
Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Beauveria Bassiana Strains
2.2. Preparation of Fungal Suspension
2.3. Bioassays
2.4. Statistical Analysis
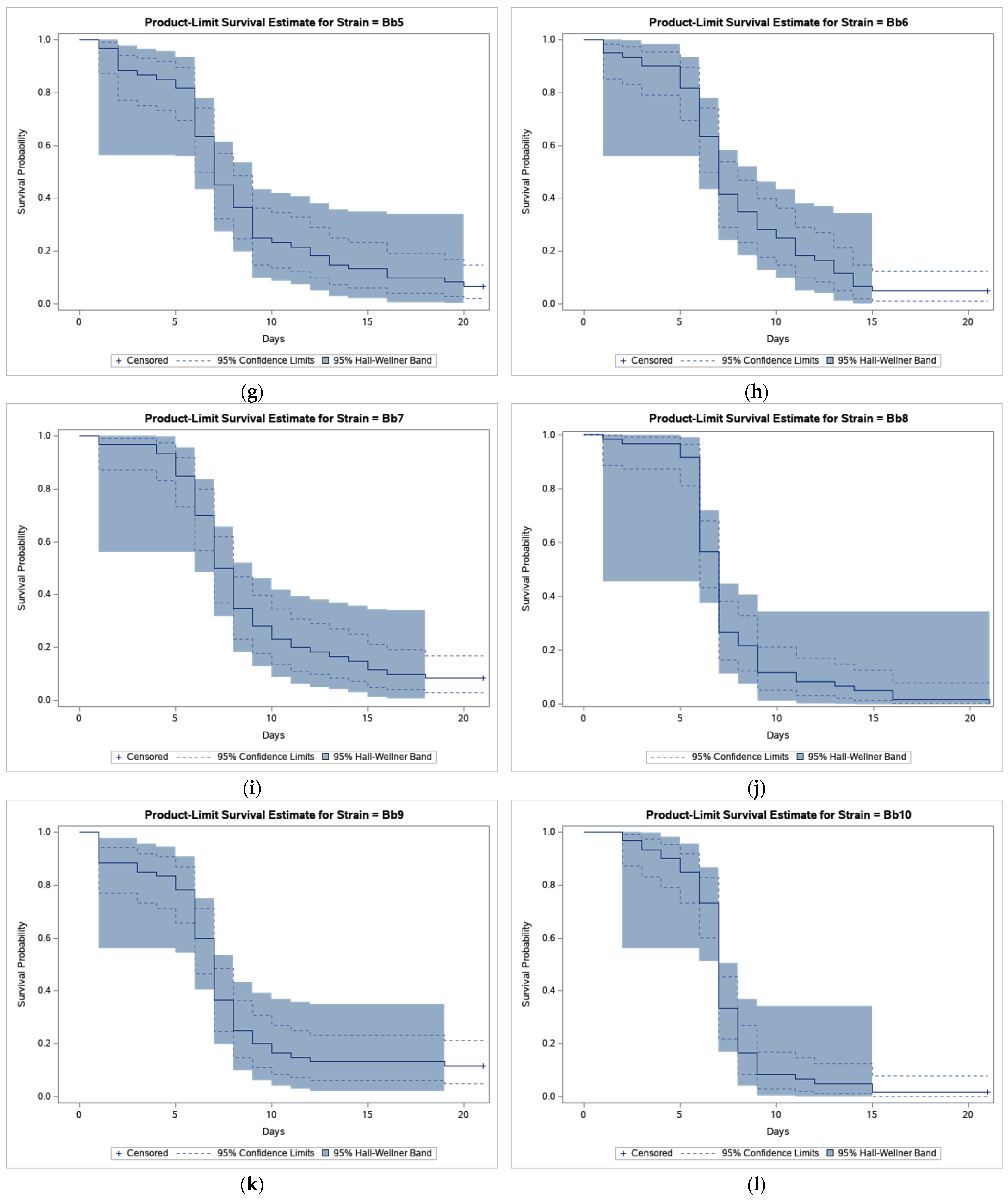
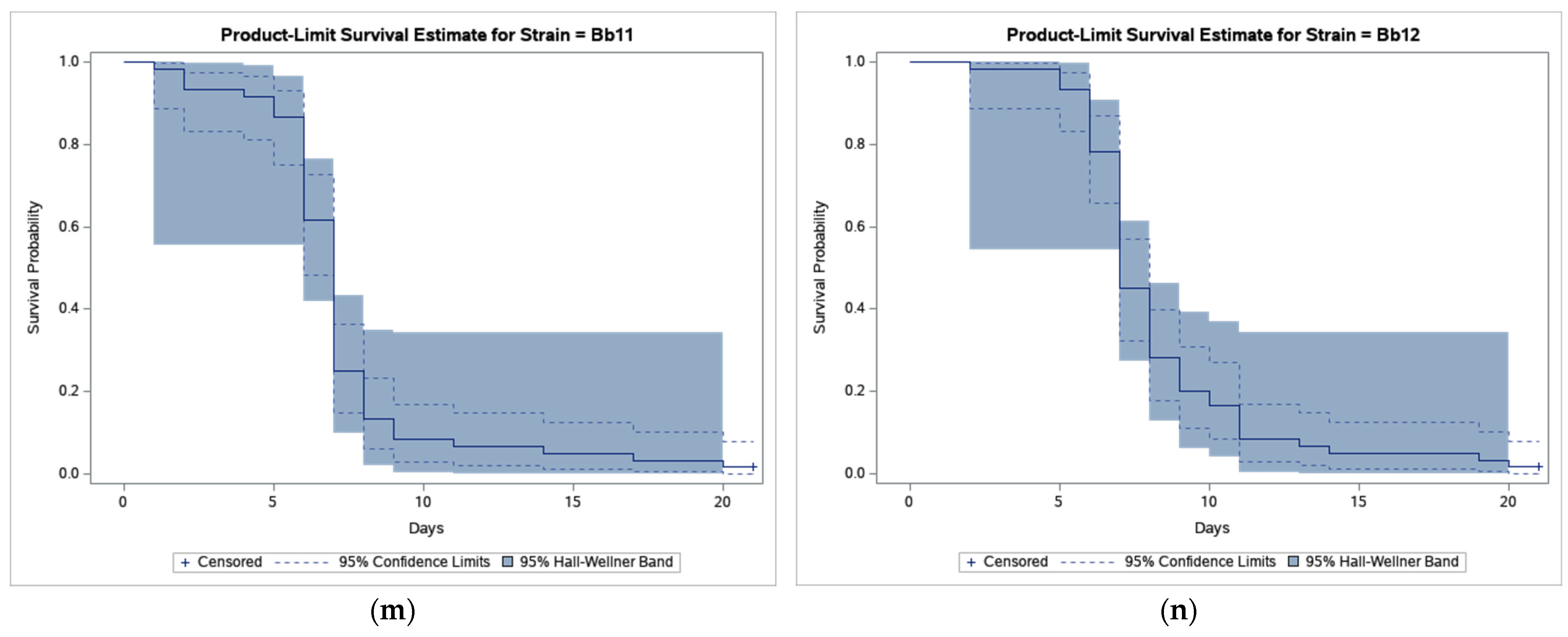
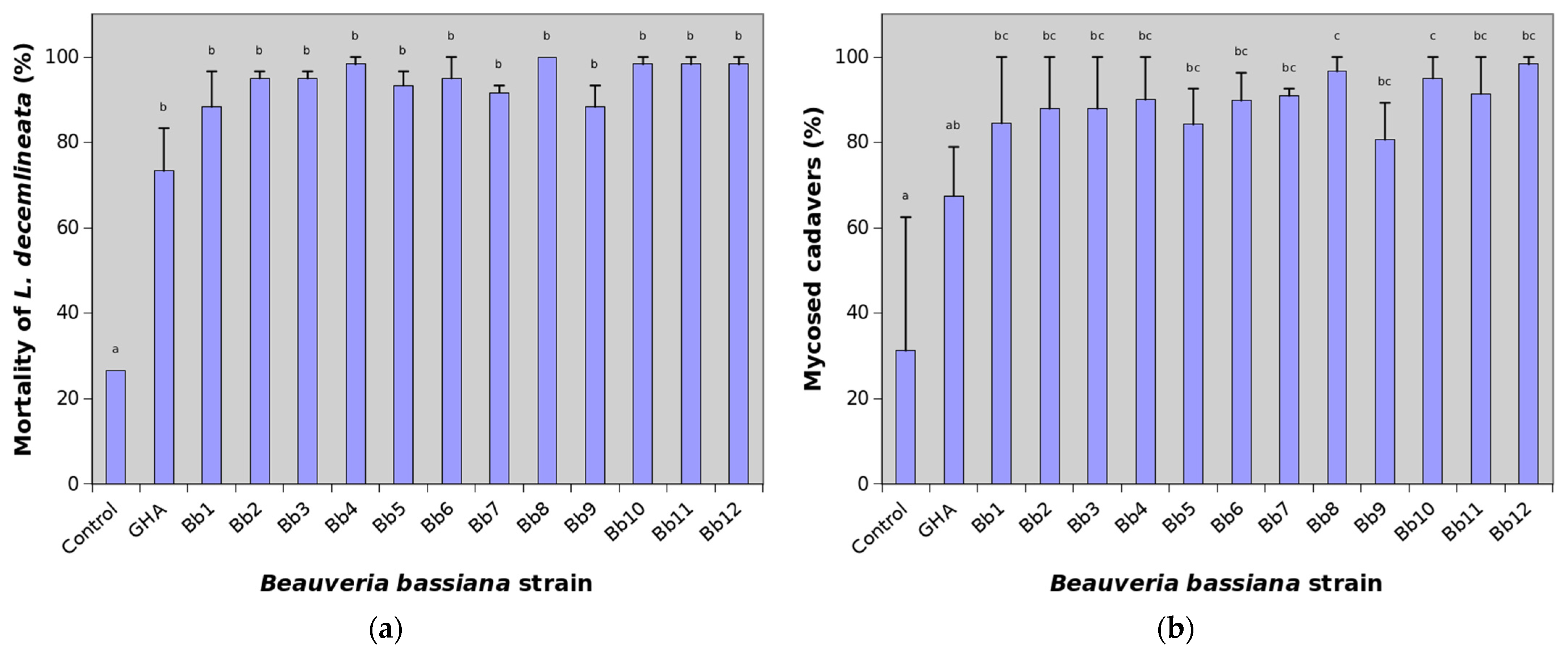
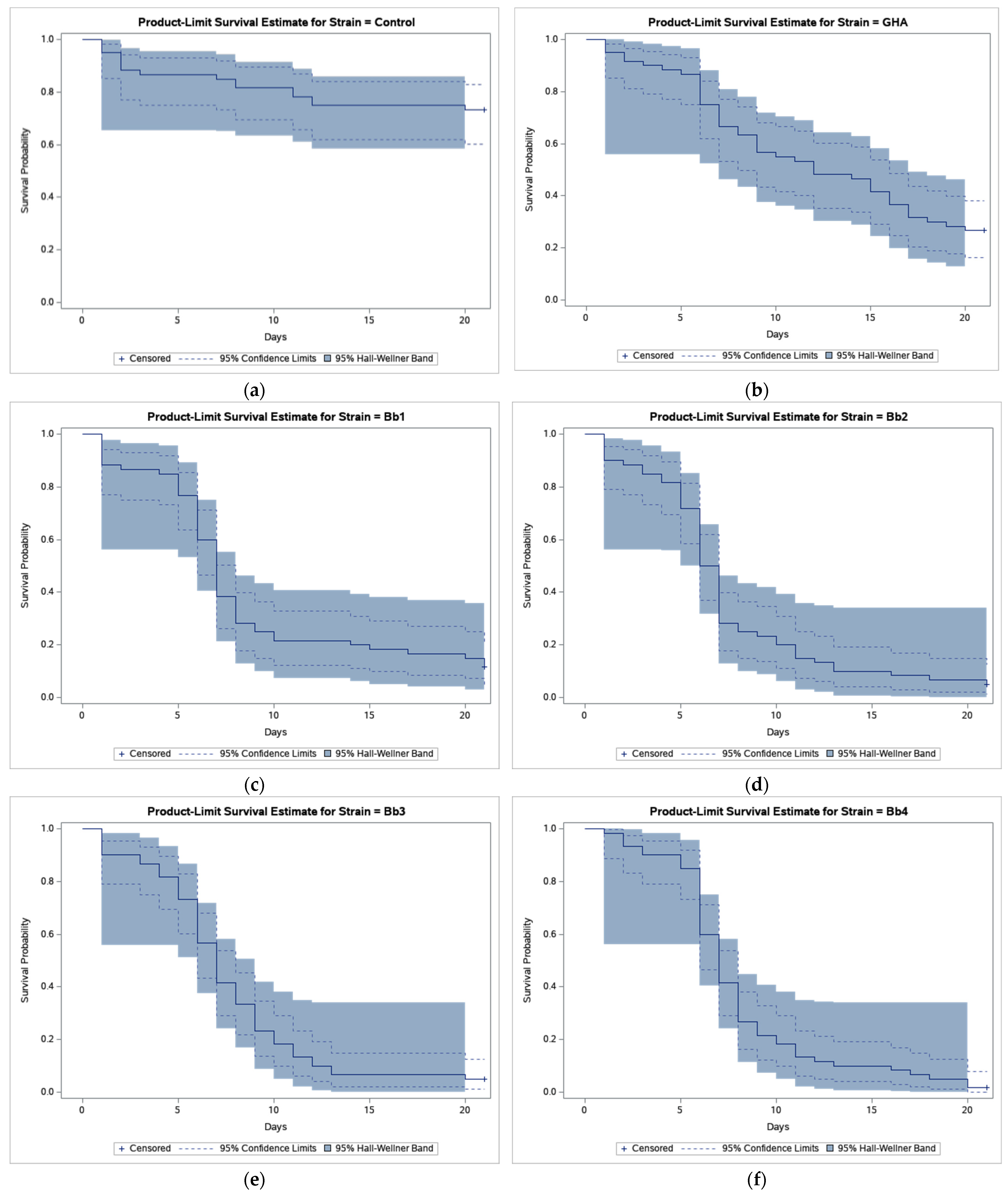
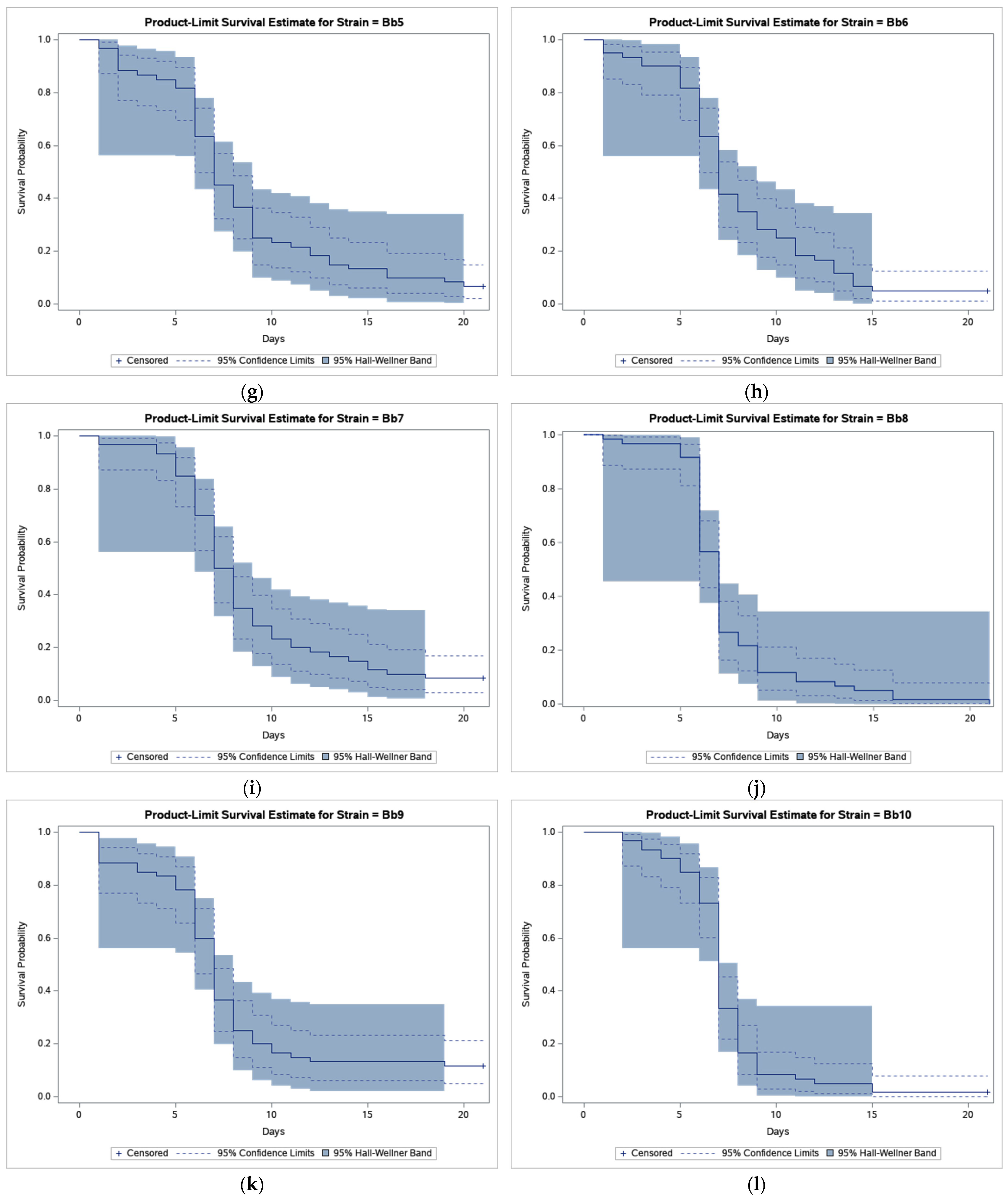
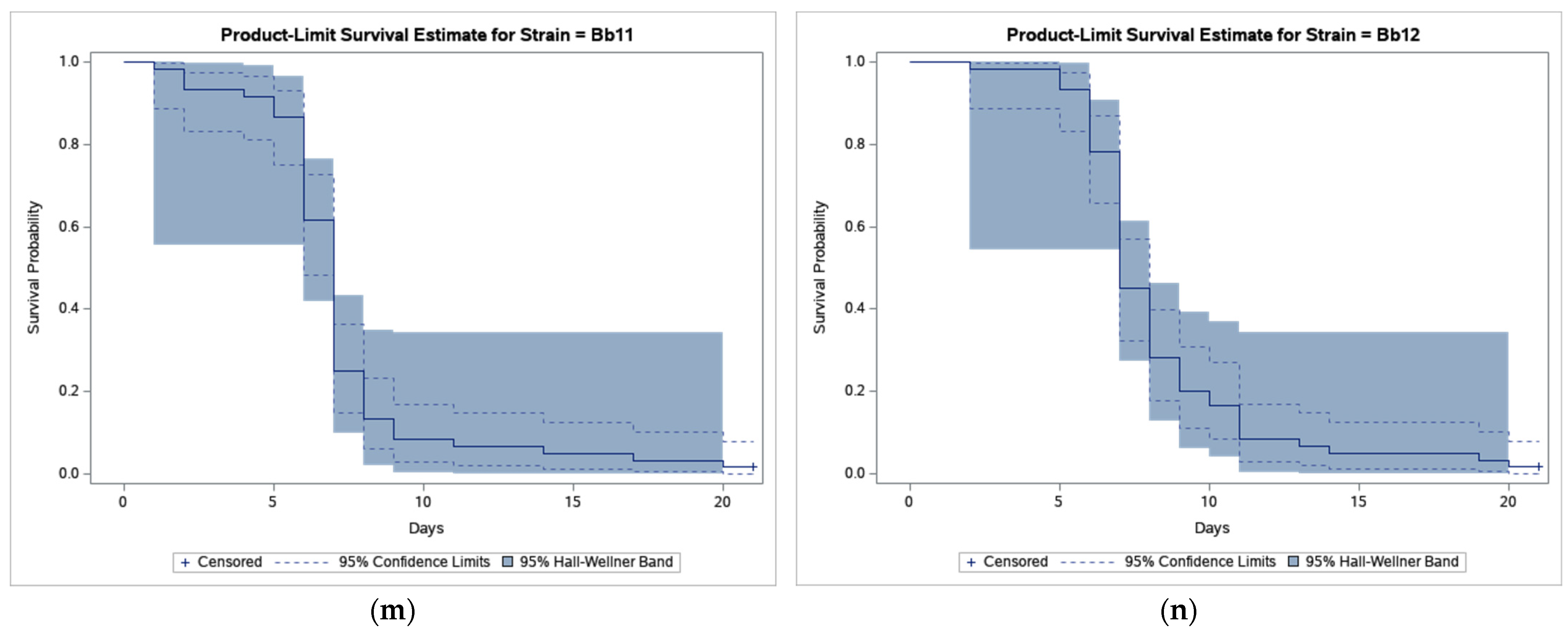
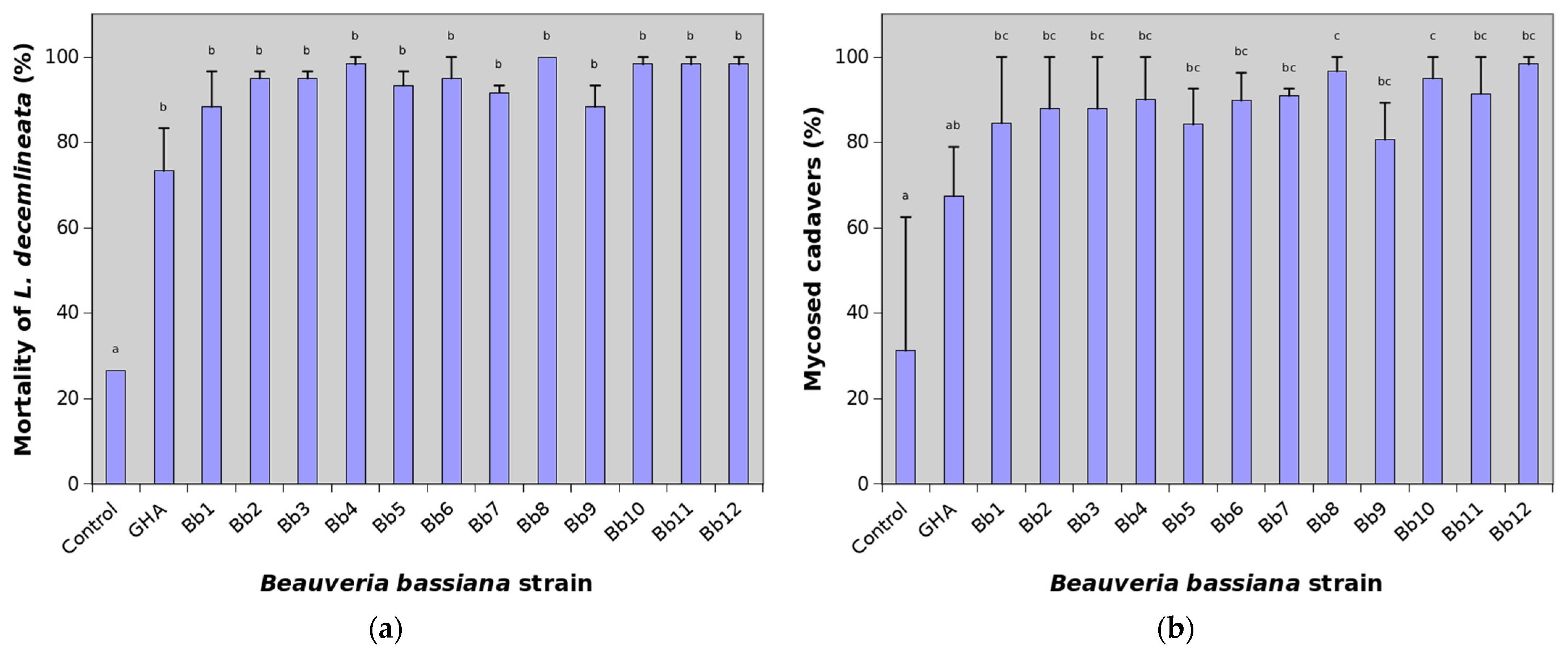
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jolivet, P. Le doryphore menace l’Asie Leptinotarsa decemlineata Say 1824 (Col. Chrysomelidae). Entomologiste 1991, 47, 29–48. [Google Scholar]
- Weber, D. Colorado Beetle: Pest on the Move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Hare, J.D. Ecology and Management of the Colorado Potato Beetle. Annu. Rev. Entomol. 1990, 35, 81–100. [Google Scholar] [CrossRef]
- Alyokhin, A. Colorado Potato Beetle Management on Potatoes: Current Challenges and Future Prospects. Fruit Veg. Cereal Sci. Biotechnol. 2009, 3, 10–19. [Google Scholar]
- Alyokhin, A.; Vincent, C.; Giordanengo, P. (Eds.) Insect Pests of Potato: Global Perspectives on Biology and Management; Elsevier: Amsterdam, The Netherlands; Academic Press: Boston, MA, USA, 2013; ISBN 978-0-12-386895-4. [Google Scholar]
- Mota-Sanchez, D.; Hollingworth, R.M.; Grafius, E.J.; Moyer, D.D. Resistance and Cross-Resistance to Neonicotinoid Insecticides and Spinosad in the Colorado Potato Beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2006, 62, 30–37. [Google Scholar] [CrossRef]
- Alyokhin, A.; Dively, G.; Patterson, M.; Castaldo, C.; Rogers, D.; Mahoney, M.; Wollam, J. Resistance and Cross-Resistance to Imidacloprid and Thiamethoxam in the Colorado Potato Beetle Leptinotarsa decemlineata. Pest Manag. Sci. 2007, 63, 32–41. [Google Scholar] [CrossRef]
- Zichová, T.; Kocourek, F.; Salava, J.; Nad’ova, K.; Stará, J. Detection of Organophosphate and Pyrethroid Resistance Alleles in Czech Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) Populations by Molecular Methods. Pest Manag. Sci. 2010, 66, 853–860. [Google Scholar] [CrossRef]
- Szendrei, Z.; Grafius, E.; Byrne, A.; Ziegler, A. Resistance to Neonicotinoid Insecticides in Field Populations of the Colorado Potato Beetle (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2012, 68, 941–946. [Google Scholar] [CrossRef]
- Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado Potato Beetle Resistance to Insecticides. Am. J. Potato Res. 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Stankovic, S.; Zabel, A.; Kostic, M.; Manojlovic, B.; Rajkovic, S. Colorado Potato Beetle [Leptinotarsa decemlineata (Say)] Resistance to Organophosphates and Carbamates in Serbia. J. Pest Sci. 2004, 77, 11–15. [Google Scholar] [CrossRef]
- Alyokhin, A.; Mota-Sanchez, D.; Baker, M.; Snyder, W.E.; Menasha, S.; Whalon, M.; Dively, G.; Moarsi, W.F. The Red Queen in a Potato Field: Integrated Pest Management versus Chemical Dependency in Colorado Potato Beetle Control. Pest Manag. Sci. 2015, 71, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Göldel, B.; Lemic, D.; Bažok, R. Alternatives to Synthetic Insecticides in the Control of the Colorado Potato Beetle (Leptinotarsa decemlineata Say) and Their Environmental Benefits. Agriculture 2020, 10, 611. [Google Scholar] [CrossRef]
- Kadoić Balaško, M.; Mikac, K.M.; Bažok, R.; Lemic, D. Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives. Insects 2020, 11, 581. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Srinivasan, R.; Cervantes, F.A. Occurrence of the Carabid Beetle, Pterostichus melanarius (Illiger), in Potato Ecosystems of Idaho and Its Predatory Potential on the Colorado Potato Beetle and Aphids. Am. J. Potato Res. 2013, 90, 83–92. [Google Scholar] [CrossRef]
- Sablon, L.; Haubruge, E.; Verheggen, F.J. Consumption of Immature Stages of Colorado Potato Beetle by Chrysoperla carnea (Neuroptera: Chrysopidae) Larvae in the Laboratory. Am. J. Potato Res. 2013, 90, 51–57. [Google Scholar] [CrossRef]
- Weber, D.C. Biological control of potato insect pests. In Insect Pests of Potato; Academic Press: Oxford, UK, 2013; pp. 399–437. ISBN 978-0-12-386895-4. [Google Scholar]
- Wright, R.J.; Agudelo-Silva, F.; Georgis, R. Soil Applications of Steinernematid and Heterorhabditid Nematodes for Control of Colorado Potato Beetles, Leptinotarsa decemlineata (Say). J. Nematol. 1987, 19, 201–206. [Google Scholar] [PubMed]
- Hussein, H.M.; Skoková Habuštová, O.; Půža, V.; Zemek, R. Laboratory Evaluation of Isaria fumosorosea CCM 8367 and Steinernema feltiae Ustinov against Immature Stages of the Colorado Potato Beetle. PLoS ONE 2016, 11, e0152399. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Arrizabalaga, M.; Villanueva, M.; Fernandez, A.B.; Caballero, P. A Strain of Bacillus thuringiensis Containing a Novel Cry7Aa2 Gene That Is Toxic to Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Insects 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-Fungal-Interactions: A Detailed Review on Entomopathogenic Fungi Pathogenicity to Combat Insect Pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A Comprehensive List with Worldwide Coverage and International Classification of Formulation Types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic Fungi as Biological Controllers: New Insights into Their Virulence and Pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect Pathogens as Biological Control Agents: Do They Have a Future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef] [Green Version]
- Bajan, C.; Kmitowa, K. The Effect of Entomogenous Fungi Paecilomyces farinosus Dicks. Brown et Smith and Beauveria bassiana Bals. Vuill. on the Oviposition by Leptinotarsa decemlineata Say Females, and on the Survival of Larvae. Ekol. Pol. 1972, 20, 423–432. [Google Scholar]
- Bajan, C.; Fedorko, A.; Kmitowa, K.; Wojciechowska, M. Utilization of Parasitic Microorganisms to Decrease Colorado Beetle Quantity. Bull. L’academie Pol. Sci. Ser. Sci. Biol. 1978, 26, 715–717. [Google Scholar]
- Samsinakova, A. Effects of Fungic Preparations on Larvae of Colorado Beetle, Leptinotarsa decemlineata. Acta Entomol. Bohemoslov. 1977, 74, 76–80. [Google Scholar]
- Ramisch, I. Paecilomyces farinosus Dicks. Ex Fr. Als Parasit Des Kartoffelkafers Leptinotarsa decemlineata Say. Nova Hedwig. 1976, 271, 199–214. [Google Scholar]
- Zemek, R.; Prenerova, E.; Weyda, F. The First Record of Entomopathogenic Fungus Paecilomyces fumosoroseus (Deuteromycota: Hyphomycetes) on the Hibernating Pupae of Cameraria ohridella (Lepidoptera: Gracillariidae). Entomol. Res. 2007, 37, A135–A136. [Google Scholar]
- Prenerová, E.; Zemek, R.; Volter, L.; Weyda, F. Strain of Entomopathogenic Fungus Isaria fumosorosea CCM 8367 (CCEFO.011.PFR) and the Method for Controlling Insect and Mite Pests. U.S. Patent No. US 08574566, 5 November 2013. [Google Scholar]
- Prenerová, E.; Zemek, R.; Volter, L.; Weyda, F. Strain of Entomopathogenic Fungus Isaria fumosorosea CCM 8367 (CCEFO.011.PFR) and the Method for Controlling Insect and Mite Pests. EPO Patent No. EP2313488, 29 April 2015. [Google Scholar]
- Ropek, D.; Kołodziejczyk, M. Efficacy of Selected Insecticides and Natural Preparations against Leptinotarsa decemlineata. Potato Res. 2019, 62, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.K.; Anderson, T.E.; Semel, M.; Roberts, D.W. Management of the Colorado Potato Beetle Using the Entomogenous Fungus Beauveria bassiana. Am. Potato J. 1985, 62, 29–37. [Google Scholar] [CrossRef]
- Fargues, J.; Delmas, J.C.; Lebrun, R.A. Leaf Consumption by Larvae of the Colorado Potato Beetle (Coleoptera: Chrysomelidae) Infected with the Entomopathogen, Beauveria bassiana. J. Econ. Entomol. 1994, 87, 67–71. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Application Parameters Affecting Field Efficacy of Beauveria bassiana Foliar Treatments against Colorado Potato Beetle Leptinotarsa decemlineata. Biol. Control 2002, 23, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Wraight, S.P.; Ramos, M.E. Delayed Efficacy of Beauveria bassiana Foliar Spray Applications against Colorado Potato Beetle: Impacts of Number and Timing of Applications on Larval and next-Generation Adult Populations. Biol. Control 2015, 83, 51–67. [Google Scholar] [CrossRef]
- Yulin, D.; Hui, W.; Zhiyan, M.; Liu, Y.; Xiaodong, D.; Deying, M. Identification and Virulence of a Fungal Entomopathogen Strain NDBJJ-BFG to Colorado Potato Beetle Leptinotarsa decemlineata (Say). J. Plant Prot. 2018, 45, 751–758. [Google Scholar]
- Baki, D.; Tosun, H.S.; Erler, F. Efficacy of Indigenous Isolates of Beauveria bassiana (Balsamo) Vuillemin (Deuteromycota: Hyphomycetes) against the Colorado Potato Beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Egypt. J. Biol. Pest Control 2021, 31, 56. [Google Scholar] [CrossRef]
- Chase, A.R.; Osborne, L.S.; Ferguson, V.M. Selective Isolation of the Entomopathogenic Fungi Beauveria bassiana and Metarhizium anisopliae from an Artificial Potting Medium. Fla. Entomol. 1986, 69, 285. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. ISBN 978-3-642-83964-1. [Google Scholar]
- O’Donnell, K. Ribosomal DNA Internal Transcribed Spacers Are Highly Divergent in the Phytopathogenic Ascomycete Fusarium sambucinum (Gibberella pulicaris). Curr. Genet. 1992, 22, 213–220. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Dillon, R.J.; Charnley, A.K. A Technique for Accelerating and Synchronising Germination of Conidia of the Entomopathogenic Fungus Metarhizium anisopliae. Arch. Microbiol. 1985, 142, 204–206. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Initiation of Germination in Conidia of the Entomopathogenic Fungus, Metarhizium anisopliae. Mycol. Res. 1990, 94, 299–304. [Google Scholar] [CrossRef]
- Skalický, A.; Bohatá, A.; Šimková, J.; Osborne, L.S.; Landa, Z. Selection of Indigenous Isolates of Entomopathogenic Soil Fungus Metarhizium anisopliae under Laboratory Conditions. Folia Microbiol. 2014, 59, 269–276. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT 14.3: User’s Guide; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- SAS Institute. SAS Stat Studio 3.8: User’s Guide; SAS Institute: Cary, NC, USA, 2018. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Kepenekci, I.; Oksal, E.; Saglam, H.; Atay, T.; Tulek, A.; Evlice, E. Identification of Turkish Isolate of the Entomopathogenic Fungi, Purpureocillium lilacinum (Syn: Paecilomyces lilacinus) and Its Effect on Potato Pests, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae) and Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Egypt. J. Biol. Pest Control 2015, 25, 121–127. [Google Scholar]
- Miętkiewski, R.; Sapieha, A.; Tkaczuk, C. The Effect of Soil-Borne Entomogenous Fungi on the Mycoses of the Colorado Potato Beetle during Hibernation Period. IOBC-WPRS Bull. 1996, 19, 162–165. [Google Scholar]
- Fargues, J.; Delmas, J.-C.; Augé, J.; Lebrun, R.A. Fecundity and Egg Fertility in the Adult Colorado Beetle (Leptinotarsa decemlineata) Surviving Larval Infection by the Fungus Beauveria bassiana. Entomol. Exp. Appl. 1991, 61, 45–51. [Google Scholar] [CrossRef]
- Long, D.W.; Drummond, F.A.; Groden, E. Susceptibility of Colorado Potato Beetle (Leptinotarsa decemlineata) Eggs to Beauveria bassiana. J. Invertebr. Pathol. 1998, 71, 182–183. [Google Scholar] [CrossRef] [PubMed]
- Fargues, J. Étude des conditions d’infection des larves de doryphore, Leptinotarsa decemlineata say, par Beauveria bassiana (Bals.) Vuill. [Fungi imperfecti]. Entomophaga 1972, 17, 319–337. [Google Scholar] [CrossRef]
- Hafez, M.; Zaki, F.N.; Moursy, A.; Sabbour, M. Biological Effects of the Entomopathogenic Fungus, Beauveria bassiana on the Potato Tuber Moth Phthorimaea operculella (Seller). Anz. Schädlingskunde Pflanzenschutz Umweltschutz 1997, 70, 158–159. [Google Scholar] [CrossRef]
- Tahir, M.; Wakil, W.; Ali, A.; Sahi, S.T. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae Isolates against Larvae of the Polyphagous Pest Helicoverpa armigera. Entomol. Gen. 2019, 38, 225–242. [Google Scholar] [CrossRef]
- Hajek, A.E.; Soper, R.S.; Roberts, D.W.; Anderson, T.E.; Biever, K.D.; Ferro, D.N.; LeBrun, R.A.; Storch, R.H. Foliar Applications of Beauveria bassiana (Balsamo) Vuillemin for Control of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae): An Overview of Pilot Test Results from the Northern United States. Can. Entomol. 1987, 119, 959–974. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Effects of Inoculation Method on Efficacy of Wettable Powder and Oil Dispersion Formulations of Beauveria bassiana against Colorado Potato Beetle Larvae under Low-Humidity Conditions. Biocontrol Sci. Technol. 2017, 27, 348–363. [Google Scholar] [CrossRef]
- Zimmermann, G. Effect of High Temperatures and Artificial Sunlight on the Viability of Conidia of Metarhizium anisopliae. J. Invertebr. Pathol. 1982, 40, 36–40. [Google Scholar] [CrossRef]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Vidal, C.; Lacey, L.A.; Lomer, C.J.; Rougier, M. Variability in Susceptibility to Simulated Sunlight of Conidia among Isolates of Entomopathogenic Hyphomycetes. Mycopathologia 1996, 135, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Inyang, E.N.; McCartney, H.A.; Oyejola, B.; Ibrahim, L.; Pye, B.J.; Archer, S.A.; Butt, T.M. Effect of Formulation, Application and Rain on the Persistence of the Entomogenous Fungus Metarhizium anisopliae on Oilseed Rape. Mycol. Res. 2000, 104, 653–661. [Google Scholar] [CrossRef]
- Klingen, I.; Hajek, A.; Meadow, R.; Renwick, J.A.A. Effect of Brassicaceous Plants on the Survival and Infectivity of Insect Pathogenic Fungi. BioControl 2002, 47, 411–425. [Google Scholar] [CrossRef]
- Pilz, C.; Enkerli, J.; Wegensteiner, R.; Keller, S. Establishment and Persistence of the Entomopathogenic Fungus Metarhizium anisopliae in Maize Fields. J. Appl. Entomol. 2011, 135, 393–403. [Google Scholar] [CrossRef]
- Zemek, R.; Konopická, J.; Ul Abdin, Z. Low Efficacy of Isaria fumosorosea against Box Tree Moth Cydalima perspectalis: Are Host Plant Phytochemicals Involved in Herbivore Defence against Fungal Pathogens? J. Fungi 2020, 6, 342. [Google Scholar] [CrossRef] [PubMed]
- Inglis, G.D.; Goettel, M.S.; Butt, T.M.; Strasser, H. Use of hyphomycetous fungi for managing insect pests. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CABI: Wallingford, UK, 2001; pp. 23–69. ISBN 978-0-85199-356-0. [Google Scholar]
- Vandenberg, J.D.; Ramos, M.; Altre, J.A. Dose–Response and Age- and Temperature-Related Susceptibility of the Diamondback Moth (Lepidoptera: Plutellidae) to Two Isolates of Beauveria bassiana (Hyphomycetes: Moniliaceae). Environ. Entomol. 1998, 27, 1017–1021. [Google Scholar] [CrossRef] [Green Version]
- Glare, T.R.; Milner, R.J. Ecology of entomopathogenic fungi. In Handbook of Applied Mycology; Arora, D.K., Ajello, L., Mukerji, K.G., Eds.; Marcel Dekker: New York, NY, USA, 1991; Volume 2, pp. 547–612. ISBN 978-0-8247-8435-5. [Google Scholar]
- Ferron, P.; Fargues, J.; Riba, D. Fungi as microbial insecticides against pests. In Handbook of Applied Mycology; Arora, D.K., Ajello, L., Mukerji, K.G., Eds.; Marcel Dekker: New York, NY, USA, 1991; Volume 2, pp. 665–705. ISBN 978-0-8247-8435-5. [Google Scholar]
- Tefera, T.; Vidal, S. Effect of Inoculation Method and Plant Growth Medium on Endophytic Colonization of Sorghum by the Entomopathogenic Fungus Beauveria bassiana. BioControl 2009, 54, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Raya-Diaz, S.; Sanchez-Rodriguez, A.R.; Segura-Fernandez, J.M.; del Campillo, M.C.; Quesada-Moraga, E. Entomopathogenic Fungi-Based Mechanisms for Improved Fe Nutrition in Sorghum Plants Grown on Calcareous Substrates. PLoS ONE 2017, 12, e0185903. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Bauer, L.S. Microbial Control of Emerald Ash Borer, Agrilus planipennis (Coleoptera: Buprestidae) with Beauveria bassiana Strain GHA: Greenhouse and Field Trials. Biol. Control 2008, 45, 124–132. [Google Scholar] [CrossRef]
- Mukawa, S.; Tooyama, H.; Ikegami, T. Influence of Humidity on the Infection of Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), by Beauveria bassiana. Appl. Entomol. Zool. 2011, 46, 255–264. [Google Scholar] [CrossRef]
- Clavet, C.; Hampton, E.; Requintina, M.; Alm, S.R. Laboratory Assessment of Beauveria bassiana (Hypocreales: Clavicipitaceae) Strain GHA for Control of Listronotus maculicollis (Coleoptera: Curculionidae) Adults. J. Econ. Entomol. 2013, 106, 2322–2326. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.L.; Skinner, M.; Gouli, S.; Gouli, V.; Kim, J.S. Virulence of BotaniGard® to Second Instar Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Insects 2015, 6, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.S.; Lim, U.T. Laboratory Bioassay of Beauveria bassiana against Tetranychus urticae (Acari: Tetranychidae) on Leaf Discs and Potted Bean Plants. Exp. Appl. Acarol. 2015, 65, 307–318. [Google Scholar] [CrossRef]
- Li, H.; Huang, D.; Wang, Z.; Yan, H.; Zheng, J. Screening Test of Highly Virulent Strains of Entomopathogenic Fungi Beauveria bassiana against Apriona germari Larvae. Sci. Silvae Sin. 2007, 43, 66–71. [Google Scholar]
- Wang, Y.-C.; Chi, D.-F. Screening of High-Virulent Entomopathogenic Fungal Strains to Infect Xylotrechus rusticus Larvae. 3Biotech 2019, 9, 80. [Google Scholar] [CrossRef]
- Guillebeau, L.P. Risk–Benefit Analysis of Pesticides: The U.S. Environmental Protection Agency Perspective. Am. Entomol. 1994, 40, 173–179. [Google Scholar] [CrossRef]
- Wakil, W.; Kavallieratos, N.G.; Ghazanfar, M.U.; Usman, M.; Habib, A.; El-Shafie, H.A.F. Efficacy of Different Entomopathogenic Fungal Isolates against Four Key Stored-Grain Beetle Species. J. Stored Prod. Res. 2021, 93, 101845. [Google Scholar] [CrossRef]
- Viaud, M.; Couteaudier, Y.; Levis, C.; Riba, G. Genome Organization in Beauveria bassiana: Electrophoretic Karyotype, Gene Mapping, and Telomeric Fingerprint. Fungal Genet. Biol. 1996, 20, 175–183. [Google Scholar] [CrossRef]
- Maurer, P.; Couteaudier, Y.; Girard, P.A.; Bridge, P.D.; Riba, G. Genetic Diversity of Beauveria bassiana and Relatedness to Host Insect Range. Mycol. Res. 1997, 101, 159–164. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, Y.; Xu, W.; Sui, L.; Du, Q.; Wang, Y.; Zhao, Y.; Li, Q. Influence of Genetic Diversity of Seventeen Beauveria bassiana Isolates from Different Hosts on Virulence by Comparative Genomics. BMC Genom. 2020, 21, 451. [Google Scholar] [CrossRef]
- Feng, M.G.; Poprawski, T.J.; Khachatourians, G.G. Production, Formulation and Application of the Entomopathogenic Fungus Beauveria bassiana for Insect Control: Current Status. Biocontrol Sci. Technol. 1994, 4, 3–34. [Google Scholar] [CrossRef]
- Jaronski, S.T. Mass Production of Entomopathogenic Fungi: State of the Art. In Mass Production of Beneficial Organisms; Elsevier: Amsterdam, The Netherlands, 2014; pp. 357–413. ISBN 978-0-12-391453-8. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sampling Site | GPS Coordinates | Genbank Accession Number |
|---|---|---|---|
| Bb1 | České Budějovice | 48.97417° N, 14.44867° E | MN560148.1 |
| Bb2 | České Budějovice | 48.97601° N, 14.44720° E | MN749309 |
| Bb3 | České Budějovice | 48.97417° N, 14.44867° E | MN749310 |
| Bb4 | Malonty 1 | 48.69105° N, 14.58950° E | MN749311 |
| Bb5 | Malonty 1 | 48.69105° N, 14.58950° E | MN749312 |
| Bb6 | Malonty 1 | 48.69105° N, 14.58950° E | MN749313 |
| Bb7 | Bělčice | 49.50702° N, 13.89545° E | MN749314 |
| Bb8 | Bělčice | 49.50702° N, 13.89545° E | MN749315 |
| Bb9 | Oblajovice | 49.44965° N, 14.88024° E | MN749316 |
| Bb10 | Bojanovice | 49.29724° N, 13.62259° E | MN749317 |
| Bb11 | Bojanovice | 49.29724° N, 13.62259° E | MN749318 |
| Bb12 | Bojanovice | 49.29724° N, 13.62259° E | MN749319 |
| Strain | Mortality 1 (%) | Survival Time 2 (Days) | LT50 (95% CI) (Days) | Log-Rank Test 3 |
|---|---|---|---|---|
| Bb1 | 84.09 | 8.83 ± 0.81 | 7.0 (6.0–8.0) | A |
| Bb2 | 93.18 | 7.58 ± 0.64 | 6.5 (6.0–7.0) | A |
| Bb3 | 93.18 | 7.65 ± 0.58 | 7.0 (6.0–8.0) | A |
| Bb4 | 97.73 | 8.05 ± 0.55 | 7.0 (6.0–8.0) | A |
| Bb5 | 90.91 | 8.52 ± 0.65 | 7.0 (6.0–8.0) | A |
| Bb6 | 93.18 | 7.97 ± 0.47 | 7.0 (6.0–8.0) | A |
| Bb7 | 88.64 | 8.77 ± 0.55 | 7.5 (7.0–8.0) | A |
| Bb8 | 100.00 | 7.50 ± 0.41 | 7.0 (6.0–7.0) | A |
| Bb9 | 84.09 | 7.90 ± 0.65 | 7.0 (6.0–7.0) | A |
| Bb10 | 97.73 | 7.27 ± 0.33 | 7.0 (NA-NA) | A |
| Bb11 | 97.73 | 7.25 ± 0.43 | 7.0 (6.0–7.0) | A |
| Bb12 | 97.73 | 8.28 ± 0.43 | 7.0 (7.0–8.0) | A |
| GHA | 63.64 | 12.33 ± 0.85 | 12.0 (8.0–16.0) | B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zemek, R.; Konopická, J.; Jozová, E.; Skoková Habuštová, O. Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata. Insects 2021, 12, 1077. https://doi.org/10.3390/insects12121077
Zemek R, Konopická J, Jozová E, Skoková Habuštová O. Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata. Insects. 2021; 12(12):1077. https://doi.org/10.3390/insects12121077
Chicago/Turabian StyleZemek, Rostislav, Jana Konopická, Eva Jozová, and Oxana Skoková Habuštová. 2021. "Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata" Insects 12, no. 12: 1077. https://doi.org/10.3390/insects12121077
APA StyleZemek, R., Konopická, J., Jozová, E., & Skoková Habuštová, O. (2021). Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata. Insects, 12(12), 1077. https://doi.org/10.3390/insects12121077