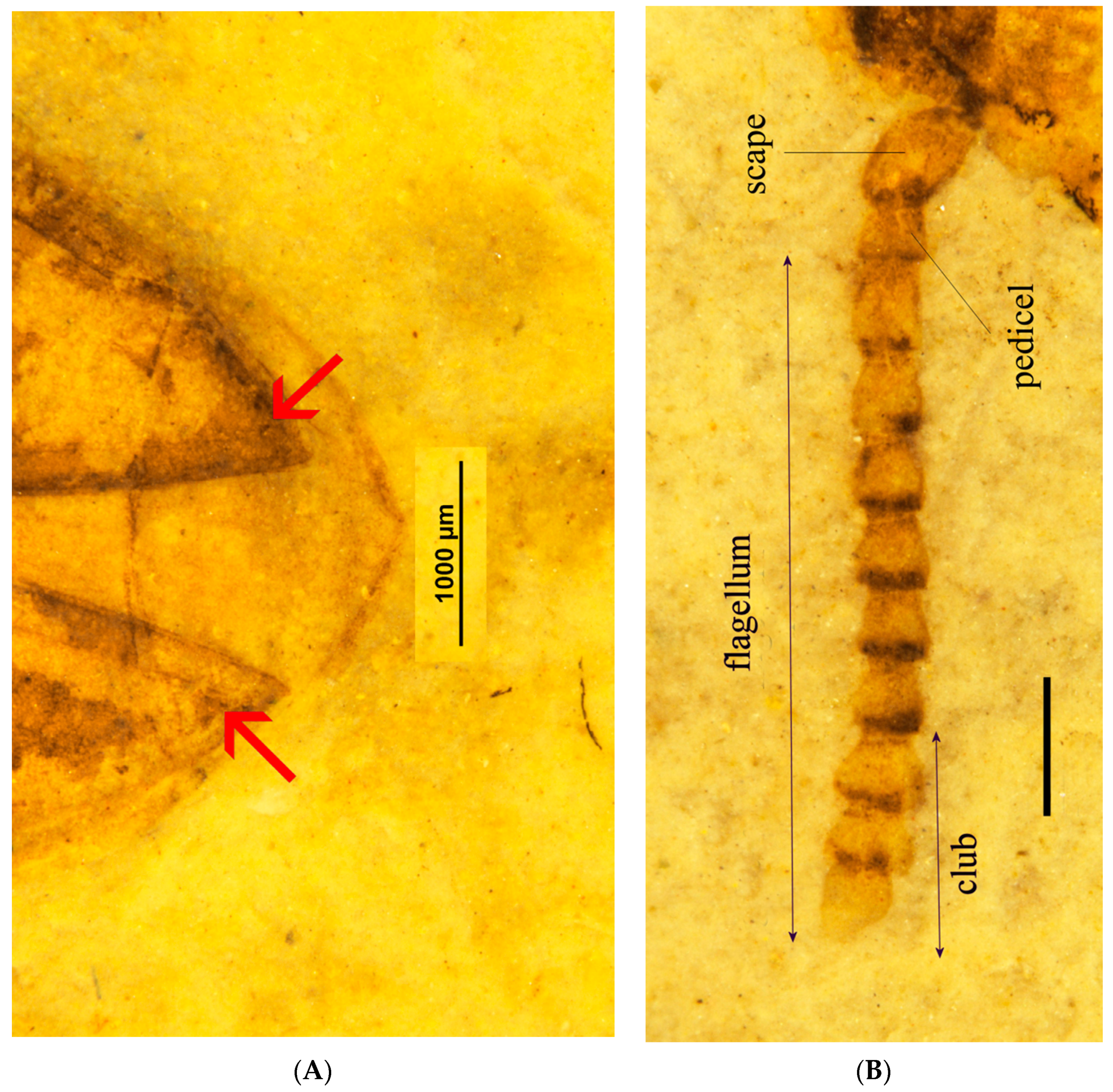
3.2. Description
3.3. Diagnosis
3.4. Description
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lawrence, J.F.; Bocak, L.; Bocakova, M.; Beutel, R.G.; Muona, J. 4. Elateroidea. In Handbook of Zoology—Arthropoda: Insecta—Coleoptera, Beetles; Beutel, R.G., Lawrence, J.F., Leschen, R.A.B., Eds.; De Gruyter: Berlin, Germany, 2010; Volume 2, pp. 35–38. [Google Scholar]
- Costa, C.; Lawrence, J.F.; Rosa, S.P. 4.7. Elateridae Leach. In Handbook of Zoology—Arthropoda: Insecta—Coleoptera, Beetles; Beutel, R.G., Lawrence, J.F., Leschen, R.A.B., Eds.; De Gruyter: Berlin, Germany, 2010; Volume 2, pp. 75–103. [Google Scholar]
- Kundrata, R.; Packova, G.; Hoffmannova, J. Fossil Genera in Elateridae (Insecta, Coleoptera): A Triassic Origin and Jurassic Diversification. Insects 2020, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Muona, J. 4.5. Eucnemidae Eschscholtz. In Handbook of Zoology—Arthropoda: Insecta—Coleoptera, Beetles; Beutel, R.G., Lawrence, J.F., Leschen, R.A.B., Eds.; De Gruyter: Berlin, Germany, 2010; Volume 2, pp. 61–68. [Google Scholar]
- Muona, J. Eucnemidae and Throscidae in Baltic Amber. Ent. Blätter and Coleoptera 1993, 89, 15–45. [Google Scholar]
- Chang, H.; Muona, J.; Pu, H.; Xu, L.; Wang, C.; Teräväinen, M.; Ren, D.; Yang, Q.; Zhang, X.; Jia, S. Chinese Cretaceous larva exposes a southern Californian living fossil (Insecta, Coleoptera, Eucnemidae). Cladistics 2016, 32, 211–214. [Google Scholar] [CrossRef]
- Oberprieler, R.; Ashman, L.; Friese, M.; Slipinski, A. The first elateroid beetles (Coleoptera: Polyphaga: Elateroidea) from the Upper Jurassic of Australia. Zootaxa 2016, 4147, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Otto, R.; Gruber, J. Eucnemid larvae of the Nearctic region. Part VI. Descriptions of the fifth instar and prepupal larval stages of Stethon pectorosus LeConte (Coleoptera: Eucnemidae) with notes on their biology. Insecta Mundi. 2019, 474, 1–11. [Google Scholar]
- Li, Y.D.; Tihelka, E.; Liu, Z.H.; Huang, D.; Cai, C.Y. Muonabuntor gen. nov., a new genus of false click beetles from mid-Cretaceous Burmese amber (Coleoptera: Elateroidea: Eucnemidae). Palaeoentomology 2020, 3, 399–406. [Google Scholar] [CrossRef]
- Muona, J.; Chang, H.; Ren, D. The Clicking Elateroidea from Chinese Mesozoic Deposits (Insecta, Coleoptera). Insects 2020, 11, 875. [Google Scholar] [CrossRef]
- Muona, J. The late Cretaceous amber Eucnemidae fossils from Myanmar. Ent. Blätter Coleopt. 2020, 116, 27–55. [Google Scholar]
- Gao, T.; Shih, C.; Ren, D. Behaviors and interactions of insects in ecosystems of Mid-Mesozoic northeastern China. Ann. Rev. Entomol. 2020, 66, 337–354. [Google Scholar] [CrossRef]
- Ren, D.; Lu, L.W.; Ji, S.A.; Guo, Z.G. Faunae and Stratigraphy of Jurassic-Cretaceous in Beijing and the Adjacent Areas; Seismic Publishing House: Beijing, China, 1995; 223p. (In Chinese) [Google Scholar]
- Ren, D. Flower-associated Brachycera flies as fossil evidence for Jurassic angiosperm origins. Science 1998, 280, 85–88. [Google Scholar] [CrossRef]
- Ren, D.; Shih, C.; Gao, T.; Wang, Y.; Yao, Y. Rhythms of Insect Evolution—Evidence from the Jurassic and Cretaceous in Northern China; John Wiley et Sons: Hoboken, NJ, USA, 2019; 710p. [Google Scholar]
- Ding, Q.H.; Zhang, L.D.; Gou, S.Z.; Zhang, C.J.; Peng, Y.D.; Jia, B.; Chen, S.W.; Xing, D.H. The stratigraphic sequence and fossil bearing horizon of the Yixian Formation in western Liaoning, China. Geol. Resour. 2001, 10, 193–198, (In Chinese with English Abstract). [Google Scholar]
- Gao, T.; Shih, C.; Rasnitsyn, P.; Xu, X.; Wang, S.; Ren, D. New Transitional Fleas from China Highlighting Diversity of Early Cretaceous Ectoparasitic Insects. Curr. Biol. 2013, 23, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Labandeira, C.; Ding, Q.; Meng, Q.; Ren, D. Exploiting nondietary resources in deep time: Patterns of oviposition on mid-Mesozoic plants from Northeastern China. Int. J. Sci. 2019, 180, 411–457. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Shih, C.; Rasnitsyn, P.A.; Yao, J.; Ren, D.; Gao, T. A new sawfly of Megalodontesidae (Insecta, Hymenoptera, Pamphilioidea) with pectinate antennae from the Early Cretaceous of China. ZooKeys 2019, 893, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.H.; Sha, J.G.; Fürsich, F.T.; Wang, Y.; Zhang, X.; Yao, X. Dynamics of the lacustrine fauna from the Early Cretaceous Yixian Formation, China: Implications of volcanic and climatic factors. Lethaia 2012, 45, 299–314. [Google Scholar] [CrossRef]
- Pan, Y.H.; Sha, J.G.; Zhou, Z.H.; Fürsich, F.T. The Jehol Biota: Definition and distribution of exceptionally preserved relicts of a continental Early Cretaceous ecosystem. Cretac. Res. 2013, 44, 30–38. [Google Scholar] [CrossRef]
- Calder, A. Click Beetles: Genera of the Australian Elateridae (Coleoptera)—Monographs of Invertebrate Taxonomy; CSIRO: Canberra, ACT, Australia, 1996; Volume 2, 401p. [Google Scholar]
- Douglas, H. Phylogenetic relationships of Elateridae inferred from adult morphology, with special reference to the postion of Cardiophorinae. Zootaxa 2011, 2900, 1–45. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Muona, J.; Teräväinen, M.; Ståhls, G.; Vahtera, V. Anischia, Perothops and the phylogeny of Elateroidea (Coleoptera: Elateriformia). Insect Syst. Evol. 2007, 38, 205–239. [Google Scholar] [CrossRef]
- Kundrata, R.; Bocakova, M.; Bocak, L. The comprehensive phylogeny of the superfamily Elateroidea (Coleoptera: Elateriformia). Mol. Phylogenetics Evol. 2014, 76, 162–171. [Google Scholar] [CrossRef]
- Muona, J.; Teräväinen, M. A Re-evaluation of the Eucnemidae larval characters (Coleoptera). Papéis Avulsos Zool. 2020, 60, 28. [Google Scholar] [CrossRef]
- Otto, R.; Young, D. Description of the Larva of Schizophilus subrufus (Randall) (Coleoptera: Eucnemidae: Pseudomeninae), with Notes on Its Natural History. Coleopt. Bull. 1998, 52, 306–312. [Google Scholar]
- Muona, J. Review of the phylogeny, classification and biology of the family Eucnemidae (Coleoptera). Scand. Entomol. 1993, 44, 1–133. [Google Scholar]
- Muona, J.; Teräväinen, M. Notes on the biology and morphology of false click-beetle larvae (Coleoptera: Eucnemidae). Coleopt. Bull. 2008, 62, 475–479. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Chang, H.; Muona, J.; Zhao, Y.; Ren, D. Subfamily Anischiinae (Coleoptera: Eucnemidae) in Early Cretaceous of Northeast China. Insects 2021, 12, 105. https://doi.org/10.3390/insects12020105
Li H, Chang H, Muona J, Zhao Y, Ren D. Subfamily Anischiinae (Coleoptera: Eucnemidae) in Early Cretaceous of Northeast China. Insects. 2021; 12(2):105. https://doi.org/10.3390/insects12020105
Chicago/Turabian StyleLi, Haolun, Huali Chang, Jyrki Muona, Yanchen Zhao, and Dong Ren. 2021. "Subfamily Anischiinae (Coleoptera: Eucnemidae) in Early Cretaceous of Northeast China" Insects 12, no. 2: 105. https://doi.org/10.3390/insects12020105
APA StyleLi, H., Chang, H., Muona, J., Zhao, Y., & Ren, D. (2021). Subfamily Anischiinae (Coleoptera: Eucnemidae) in Early Cretaceous of Northeast China. Insects, 12(2), 105. https://doi.org/10.3390/insects12020105