
The Automatic Classification of Pyriproxyfen-Affected Mosquito Ovaries


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
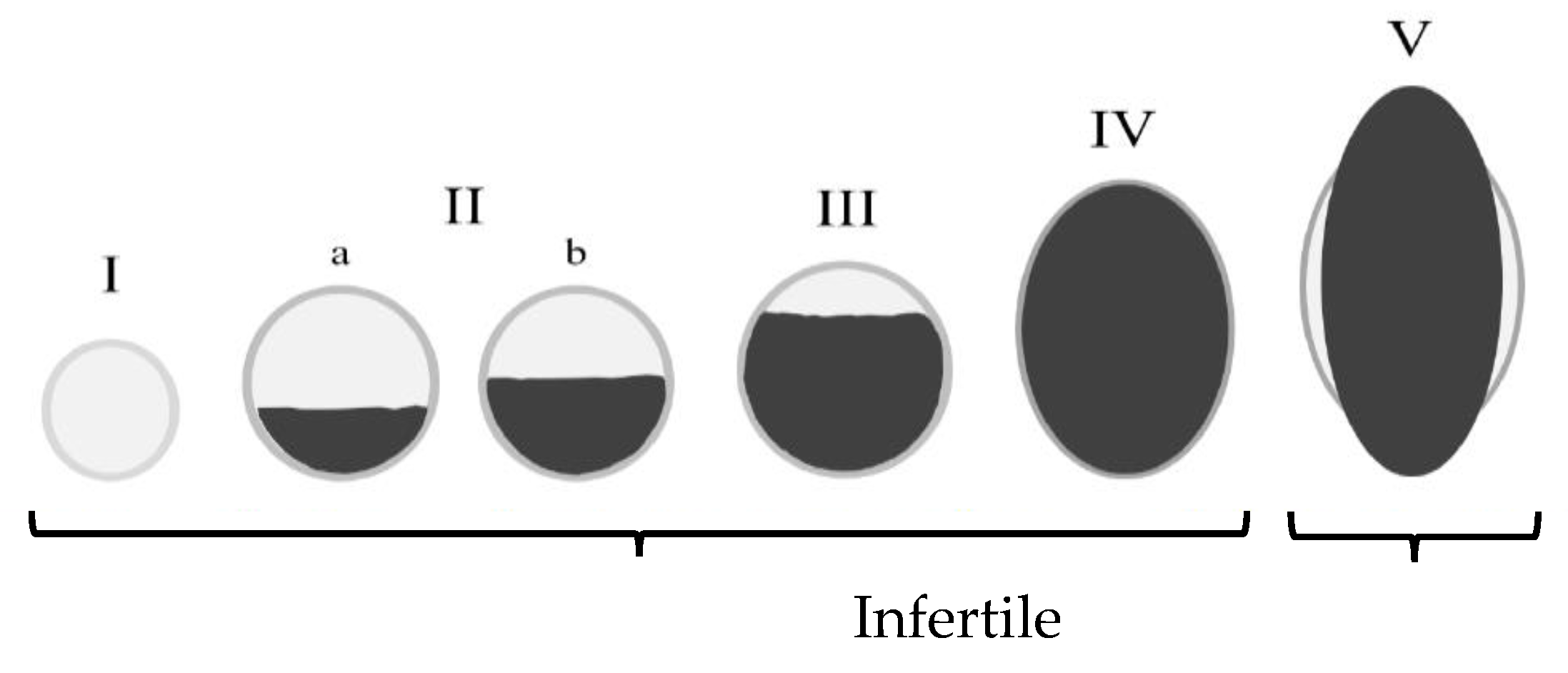
2.1. Image Dataset
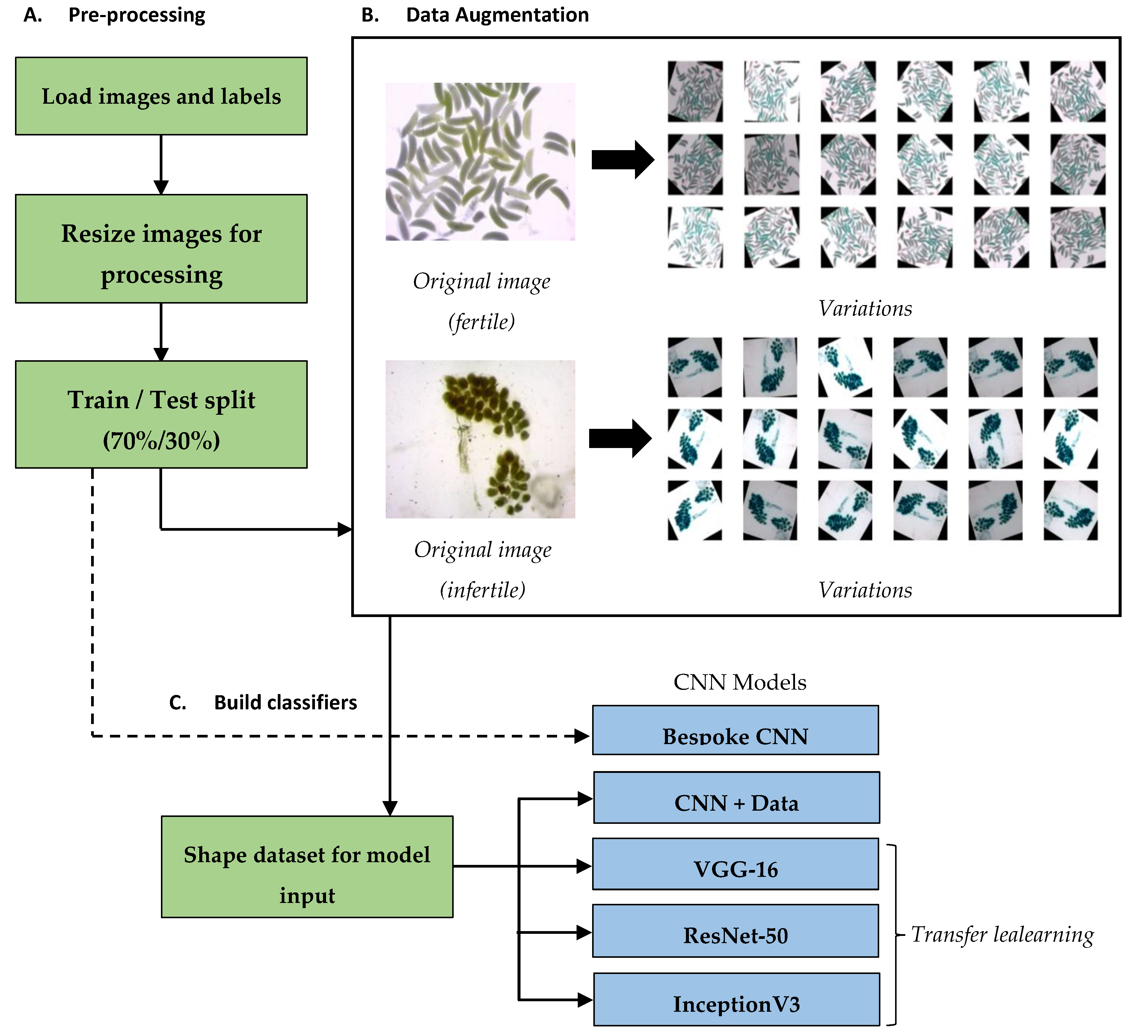
2.2. Pre-Processing and Train/Test Split
2.3. Data Augmentation
2.4. Analysis
2.5. Resources and Requirements
3. Results
3.1. Classification Accuracy
3.2. Classification Speed
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Global Report on Insecticide Resistance in Malaria Vectors: 2010–2016; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- World Health Organization. Implications of Insecticide Resistance for Malaria Vector Control; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitos: A Worsening Situation That Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- Ngufor, C.; N’guessan, R.; Fagbohoun, J.; Odjo, A.; Malone, D.; Akogbeto, M.; Rowland, M. Olyset Duo® (a Pyriproxyfen and Permethrin Mixture Net): An Experimental Hut Trial against Pyrethroid Resistant Anopheles Gambiae and Culex Quinquefasciatus in Southern Benin. PLoS ONE 2014, 3, e93603. [Google Scholar] [CrossRef]
- Tiono, A.B.; Ouédraogo, A.; Ouattara, A.; Bougouma, E.C.; Coulibaly, S.; Diarra, A.; Faragher, B.; Guelbeogo, W.M.; Grisales, N.; Ouédraogo, Z.A.; et al. Efficacy of Olyset Duo, a Bednet Containing Pyriproxyfen and Permethrin, versus a Permethrin-Only Net against Clinical Malaria in an Area with Highly Pyrethroid-Resistant Vectors in Rural Burkina Faso: A Cluster-Randomised Controlled Trial. Lancet 2018, 18, 569–580. [Google Scholar] [CrossRef]
- Toé, K.H.; Mechan, F.; Tangena, J.A.; Morris, M.; Solino, J.; Tchicaya, E.F.S.; Traoré, A.; Ismail, H.; Maas, J.; Lissenden, N.; et al. Assessing the Impact of the Addition of Pyriproxyfen on the Durability of Permethrin-Treated Bed Nets in Burkina Faso: A Compound-Randomized Controlled Trial. Malar. J. 2019, 18, 383. [Google Scholar] [CrossRef] [Green Version]
- Ngufor, C.; Agbevo, A.; Fagbohoun, J.; Fongnikin, A.; Rowland, M. Efficacy of Royal Guard, a New Alpha-Cypermethrin and Pyriproxyfen Treated Mosquito Net, against Pyrethroid-Resistant Malaria Vectors. Sci. Rep. 2020, 22, 12777. [Google Scholar] [CrossRef]
- Ohashi, K.; Nakada, K.; Ishiwateri, T.; Miyaguchi, J.; Shono, Y.; Lucas, J.R.; Mito, N. Efficacy of Pyriproxyfen-Treated Nets in Sterilizing and Shortening the Longevity of Anopheles Gambiae (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Kawada, H.; Dida, G.O.; Ohash, K.; Kawashima, E.; Sonye, G.; Njenga, S.M.; Mwandawiro, C.; Minakawa, N. A Small-Scale Field Trial of Pyriproxyfen-Impregnated Bed Nets against Pyrethroid-Resistant Anopheles Gambiae s.s. in Western Kenya. PLoS ONE 2014, 9, e111195. [Google Scholar] [CrossRef] [Green Version]
- Ngufor, C.; Fagbohoun, J.; Todjinou, D.; Odjo, A.; Malone, D.; Ismail, H.; Akogbeto, M.; Rowland, M. Efficacy of the Olyset Duo Net against Insecticide-Resistant Mosquito Vectors of Malaria. Sci. Transl. Med. 2016, 8, 356ra121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisales, N.; Lees, R.S.; Maas, J.; Morgan, J.C.; Wangrawa, D.W.; Guelbeogo, W.M.; N’Fale, S.; Lindsay, S.W.; McCall, P.J.; Ranson, H. Pyriproxyfen-Treated Bed Nets Reduce Reproductive Fitness and Longevity of Pyrethroid-Resistant Anopheles Gambiae under Laboratory and Field Conditions. Malar. J. 2021, 20, 273. [Google Scholar] [CrossRef] [PubMed]
- Koama, B.; Namountougou, M.; Sanou, R.; Ouattara, A.; Dabiré, R.K.; Malone, D.; Diabaté, A. The Sterilizing Effect of Pyriproxyfen on the Malaria Vector Anopheles Gambiae: Physiological Impact on Ovaries Development. Malar. J. 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, K.; Dhiman, S.; Acharya, B.; Ghorpade, R.R.; Sukumaran, D. Pyriproxyfen Treated Surface Exposure Exhibits Reproductive Disruption in Dengue Vector Aedes Aegypti. PLoS Negl. Trop. Dis. 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christophers, S.R. The Development of the Egg Follicle in Anophelines. Paludism 1911, 2, 73–88. [Google Scholar]
- Albawi, S.; Mohammed, T.A.; Al-Zawi, S. Understanding of a Convolutional Neural Network. In Proceedings of the International Conference on Engineering and Technology (ICET), Antalya, Turkey, 21–23 August 2017; pp. 1–6. [Google Scholar] [CrossRef]
- Rawat, W.; Wang, Z. Deep Convolutional Neural Networks for Image Classification: A Comprehensive Review. Neural Comput. 2017, 29, 2352–2449. [Google Scholar] [CrossRef]
- Hopfield, J.J. Neural Networks and Physical Systems with Emergent Collective Computational Abilities. Proc. Natl. Acad. Sci. USA 1982, 79, 2554–2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivas, S.; Sarvadevabhatla, R.K.; Mopuri, K.R.; Prabhu, N.; Kruthiventi, S.S.S.; Babu, R.V. A Taxonomy of Deep Convolutional Neural Nets for Computer Vision. Front. Robot. AI 2016, 2, 36. [Google Scholar] [CrossRef]
- Zeiler, M.D.; Fergus, R. Visualizing and Understanding Convolutional Networks. In Computer Vision—ECCV 2014; Lecture Notes in Computer Science; Springer: Zurich, Switzerland, 2014; Volume 8689, pp. 818–833. [Google Scholar]
- Venkatesan, R.; Li, B. Convolutional Neural Networks in Visual Computing: A Concise Guide; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Chen, X.; Lin, X. Big Data Deep Learning: Challenges and Perspectives. IEEE Access 2014, 2, 514–525. [Google Scholar] [CrossRef]
- Shorten, C.; Khoshgoftaar, T.M. A Survey on Image Data Augmentation for Deep Learning. J. Big Data 2019, 6, 60. [Google Scholar] [CrossRef]
- Weiss, K.; Khoshgoftaar, T.M.; Wang, D. A Survey of Transfer Learning. J. Big Data 2016, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Manzo, M.; Pellino, S. Voting in Transfer Learning System for Ground-Based Cloud Classification. Mach. Learn. Knowl. Extr. 2021, 3, 542–553. [Google Scholar] [CrossRef]
- Simonyan, K.; Zisserman, A. Very Deep Convolutional Networks for Large-Scale Image Recognition. arXiv 2014, arXiv:1409.1556. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep Residual Learning for Image Recognition. In Proceedings of the 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Szegedy, C.; Vanhoucke, V.; Ioffe, S.; Shlens, J.; Woja, Z. Rethinking the Inception Architecture for Computer Vision. In Proceedings of the 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016; pp. 2818–2826. [Google Scholar]
- Szegedy, C.; Liu, W.; Jia, Y.; Reed, S.; Anguleox, D.; Erhan, D.; Vanhoucke, V.; Rabinovich, A. Going Deeper with Convolutions. In Proceedings of the 2015 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Boston, MA, USA, 7–12 June 2015; pp. 1–9. [Google Scholar]
- Liu, Z.; Gao, J.; Yang, G.; Zhang, H.; He, Y. Localization and Classification of Paddy Field Pests Using a Saliency Map and Deep Convolutional Neural Network. Sci. Rep. 2016, 6, 20410. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Ma, M.; Zhang, Z.; Zhang, P. Hybrid Deep Learning for Automated Lepidopteran Insect Image Classification. Orient. Insects 2017, 51, 79–91. [Google Scholar] [CrossRef]
- Couret, J.; Moreira, D.C.; Bernier, D.; Loberti, A.M.; Dotson, E.M.; Alvarez, M. Delimiting Cryptic Morphological Variation among Human Malaria Vector Species Using Convolutional Neural Networks. PLoS Negl. Trop. Dis. 2020, 17. [Google Scholar] [CrossRef]
- Minakshi, M.; Bharti, P.; Bhuiyan, T.; Kariev, S.; Chellappan, S. A Framework Based on Deep Neural Networks to Extract Anatomy of Mosquitoes from Images. Sci. Rep. 2020, 10, 13059. [Google Scholar] [CrossRef]
- Motta, D.; Santos, A.Á.B.; Machado, B.A.S.; Ribeiro-Filho, O.G.V.; Camargo, L.O.A.; Valdenegro-Toro, M.A.; Kirhner, F.; Badaró, R. Optimization of Convolutional Neural Network Hyperparameters for Automatic Classification of Adult Mosquitoes. PLoS ONE 2020, 15, e0234959. [Google Scholar] [CrossRef]
- Park, J.; Kim, D.I.; Choi, B.; Kang, W.; Kwon, H.W. Classification and Morphological Analysis of Vector Mosquitoes Using Deep Convolutional Neural Networks. Sci. Rep. 2020, 10, 1012. [Google Scholar] [CrossRef] [PubMed]
- Mains, J.W.; Mercer, D.R.; Dobson, S.L. Digital Image Analysis to Estimate Numbers of Aedes Eggs Oviposited in Containers. J. Am. Mosq. Control. Assoc. 2008, 24, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Dembo, E.; Ogboi, J.; Abay, S.; Lupidi, G.; Dahiya, N.; Habluetzal, A.; Lucantoni, L. A User Friendly Method to Assess Anopheles Stephensi (Diptera: Culicidae) Vector Fitness: Fecundity. J. Med. Entomol. 2014, 51, 831–836. [Google Scholar] [CrossRef]
- Gaburro, J.; Duchemin, J.B.; Paradkar, P.N.; Nahavandi, S.; Bhatti, A. Assessment of ICount software, a precise and fast egg counting tool for the mosquito vector Aedes aegypti. Parasites Vectors 2016, 9, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Brogdon, W.G.; Chan, A. A Guideline for Evaluating Insecticide Resistance in Vectors Using the CDC Bottle Bioassay; Centers for Disease Control and Prevention: Altlanta, GA, USA, 2012. [Google Scholar]
- Géron, A. Hands-On Machine Learning with Scikit-Learn and TensorFlow; O’Reilly: Farnham, UK, 2017. [Google Scholar]
- Versaci, M.; Angiulli, G.; di Barba, P.; Morabito, F.C. Joint Use of Eddy Current Imaging and Fuzzy Similarities to Assess the Integrity of Steel Plates. Open Phys. 2020, 18, 230–240. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Dataset | Source | Strain | Image Count | Fertile | Infertile |
|---|---|---|---|---|---|
| 1 | Cove, Benin | An. gambiae s.l. | 124 | 79 | 45 |
| 2 | Insectary colony | An. gambiae Akron | 187 | 43 | 144 |
| 3 | Mwanza, Tanzania | An. funestus s.l. | 125 | 67 | 58 |
| 4 | Mwanza, Tanzania | An. gambiae s.l. | 88 | 38 | 50 |
| Total | 524 | 227 | 297 | ||
| Architecture | Accuracy 1 | Recall (Fer) 2 | Recall (Inf) 3 | Precision (Fer) 4 | Precision (Inf) 5 | Speed 6 |
|---|---|---|---|---|---|---|
| Bespoke CNN | 0.777 | 0.951 | 0.592 | 0.918 | 0.713 | 28.1 s |
| CNN + data Augmentation | 0.815 | 0.901 | 0.724 | 0.873 | 0.777 | 28.7 s |
| VGG-16 | 0.885 | 0.901 | 0.868 | 0.892 | 0.880 | 41.7 s |
| ResNet-50 | 0.943 | 0.951 | 0.934 | 0.947 | 0.939 | 38.5 s |
| InceptionV3 | 0.803 | 0.716 | 0.895 | 0.747 | 0.879 | 36.5 s |
| n = 157 1 | Predicted Infertile | Predicted Fertile | |
|---|---|---|---|
| Actual Infertile | 77 | 4 | 81 |
| Actual Fertile | 5 | 71 | 76 |
| 82 | 75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fowler, M.T.; Lees, R.S.; Fagbohoun, J.; Matowo, N.S.; Ngufor, C.; Protopopoff, N.; Spiers, A. The Automatic Classification of Pyriproxyfen-Affected Mosquito Ovaries. Insects 2021, 12, 1134. https://doi.org/10.3390/insects12121134
Fowler MT, Lees RS, Fagbohoun J, Matowo NS, Ngufor C, Protopopoff N, Spiers A. The Automatic Classification of Pyriproxyfen-Affected Mosquito Ovaries. Insects. 2021; 12(12):1134. https://doi.org/10.3390/insects12121134
Chicago/Turabian StyleFowler, Mark T., Rosemary S. Lees, Josias Fagbohoun, Nancy S. Matowo, Corine Ngufor, Natacha Protopopoff, and Angus Spiers. 2021. "The Automatic Classification of Pyriproxyfen-Affected Mosquito Ovaries" Insects 12, no. 12: 1134. https://doi.org/10.3390/insects12121134
APA StyleFowler, M. T., Lees, R. S., Fagbohoun, J., Matowo, N. S., Ngufor, C., Protopopoff, N., & Spiers, A. (2021). The Automatic Classification of Pyriproxyfen-Affected Mosquito Ovaries. Insects, 12(12), 1134. https://doi.org/10.3390/insects12121134