
Nitrogenous Fertilizer Reduces Resistance but Enhances Tolerance to the Brown Planthopper in Fast-Growing, Moderately Resistant Rice

 ,
,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Planthoppers
2.3. Effects of Nitrogen on Rice–Planthopper Interactions
2.4. Data Analyses
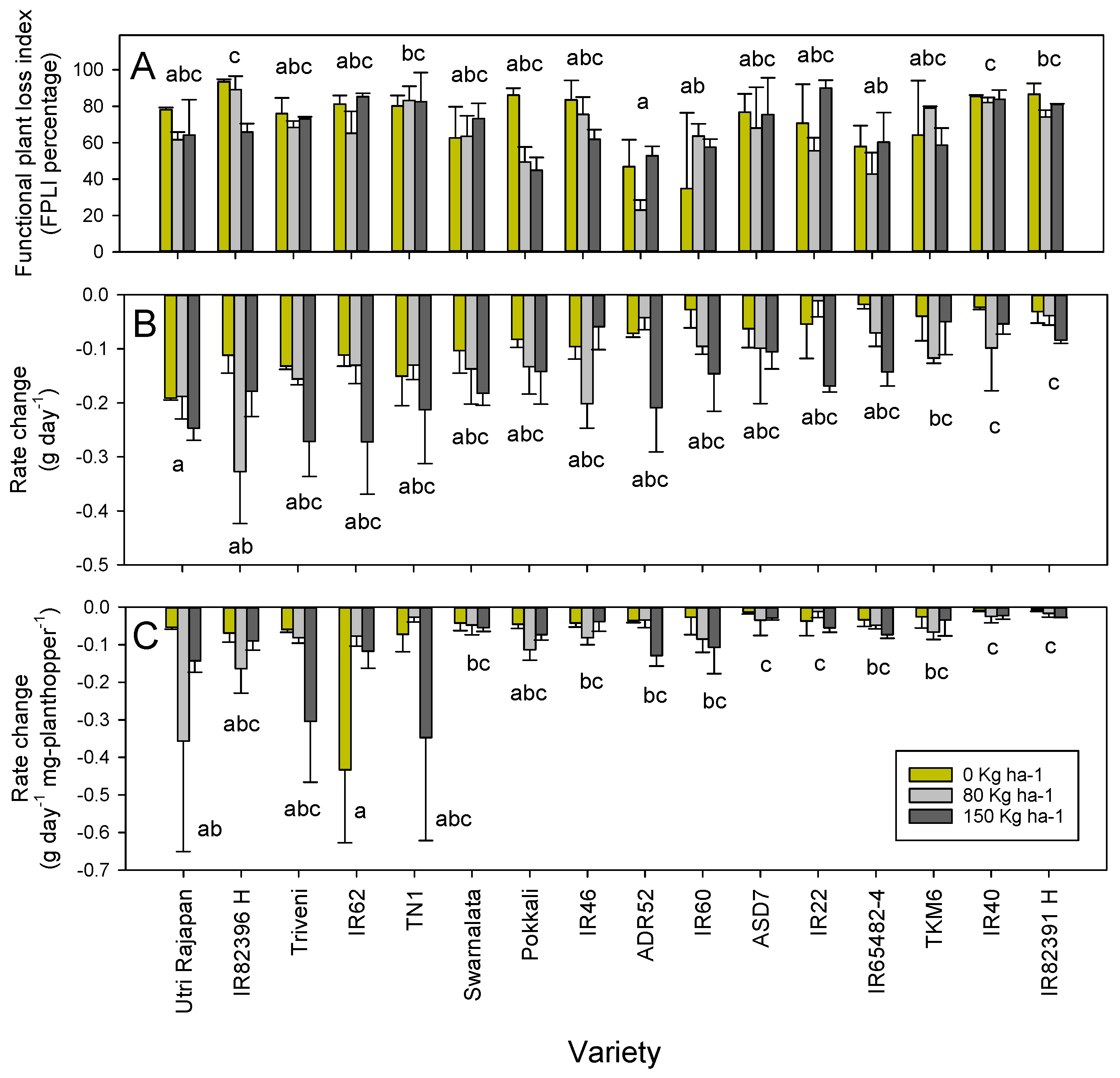
3. Results
3.1. Host Resistance
3.2. Plant Biomass and Growth Rates
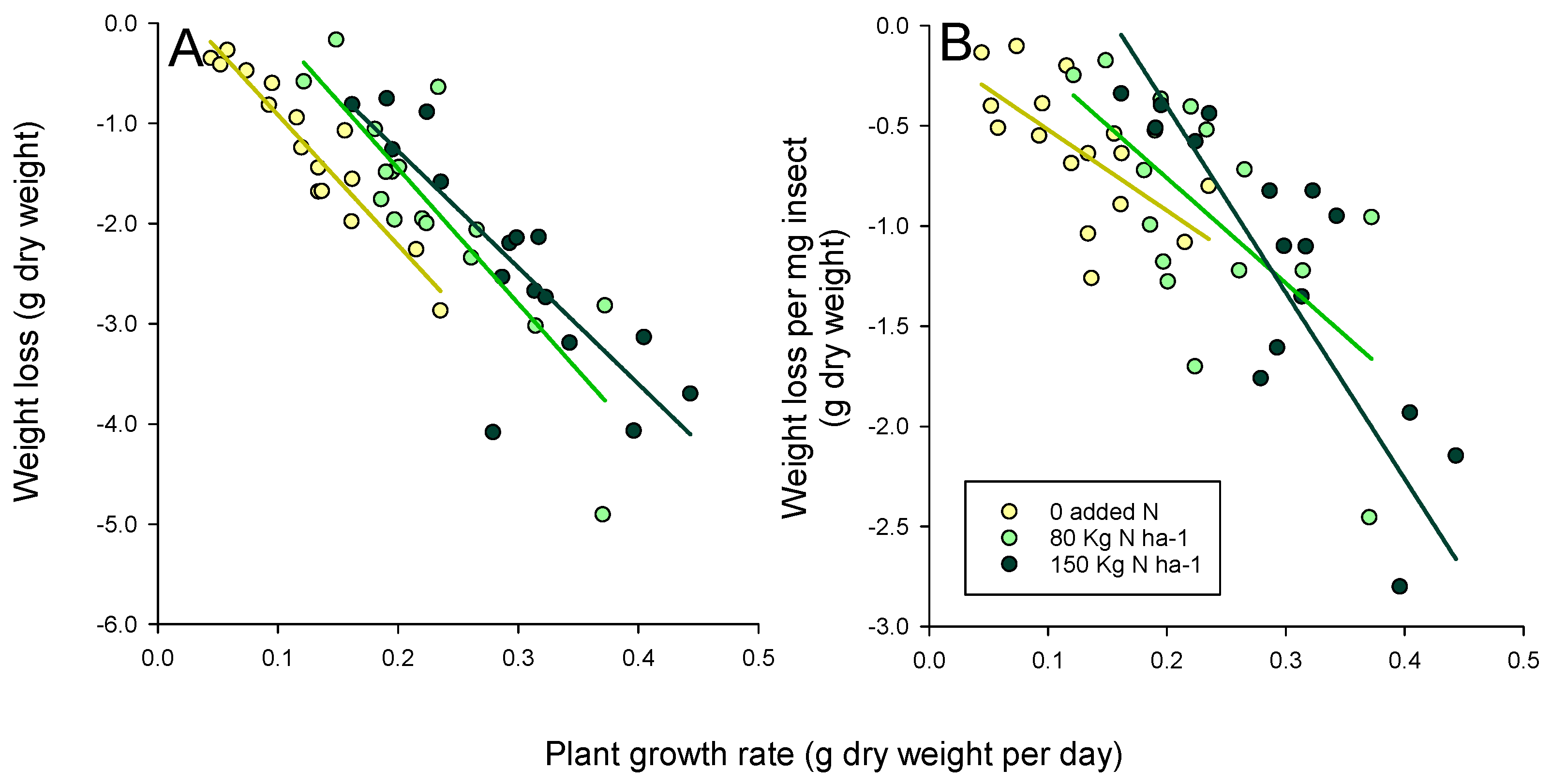
3.3. Planthopper Effects on Plants
4. Discussion
4.1. Effects of Adding Nitrogen on Planthopper Fitness
4.2. Effects of Adding Nitrogen on Rice Tolerance
4.3. The Need to Combine Resistance with Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Stout, M.J. Reevaluating the conceptual framework for applied research on host-plant resistance. Insect Sci. 2013, 20, 263–272. [Google Scholar] [CrossRef]
- Horgan, F.G. Diversity and defence: Plant-herbivore interactions at multiple scales and trophic levels. In Biodiversity and Insect Pests: Key Issues for Sustainable Management, 1st ed.; Gurr, G.M., Wratten, S.D., Snyder, W.E., Read, D.M.Y., Eds.; John Wiley & Sons: Oxford, UK, 2012; pp. 421–458. [Google Scholar]
- Bari, A.; Street, K.; Mackay, M.; Endresen, D.T.F.; De Pauw, E.; Ahmed, A. Focused identification of germplasm strategy (FIGS) detects wheat stem rust resistance linked to environmental variables. Genet. Resour. Crop. Evol. 2012, 59, 1465–1481. [Google Scholar] [CrossRef]
- Flanders, K.L.; Radcliffe, E.B.; Hawkes, J.G. Geographic distribution of insect resistance in potatoes. Euphytica 1997, 93, 201–221. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-López, S.; Villamil, N.; Zedillo-Avelleyra, P.; Boege, K. Plant defence as a complex and changing phenotype throughout ontogeny. Ann. Bot. 2015, 116, 797–806. [Google Scholar] [CrossRef]
- Horgan, F.G. Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers. Crop. Prot. 2018, 110, 21–33. [Google Scholar] [CrossRef]
- William, J.; Mattson, J. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Zebitz, C.P.W.; Kehlenbeck, H. Performance of Aphis fabae on chocolate spot diseased faba bean plants. Phytoparasitica 1991, 19, 113–119. [Google Scholar] [CrossRef]
- Prüter, C.; Zebitz, C.P.W. Effects of Aphis fabae and Uromyces viciae-fabae on the growth of a susceptible and an aphid resistant cultivar of Vicia faba. Ann. Appl. Biol. 1991, 119, 215–226. [Google Scholar] [CrossRef]
- Cronin, J.T.; Abrahamson, W.G. Host-plant genotype and other herbivores influence goldenrod stem galler preference and performance. Oecologia 1999, 121, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Astles, P.A.; Moore, A.J.; Preziosi, R.F. Genetic variation in response to an indirect ecological effect. Proc. R. Soc. B Biol. Sci. 2005, 272, 2577–2581. [Google Scholar] [CrossRef]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Postive and negative interspecific interactions between coexisting rice planthoppers neutralize the effects of elevated temperatures. Funct. Ecol. 2021, 35, 181–192. [Google Scholar] [CrossRef]
- Butler, J.; Garratt, M.P.D.; Leather, S.R. Fertilisers and insect herbivores: A meta-analysis. Ann. Appl. Biol. 2012, 161, 223–233. [Google Scholar] [CrossRef]
- Jiang, M.X.; Cheng, J.A. Feeding, oviposition and survival of overwintered rice water weevil (Coleoptera: Curculionidae) adults in response to nitrogen fertilization of rice at seedling stage. Appl. Entomol. Zool. 2003, 38, 543–549. [Google Scholar] [CrossRef][Green Version]
- Jiang, M.X.; Cheng, J.A. Interactions between the striped stem borer Chilo suppressalis (Walk.) (Lep., Pyralidae) larvae and rice plants in response to nitrogen fertilization. J. Pest Sci. 2003, 76, 124–128. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol-Martínez, E.; Almazan, M.L.P.; Romena, A.; Ramal, A.F.; Ferrater, J.B.; Bernal, C.C. Susceptibility and tolerance in hybrid and pure-line rice varieties to herbivore attack: Biomass partitioning and resource-based compensation in response to damage. Ann. Appl. Biol. 2016, 169, 200–213. [Google Scholar] [CrossRef]
- Horgan, F.G.; Cruz, A.P.; Bernal, C.C.; Ramal, A.F.; Almazan, M.L.P.; Wilby, A. Resistance and tolerance to the brown planthopper, Nilaparvata lugens (Stål), in rice infested at different growth stages across a gradient of nitrogen applications. Field Crop. Res. 2018, 217, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Rubia-Sanchez, E.; Suzuki, Y.; Miyamoto, K.; Watanabe, T. The potential for compensation of the effects of the brown planthopper Nilaparvata lugens Stal (Homoptera: Delphacidae) feeding on rice. Crop. Prot. 1999, 18, 39–45. [Google Scholar] [CrossRef]
- Rubia, E.G.; Heong, K.L.; Zalucki, M.; Gonzales, B.; Norton, G.A. Mechanisms of compensation of rice plants to yellow stem borer Scirpophaga incertulas (Walker) injury. Crop. Prot. 1996, 15, 335–340. [Google Scholar] [CrossRef]
- Mgoo, V.H.; Makundi, R.H.; Pallangyo, B.; Schulthess, F.; Jiang, N.; Omwega, C.O. Yield loss due to the stemborer Chilo partellus (Swinhoe) (Lepidoptera: Crambidae) at different nitrogen application rates to maize. Ann. Soc. Entomol. Fr. (NS) 2006, 42, 487–494. [Google Scholar] [CrossRef]
- Horgan, F.G. Integrated pest management for sustainable rice cultivation: A holistic approach. In Achieving Sustainable Cultivation of Rice: Cultivation, Pest and Disease Management; Sasaki, T., Ed.; Burleigh Dodds Scientific: Cambridge, UK, 2017; pp. 309–342. [Google Scholar]
- Koch, K.G.; Chapman, K.; Louis, J.; Heng-Moss, T.; Sarath, G. Plant tolerance: A unique approach to control hemipteran pests. Front. Plant Sci. 2016, 7, 1363. [Google Scholar] [CrossRef]
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant defense against herbivorous pests: Exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 7, 1132. [Google Scholar] [CrossRef]
- Panda, N.; Heinrichs, E.A. Levels of tolerance and antibiosis in rice varieties having moderate resistance to the brown planthopper, Nilaparvata lugens (Stål) (Hemiptera: Delphacidae). Environ. Entomol. 1983, 12, 1204–1214. [Google Scholar] [CrossRef]
- Roy, D.; Chakraborty, G.; Biswas, A.; Sarkar, P.K. Antixenosis, tolerance and genetic analysis of some rice landraces for resistance to Nilaparvata lugens (Stål.). J. Asia-Pac. Entomol. 2021, 24, 448–460. [Google Scholar] [CrossRef]
- Sarao, P.S.; Bentur, J.S. Antixenosis and tolerance of rice genotypes against brown planthopper. Rice Sci. 2016, 23, 96–103. [Google Scholar] [CrossRef]
- Villegas, J.M.; Wilson, B.E.; Stout, M.J. Assessment of tolerance and resistance of inbred rice cultivars to combined infestations of rice water weevil and stemborers. Entomol. Exp. Appl. 2021, 169, 629–639. [Google Scholar] [CrossRef]
- Villegas, J.M.; Wilson, B.E.; Way, M.O.; Gore, J.; Stout, M.J. Tolerance to rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), infestations among hybrid and inbred rice cultivars in the Southern U.S. Crop. Prot. 2021, 139, 105368. [Google Scholar] [CrossRef]
- Islam, Z.; Karim, A.N.M.R. Whiteheads associated with stem borer infestation in modern rice varieties: An attempt to resolve the dilemma of yield losses. Crop. Prot. 1997, 16, 303–311. [Google Scholar] [CrossRef]
- Stout, M.J.; Rice, W.C.; Ring, D.R. The influence of plant age on tolerance of rice to injury by the rice water weevil, Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Bull. Entomol. Res. 2007, 92, 177–184. [Google Scholar] [CrossRef]
- Rashid, M.M.; Jahan, M.; Islam, K.S.; Latif, M.A. Ecological fitness of brown planthopper, Nilaparvata lugens (Stål), to rice nutrient management. Ecol. Process. 2017, 6, 15. [Google Scholar] [CrossRef]
- Visarto, P.; Zalucki, M.P.; Nesbitt, H.J.; Jahn, G.C. Effect of fertilizer, pesticide treatment, and plant variety on the realized fecundity and survival rates of brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae) generating outbreaks in Cambodia. J. Asia-Pac. Entomol. 2001, 4, 75–84. [Google Scholar] [CrossRef]
- Rashid, M.M.; Ahmed, N.; Jahan, M.; Islam, K.S.; Nansen, C.; Willers, J.L.; Ali, M.P. Higher fertilizer inputs increase fitness traits of brown planthopper in rice. Sci. Rep. 2017, 7, 4719. [Google Scholar] [CrossRef]
- Horgan, F.G.; Ramal, A.F.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Iswanto, E.H.; Van Chien, H.; Phyu, M.H.; Bernal, C.C.; et al. Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia against resistant rice varieties. Crop. Prot. 2015, 78, 222–231. [Google Scholar] [CrossRef]
- Khush, G.S.; Virk, P.S. IR Varieties and Their Impact; International Rice Research Institute: Los Baños, Philippines, 2005. [Google Scholar]
- Martinez, C.R.; Khush, G.S. Sources and inheritance of resistance to brown planthopper in some breeding lines of rice. Crop Sci. 1974, 14, 264–267. [Google Scholar] [CrossRef]
- Horgan, F.G.; Srinivasan, T.S.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Van Chien, H.; Almazan, M.L.P.; Bernal, C.C.; Ramal, A.F.; et al. Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers: Implications for rice breeding and gene deployment. Agronomy 2017, 7, 62. [Google Scholar] [CrossRef]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds. Theor. Appl. Genet. 2010, 121, 1601–1611. [Google Scholar] [CrossRef]
- Murata, K.; Fujiwara, M.; Murai, H.; Takumi, S.; Mori, N.; Nakamura, C. Mapping of a brown planthopper (Nilaparvata lugens Stål) resistance gene Bph9 on the long arm of rice chromosome 12. Cereal Res. Commun. 2001, 29, 245–250. [Google Scholar] [CrossRef]
- Srinivasan, T.S.; Almazan, M.L.P.; Bernal, C.C.; Fujita, D.; Ramal, A.F.; Yasui, H.; Subbarayalu, M.K.; Horgan, F.G. Current utility of the BPH25 and BPH26 genes and possibilities for further resistance against plant- and leafhoppers from the donor cultivar ADR52. Appl. Entomol. Zool. 2015, 50, 533–543. [Google Scholar] [CrossRef]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 2012, 124, 495–504. [Google Scholar] [CrossRef]
- Jena, K.K.; Jeung, J.U.; Lee, J.H.; Choi, H.C.; Brar, D.S. High-resolution mapping of a new brown planthopper (BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 112, 288–297. [Google Scholar] [CrossRef]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J.; Su, X.; Luo, H.; Ren, G. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 37645. [Google Scholar] [CrossRef]
- Horgan, F.G.; Srinivasan, T.S.; Naik, B.S.; Ramal, A.F.; Bernal, C.C.; Almazan, M.L.P. Effects of nitrogen on egg-laying inhibition and ovicidal response in planthopper-resistant rice varieties. Crop. Prot. 2016, 89, 223–230. [Google Scholar] [CrossRef]
- Magalit, V.; Saxena, R. Tolerance of Utri Rajapan to biotype 2 of Brown Planthopper, Nilaparvata Lugens (Stål). In Proceedings of the 1988 Philippines Plant Protection Conference, Cebu City, Philippines, 3–7 May 1988; University of the Philippines: Los Baños, Philippines, 1991; p. 15. [Google Scholar]
- Ho, D.T.; Heinrichs, E.A.; Medrano, F. Tolerance of the rice variety Triveni to the brown planthopper, Nilaparvata lugens. Environ. Entomol. 1982, 11, 598–602. [Google Scholar] [CrossRef]
- Horgan, F.G.; Mundaca, E.A.; Quintana, R.; Naredo, A.I.; Almazan, M.L.P.; Bernal, C.C. Efficacy and cost-effectiveness of phenotyping for rice resistance and tolerance to planthoppers. Insects 2021, 12, 847. [Google Scholar] [CrossRef]
- Horgan, F.G.; Jauregui, A.; Peñalver Cruz, A.; Crisol Martínez, E.; Bernal, C.C. Changes in reflectance of rice seedlings during planthopper feeding as detected by digital camera: Potential applications for high-throughput phenotyping. PLoS ONE 2020, 15, e0238173. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, B., Everitt, W., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley Online Library: Oxford, UK, 2017; pp. 1–15. [Google Scholar] [CrossRef]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol Martínez, E.; Stuart, A.M.; Bernal, C.C.; de Cima Martín, E.; Almazan, M.L.P.; Ramal, A.F. Effects of vegetation strips, fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystem. Insects 2019, 10, 328. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-X.; Heong, K.-L.; Yu, X.-P.; Hu, C. Effects of plant nitrogen on ecological fitness of the brown planthopper, Nilaparvata lugens Stal. in rice. J. Asia-Pac. Entomol. 2004, 7, 97–104. [Google Scholar] [CrossRef]
- Rashid, M.M.; Jahan, M.; Islam, K.S. Response of adult brown planthopper Nilaparvata lugens (Stål) to rice nutrient management. Neotrop. Entomol. 2016, 45, 588–596. [Google Scholar] [CrossRef]
- Rashid, M.M.; Jahan, M.; Islam, K.S. Effects of nitrogen, phosphorous and potassium on host-choice behavior of brown planthopper, Nilaparvata lugens (Stål) on rice cultivar. J. Insect Behav. 2017, 30, 1–15. [Google Scholar] [CrossRef]
- Swati, S.; Sharma, A.; Dubey, V. Probing behaviour of brown planthopper, Nilaparvata lugens (Stal.) on rice plant as influenced by nitrogen application. J. Appl. Zool. Res. 2013, 24, 29–32. [Google Scholar]
- Zhong-xian, L.; Villareal, S.; Xiao-ping, Y.; Heong, K.; Cui, H. Effect of nitrogen on water content, sap flow, and tolerance of rice plants to brown planthopper, Nilaparvata lugens. Rice Sci. 2004, 11, 129–134. [Google Scholar]
- Cagampang, G.; Pathak, M.; Juliano, B. Metabolic changes in the rice plant during infestation by the brown planthopper, Nilaparvata lugens Stål (Hemiptera: Delphacidae). Appl. Entomol. Zool. 1974, 9, 174–184. [Google Scholar] [CrossRef]
- Liu, J.; Wu, J. Effects of Nilaparvata lugens (Stål)(Homoptera: Delphacidae) infestation on the uptake level of nitrogen, phosphorus and potassium by roots of rice varieties with different resistant levels. Acta Entomol. Sin. 2010, 53, 411–419. [Google Scholar]
- Liu, J.-L.; Yu, J.-F.; Wu, J.-C.; Yin, J.-L.; Gu, H.-N. Physiological responses to Nilaparvata lugens in susceptible and resistant rice varieties: Allocation of assimilates between shoots and roots. J. Econ. Entomol. 2008, 101, 384–390. [Google Scholar] [CrossRef]
- Ferrater, J.B.; Naredo, A.I.; Almazan, M.L.P.; de Jong, P.W.; Dicke, M.; Horgan, F.G. Varied responses by yeast-like symbionts during virulence adaptation in a monophagous phloem-feeding insect. Arthropod-Plant Interact. 2015, 9, 215–224. [Google Scholar] [CrossRef]
- Pompon, J.; Quiring, D.; Giordanengo, P.; Pelletier, Y. Role of xylem consumption on osmoregulation in Macrosiphum euphorbiae (Thomas). J. Insect Physiol. 2010, 56, 610–615. [Google Scholar] [CrossRef]
- Pompon, J.; Quiring, D.; Goyer, C.; Giordanengo, P.; Pelletier, Y. A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential. J. Insect Physiol. 2011, 57, 1317–1322. [Google Scholar] [CrossRef]
- Wu, X.; Yu, Y.; Baerson, S.R.; Song, Y.; Liang, G.; Ding, C.; Niu, J.; Pan, Z.; Zeng, R. Interactions between nitrogen and silicon in rice and their effects on resistance toward the brown planthopper Nilaparvata lugens. Front. Plant Sci. 2017, 8, 28. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Li, P.; Wen, L.; Hou, M. Silicon amendment to rice plants impairs sucking behaviors and population growth in the phloem feeder Nilaparvata lugens (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 1101. [Google Scholar] [CrossRef]
- Lu, Z.; Yu, X.; Hu, C. Predatory behavior of mirid bug, Cyrthorhinus lividipennis, on rice plants with different nitrogen regimes. Int. Rice Res. Notes 2004, 29, 38–40. [Google Scholar]
- Watanabe, T.; Kitagawa, H. Photosynthesis and translocation of assimilates in rice plants following phloem feeding by the planthopper Nilaparvata lugens (Homoptera: Delphacidae). J. Econ. Entomol. 2000, 93, 1192–1198. [Google Scholar] [CrossRef]
- Wu, J.-C.; Qiu, H.-M.; Yang, G.-Q.; Dong, B.; Gu, H. Nutrient uptake of rice roots in response to infestation of Nilaparvata lugens (Stål) (Homoptera: Delphacidae). J. Econ. Entomol. 2003, 96, 1798–1804. [Google Scholar] [CrossRef]
- Rubia-Sanchez, E.; Suzuki, Y.; Arimura, K.; Miyamoto, K.; Matsumura, M.; Watanabe, T. Comparing Nilaparvata lugens (Stal) and Sogatella furcifera (Horvath) (Homoptera: Delphacidae) feeding effects on rice plant growth processes at the vegetative stage. Crop. Prot. 2003, 22, 967–974. [Google Scholar] [CrossRef]
- Hongoh, Y.; Ishikawa, H. Uric acid as a nitrogen resource for the brown planthopper, Nilaparvata lugens: Studies with synthetic diets and aposymbiotic insects. Zool. Sci. 1997, 14, 581–586. [Google Scholar] [CrossRef]
- Sasaki, T.; Kawamura, M.; Ishikawa, H. Nitrogen recycling in the brown planthopper, Nilaparvata lugens: Involvement of yeast-like endosymbionts in uric acid metabolism. J. Insect Physiol. 1996, 42, 125–129. [Google Scholar] [CrossRef]
- Room, P.M.; Julien, M.H.; Forno, I.W. Vigorous plants suffer most from herbivores: Latitude, nitrogen and biological control of the weed Salvinia molesta. Oikos 1989, 54, 92–100. [Google Scholar] [CrossRef]
- Hunt, R. Plant growth analysis: Second derivatives and compounded second derivatives of splined plant growth curves. Ann. Bot. 1982, 50, 317–328. [Google Scholar] [CrossRef]
- Alam, S.N.; Cohen, M.B. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a doubled-haploid rice population. Theor. Appl. Genet. 1998, 97, 1370–1379. [Google Scholar] [CrossRef]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia-Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Panda, N.; Heinrichs, E.A.; Hibino, H. Resistance of the rice variety Utri Rajapan to ragged stunt and tungro viruses. Crop. Prot. 1984, 3, 491–500. [Google Scholar] [CrossRef]
- Crisol, E.; Almazan, M.L.P.; Jones, P.W.; Horgan, F.G. Planthopper-rice interactions: Unequal stresses on pure-line and hybrid rice under similar experimental conditions. Entomol. Exp. Appl. 2013, 147, 18–32. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol, E. Hybrid rice and insect herbivores in Asia. Entomol. Exp. Appl. 2013, 148, 1–19. [Google Scholar] [CrossRef]
- Chen, J.; Yu, X.; Cheng, J. Evaluation for tolerance characteristics of different rice varieties to brown planthopper (BPH), Nilaparvata lugens Stål. Acta Agron. Sin. 2009, 35, 795–801. [Google Scholar] [CrossRef]
- Kuang, Y.-H.; Fang, Y.-F.; Lin, S.-C.; Tsai, S.-F.; Yang, Z.-W.; Li, C.-P.; Huang, S.-H.; Hechanova, S.L.; Jena, K.K.; Chuang, W.-P. The impact of climate change on the resistance of rice near-isogenic lines with resistance genes against brown planthopper. Rice 2021, 14, 64. [Google Scholar] [CrossRef]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Temperature-dependent oviposition and nymph performance reveal distinct thermal niches of coexisting planthoppers with similar thresholds for development. PLoS ONE 2020, 15, e0235506. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Elevated temperatures diminish the effects of a highly resistant rice variety on the brown planthopper. Sci. Rep. 2021, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.-J.; Xu, H.-X.; Zheng, X.-S.; Fu, Q.; Lu, Z.-X. High temperature modifies resistance performances of rice varieties to brown planthopper, Nilaparvata lugens (Stål). Rice Sci. 2010, 17, 334–338. [Google Scholar] [CrossRef]
- Cohen, M.B.; Alam, S.N.; Medina, E.B.; Bernal, C.C. Brown planthopper, Nilaparvata lugens, resistance in rice cultivar IR64: Mechanism and role in successful N. lugens management in Central Luzon, Philippines. Entomol. Exp. Appl. 1997, 85, 221–229. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variety | Nitrogen 1 | Xylem as Proportion of Total Honeydew 2 | Development to Adult Stage (Proportion) 2 | Adult Females (Proportion) 2 | Nymph Biomass (mg Dry Weight) 2 | Eggs Per Plant 2 |
|---|---|---|---|---|---|---|
| IR62 | 1 | 0.89 ± 0.11 ef | 0.25 ± 0.14 a | 0.75 ± 0.14 | 0.48 ± 0.25 ab | 141.00 ± 31.01 a–c |
| 2 | 0.24 ± 0.18 | 0.42 ± 0.22 | 0.15 ± 0.10 | 1.78 ± 0.20 | 111.67 ± 7.22 | |
| 3 | 0.73 ± 0.16 | 0.35 ± 0.05 | 0.67 ± 0.33 | 2.45 ± 0.38 | 144.00 ± 40.25 | |
| ADR52 | 1 | 0.81 ± 0.09 c–f | 0.69 ± 0.06 ab | 0.53 ± 0.24 | 2.06 ± 0.27 a–c | 93.67 ± 29.63 a–c |
| 2 | 0.67 ± 0.33 | 0.81 ± 0.10 | 0.28 ± 0.15 | 1.29 ± 0.11 | 166.33 ± 22.70 | |
| 3 | 0.33 ± 0.12 | 0.47 ± 0.14 | 0.44 ± 0.29 | 1.51 ± 0.25 | 206.33 ± 84.94 | |
| IR60 | 1 | 0.48 ± 0.25 c–f | 0.33 ± 0.33 a–c | 1.00 ± 0.00 | 1.02 ± 0.34 a | 110.33 ± 29.69 ab |
| 2 | 0.79 ± 0.11 | 0.62 ± 0.22 | 0.61 ± 0.20 | 1.47 ± 0.45 | 89.00 ± 25.36 | |
| 3 | 0.46 ± 0.07 | 0.88 ± 0.06 | 0.47 ± 0.03 | 1.78 ± 0.35 | 158.33 ± 38.67 | |
| IR65482-4 | 1 | 0.77 ± 0.13 f | 0.42 ± 0.30 a–d | 0.42 ± 0.22 | 1.01 ± 0.48 ab | 47.67 ± 15.77 a–d |
| 2 | 0.97 ± 0.03 | 0.77 ± 0.15 | 0.28 ± 0.15 | 1.39 ± 0.23 | 102.67 ± 16.71 | |
| 3 | 0.67 ± 0.16 | 0.92 ± 0.08 | 0.57 ± 0.05 | 1.92 ± 0.13 | 180.33 ± 43.84 | |
| Pokkali | 1 | 0.22 ± 0.10 a–e | 1.00 ± 0.00 d | 0.33 ± 0.19 | 2.18 ± 0.80 a–c | 111.67 ± 31.64 ab |
| 2 | 0.47 ± 0.21 | 1.00 ± 0.00 | 0.17 ± 0.08 | 1.16 ± 0.23 | 69.33 ± 25.56 | |
| 3 | 0.38 ± 0.14 | 1.00 ± 0.00 | 0.62 ± 0.22 | 1.70 ± 0.61 | 157.67 ± 30.07 | |
| Swarnalata | 1 | 0.94 ± 0.06 d–f | 0.81 ± 0.10 b–d | 0.92 ± 0.08 | 2.61 ± 0.21 cd | 131.00 ± 37.31 a–d |
| 2 | 0.40 ± 0.31 | 0.83 ± 0.17 | 0.80 ± 0.12 | 3.15 ± 0.33 | 222.33 ± 32.84 | |
| 3 | 0.62 ± 0.10 | 0.94 ± 0.06 | 0.45 ± 0.03 | 3.39 ± 0.21 | 173.00 ± 10.79 | |
| Triveni | 1 | 0.28 ± 0.21 a–d | 0.48 ± 0.08 a–d | 1.00 ± 0.00 | 2.31 ± 0.36 a–d | 58.33 ± 13.53 a |
| 2 | 0.23 ± 0.03 | 0.72 ± 0.15 | 0.56 ± 0.06 | 1.99 ± 0.22 | 96.00 ± 24.11 | |
| 3 | 0.45 ± 0.29 | 0.94 ± 0.06 | 0.34 ± 0.09 | 2.32 ± 1.46 | 100.67 ± 5.36 | |
| IR82396 H | 1 | 0.32 ± 0.10 b–f | 0.83 ± 0.08 b–d | 0.29 ± 0.04 | 1.68 ± 0.13 a–d | 93.00 ± 29.14 a–c |
| 2 | 0.44 ± 0.12 | 1.00 ± 0.00 | 0.56 ± 0.12 | 2.17 ± 0.45 | 173.33 ± 13.92 | |
| 3 | 0.64 ± 0.10 | 1.00 ± 0.00 | 0.28 ± 0.15 | 2.05 ± 0.54 | 125.33 ± 41.90 | |
| IR82391 H | 1 | 0.41 ± 0.09 a–d | 0.94 ± 0.06 d | 0.87 ± 0.13 | 4.61 ± 0.06 d | 66.33 ± 14.62 a–d |
| 2 | 0.30 ± 0.07 | 1.00 ± 0.00 | 0.67 ± 0.09 | 3.00 ± 0.67 | 203.00 ± 64.14 | |
| 3 | 0.27 ± 0.06 | 1.00 ± 0.00 | 0.45 ± 0.03 | 3.18 ± 0.20 | 224.67 ± 17.98 | |
| IR46 | 1 | 0.28 ± 0.10 a–c | 0.78 ± 0.22 b–d | 0.42 ± 0.30 | 2.35 ± 0.46 a–d | 121.67 ± 48.70 a–d |
| 2 | 0.21 ± 0.10 | 1.00 ± 0.00 | 0.43 ± 0.03 | 2.52 ± 0.33 | 215.33 ± 37.23 | |
| 3 | 0.21 ± 0.13 | 0.85 ± 0.08 | 0.19 ± 0.10 | 1.49 ± 0.19 | 149.67 ± 35.80 | |
| TKM6 | 1 | 0.16 ± 0.07 a | 0.87 ± 0.13 b–d | 0.33 ± 0.17 | 1.64 ± 0.28 a–d | 199.67 ± 22.26 a–d |
| 2 | 0.07 ± 0.03 | 1.00 ± 0.00 | 0.50 ± 0.14 | 2.23 ± 0.77 | 158.67 ± 58.29 | |
| 3 | 0.09 ± 0.02 | 0.92 ± 0.08 | 0.56 ± 0.29 | 2.03 ± 0.47 | 145.33 ± 8.25 | |
| Utri Rajapan | 1 | 0.35 ± 0.19 ab | 1.00 ± 0.00 b–d | 0.65 ± 0.05 | 3.66 ± 0.39 a–d | 216.33 ± 37.43 a–d |
| 2 | 0.12 ± 0.09 | 1.00 ± 0.00 | 0.40 ± 0.31 | 1.77 ± 0.75 | 159.33 ± 33.53 | |
| 3 | 0.13 ± 0.10 | 0.87 ± 0.13 | 0.47 ± 0.24 | 1.95 ± 0.58 | 143.00 ± 15.62 | |
| IR40 | 1 | 0.39 ± 0.18 a–d | 1.00 ± 0.00 d | 0.62 ± 0.22 | 2.89 ± 0.58 a–d | 215.00 ± 29.67 a–d |
| 2 | 0.25 ± 0.04 | 1.00 ± 0.00 | 0.67 ± 0.17 | 3.40 ± 0.51 | 207.67 ± 57.78 | |
| 3 | 0.16 ± 0.07 | 1.00 ± 0.00 | 0.63 ± 0.19 | 2.75 ± 0.47 | 110.00 ± 37.04 | |
| ASD7 | 1 | 0.39 ± 0.16 ab | 0.93 ± 0.07 b–d | 0.67 ± 0.33 | 3.92 ± 1.48 cd | 168.00 ± 30.05 b–d |
| 2 | 0.15 ± 0.12 | 0.81 ± 0.19 | 0.47 ± 0.12 | 2.48 ± 0.19 | 189.00 ± 27.00 | |
| 3 | 0.03 ± 0.02 | 0.79 ± 0.21 | 0.73 ± 0.18 | 3.46 ± 0.43 | 270.00 ± 41.58 | |
| IR22 | 1 | 0.12 ± 0.03 a | 1.00 ± 0.00 b–d | 0.39 ± 0.14 | 1.69 ± 0.17 a–d | 263.33 ± 15.43 cd |
| 2 | 0.11 ± 0.04 | 1.00 ± 0.00 | 0.56 ± 0.08 | 3.05 ± 0.75 | 235.33 ± 41.30 | |
| 3 | 0.07 ± 0.02 | 0.76 ± 0.12 | 0.89 ± 0.11 | 3.25 ± 0.41 | 184.67 ± 3.18 | |
| TN1 | 1 | 0.09 ± 0.03 a | 0.93 ± 0.07 b–d | 0.55 ± 0.10 | 3.15 ± 0.84 cd | 234.33 ± 8.09 d |
| 2 | 0.01 ± 0.00 | 0.72 ± 0.19 | 0.78 ± 0.15 | 6.28 ± 1.74 | 271.00 ± 44.88 | |
| 3 | 0.02 ± 0.00 | 1.00 ± 0.00 | 0.13 ± 0.13 | 3.60 ± 1.75 | 272.00 ± 32.88 | |
| F-variety 3 | 10.045 *** | 6.636 *** | 1.519ns | 4.559 *** | 5.397 *** | |
| F-nitrogen 3 | 2.547ns | 2.448ns | 3.551 * | 0.024ns | 3.496 * | |
| F-interaction 3 | 1.075ns | 1.090ns | 1.718ns | 1.650 * | 1.830 * |
| Variety | Nitrogen 1 | Shoot Biomass (g Dry Weight) 2 | Root Biomass (g Dry Weight) 2 | Growth Rate (g Dry Weight Day−1) 2 | Weight Loss (g) 2 | Weight Loss (g mg − Insect−1) 2 |
|---|---|---|---|---|---|---|
| IR82391 H | 1 | 0.70 ± 0.07 a | 0.44 ± 0.09 a | 0.07 ± 0.00 a | −0.47 ± 0.32 a | −0.10 ± 0.07 b |
| 2 | 1.31 ± 0.21 | 0.77 ± 0.10 | 0.12 ± 0.02 | −0.58 ± 0.26 | −0.25 ± 0.16 | |
| 3 | 2.34 ± 0.06 | 1.08 ± 0.03 | 0.20 ± 0.00 | −1.26 ± 0.09 | −0.40 ± 0.02 | |
| IR40 | 1 | 0.59 ± 0.04 a | 0.41 ± 0.04 ab | 0.04 ± 0.01 ab | −0.35 ± 0.06 a | −0.13 ± 0.04 b |
| 2 | 1.90 ± 0.43 | 1.23 ± 0.94 | 0.19 ± 0.09 | −1.48 ± 1.18 | −0.37 ± 0.26 | |
| 3 | 1.96 ± 0.11 | 0.87 ± 0.16 | 0.16 ± 0.01 | −0.81 ± 0.28 | −0.34 ± 0.14 | |
| ASD7 | 1 | 1.42 ± 0.46 b–f | 0.90 ± 0.14 a–e | 0.12 ± 0.04 a–c | −0.94 ± 0.53 a–c | −0.20 ± 0.07 b |
| 2 | 2.74 ± 0.82 | 1.45 ± 0.24 | 0.19 ± 0.05 | −1.48 ± 1.53 | −0.52 ± 0.61 | |
| 3 | 3.03 ± 0.52 | 1.64 ± 0.41 | 0.24 ± 0.05 | −1.58 ± 0.47 | −0.44 ± 0.08 | |
| IR22 | 1 | 0.90 ± 0.49 ab | 0.62 ± 0.22 ab | 0.09 ± 0.05 a–c | −0.81 ± 0.95 a–c | −0.55 ± 0.59 b |
| 2 | 1.54 ± 0.31 | 0.88 ± 0.20 | 0.15 ± 0.03 | −0.16 ± 0.45 | −0.17 ± 0.25 | |
| 3 | 2.93 ± 0.01 | 1.73 ± 0.31 | 0.29 ± 0.03 | −2.53 ± 0.16 | −0.82 ± 0.17 | |
| TKM6 | 1 | 1.04 ± 0.13 a–d | 0.72 ± 0.06 ab | 0.10 ± 0.01 a–c | −0.60 ± 0.68 ab | −0.39 ± 0.44 b |
| 2 | 2.03 ± 0.07 | 1.28 ± 0.20 | 0.19 ± 0.02 | −1.76 ± 0.15 | −0.99 ± 0.30 | |
| 3 | 2.36 ± 0.30 | 0.95 ± 0.22 | 0.19 ± 0.03 | −0.75 ± 0.91 | −0.51 ± 0.64 | |
| Swarnalata | 1 | 1.79 ± 0.24 c–f | 1.26 ± 0.17 b–e | 0.16 ± 0.02 b–d | −1.55 ± 0.62 a–c | −0.64 ± 0.30 ab |
| 2 | 2.91 ± 0.50 | 1.71 ± 0.29 | 0.27 ± 0.05 | −2.06 ± 0.98 | −0.72 ± 0.39 | |
| 3 | 3.86 ± 0.27 | 1.75 ± 0.21 | 0.32 ± 0.04 | −2.73 ± 0.33 | −0.82 ± 0.14 | |
| IR65482−4 | 1 | 0.71 ± 0.01 a–e | 0.49 ± 0.05 a–c | 0.06 ± 0.00 a–c | −0.27 ± 0.12 a–c | −0.51 ± 0.26 ab |
| 2 | 2.13 ± 0.13 | 1.14 ± 0.16 | 0.18 ± 0.01 | −1.06 ± 0.38 | −0.72 ± 0.15 | |
| 3 | 3.53 ± 0.43 | 1.81 ± 0.17 | 0.30 ± 0.00 | −2.14 ± 0.39 | −1.10 ± 0.14 | |
| TN1 | 1 | 2.27 ± 0.23 d–f | 1.60 ± 0.33 c–e | 0.22 ± 0.05 cd | −2.26 ± 0.83 a–c | −1.08 ± 0.70 ab |
| 2 | 2.56 ± 0.12 | 1.50 ± 0.00 | 0.22 ± 0.01 | −1.95 ± 0.41 | −0.40 ± 0.19 | |
| 3 | 3.49 ± 0.81 | 2.58 ± 0.21 | 0.34 ± 0.07 | −3.19 ± 1.49 | −0.95 ± 0.50 | |
| IR46 | 1 | 1.45 ± 0.37 b–f | 0.87 ± 0.07 a–d | 0.13 ± 0.03 a–c | −1.44 ± 0.35 a–c | −0.64 ± 0.16 ab |
| 2 | 3.71 ± 0.34 | 1.63 ± 0.16 | 0.31 ± 0.03 | −3.02 ± 0.68 | −1.22 ± 0.28 | |
| 3 | 2.70 ± 0.43 | 1.45 ± 0.15 | 0.22 ± 0.04 | −0.88 ± 0.64 | −0.58 ± 0.38 | |
| ADR52 | 1 | 1.79 ± 0.24 fg | 1.15 ± 0.27 a–e | 0.16 ± 0.04 cd | −1.07 ± 0.11 a–c | −0.54 ± 0.08 ab |
| 2 | 3.17 ± 0.23 | 1.28 ± 0.10 | 0.23 ± 0.01 | −0.64 ± 0.33 | −0.52 ± 0.30 | |
| 3 | 5.42 ± 0.14 | 1.98 ± 0.16 | 0.40 ± 0.01 | −3.13 ± 1.22 | −1.93 ± 0.42 | |
| IR60 | 1 | 0.79 ± 0.31 a–e | 0.47 ± 0.17 a–d | 0.05 ± 0.02 a–c | −0.41 ± 0.51 a–c | −0.40 ± 0.70 ab |
| 2 | 2.40 ± 0.23 | 1.28 ± 0.28 | 0.20 ± 0.04 | −1.43 ± 0.22 | −1.28 ± 0.52 | |
| 3 | 3.35 ± 0.71 | 2.03 ± 0.55 | 0.29 ± 0.08 | −2.19 ± 1.04 | −1.61 ± 1.05 | |
| Pokkali | 1 | 1.41 ± 0.12 e–g | 0.86 ± 0.01 a–d | 0.12 ± 0.02 abc | −1.24 ± 0.22 a–c | −0.69 ± 0.17 ab |
| 2 | 3.12 ± 0.95 | 1.65 ± 0.58 | 0.22 ± 0.08 | −2.00 ± 0.76 | −1.70 ± 0.42 | |
| 3 | 4.36 ± 0.16 | 1.17 ± 0.07 | 0.32 ± 0.02 | −2.13 ± 0.91 | −1.10 ± 0.22 | |
| Utri Rajapan | 1 | 2.76 ± 0.22 g | 1.52 ± 0.18 e | 0.24 ± 0.01 d | −2.87 ± 0.06 c | −0.80 ± 0.08 ab |
| 2 | 4.45 ± 0.67 | 2.01 ± 0.04 | 0.37 ± 0.04 | −2.82 ± 0.63 | −0.96 ± 0.17 | |
| 3 | 4.84 ± 0.32 | 3.17 ± 0.49 | 0.44 ± 0.03 | −3.70 ± 0.34 | −2.15 ± 0.46 | |
| IR62 | 1 | 1.24 ± 0.22 a–c | 0.92 ± 0.27 a–d | 0.14 ± 0.03 a–c | −1.67 ± 0.31 a–c | −1.26 ± 0.15 ab |
| 2 | 1.47 ± 0.27 | 1.65 ± 0.36 | 0.20 ± 0.04 | −1.96 ± 0.51 | −1.18 ± 0.42 | |
| 3 | 2.72 ± 1.09 | 1.85 ± 0.59 | 0.28 ± 0.10 | −4.08 ± 1.45 | −1.76 ± 0.68 | |
| IR82396 H | 1 | 1.28 ± 0.30 b–f | 1.03 ± 0.18 a–e | 0.13 ± 0.03 cd | −1.68 ± 0.49bc | −1.04 ± 0.36 ab |
| 2 | 3.56 ± 0.57 | 2.21 ± 0.77 | 0.37 ± 0.07 | −4.90 ± 1.45 | −2.45 ± 0.98 | |
| 3 | 3.46 ± 0.64 | 1.79 ± 0.32 | 0.31 ± 0.06 | −2.67 ± 0.71 | −1.35 ± 0.37 | |
| Triveni | 1 | 1.81 ± 0.28 e–g | 1.47 ± 0.27 de | 0.16 ± 0.02 cd | −1.98 ± 0.09 a–c | −0.89 ± 0.11 a |
| 2 | 2.76 ± 0.14 | 2.28 ± 0.03 | 0.26 ± 0.02 | −2.34 ± 0.15 | −1.22 ± 0.22 | |
| 3 | 4.23 ± 0.50 | 2.37 ± 0.39 | 0.40 ± 0.06 | −4.07 ± 0.98 | −4.55 ± 2.44 | |
| F−variety 3 | 11.225 *** | 6.608 *** | 6.174 *** | 3.655 *** | 2.974 *** | |
| F−nitrogen 3 | 99.596 *** | 37.459 *** | 66.639 *** | 10.269 *** | 6.327 *** | |
| F−interaction 3 | 1.365ns | 1.029ns | 1.136ns | 1.171ns | 1.323ns |
| Model | Slope | Intercept | R2 | F1,15 1 |
|---|---|---|---|---|
| Weight loss | ||||
| 0 added nitrogen | −13.007 | 0.385 | 0.90 | 127.269 *** |
| 80 Kg N ha−1 | −13.463 | 1.242 | 0.74 | 38.847 *** |
| 150 Kg N ha−1 | −11.634 | 1.053 | 0.71 | 33.592 *** |
| Weight loss per mg insect | ||||
| 0 added nitrogen | −4.038 | −0.115 | 0.43 | 10.559 ** |
| 80 Kg N ha−1 | −5.244 | 0.288 | 0.41 | 9.219 ** |
| 150 Kg N ha−1 | −7.417 | 1.014 | 0.69 | 30.578 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horgan, F.G.; de Freitas, T.F.S.; Crisol-Martínez, E.; Mundaca, E.A.; Bernal, C.C. Nitrogenous Fertilizer Reduces Resistance but Enhances Tolerance to the Brown Planthopper in Fast-Growing, Moderately Resistant Rice. Insects 2021, 12, 989. https://doi.org/10.3390/insects12110989
Horgan FG, de Freitas TFS, Crisol-Martínez E, Mundaca EA, Bernal CC. Nitrogenous Fertilizer Reduces Resistance but Enhances Tolerance to the Brown Planthopper in Fast-Growing, Moderately Resistant Rice. Insects. 2021; 12(11):989. https://doi.org/10.3390/insects12110989
Chicago/Turabian StyleHorgan, Finbarr G., Thais Fernanda S. de Freitas, Eduardo Crisol-Martínez, Enrique A. Mundaca, and Carmencita C. Bernal. 2021. "Nitrogenous Fertilizer Reduces Resistance but Enhances Tolerance to the Brown Planthopper in Fast-Growing, Moderately Resistant Rice" Insects 12, no. 11: 989. https://doi.org/10.3390/insects12110989
APA StyleHorgan, F. G., de Freitas, T. F. S., Crisol-Martínez, E., Mundaca, E. A., & Bernal, C. C. (2021). Nitrogenous Fertilizer Reduces Resistance but Enhances Tolerance to the Brown Planthopper in Fast-Growing, Moderately Resistant Rice. Insects, 12(11), 989. https://doi.org/10.3390/insects12110989