
Immature Insect Assemblages from the Early Cretaceous (Purbeck/Wealden) of Southern England

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
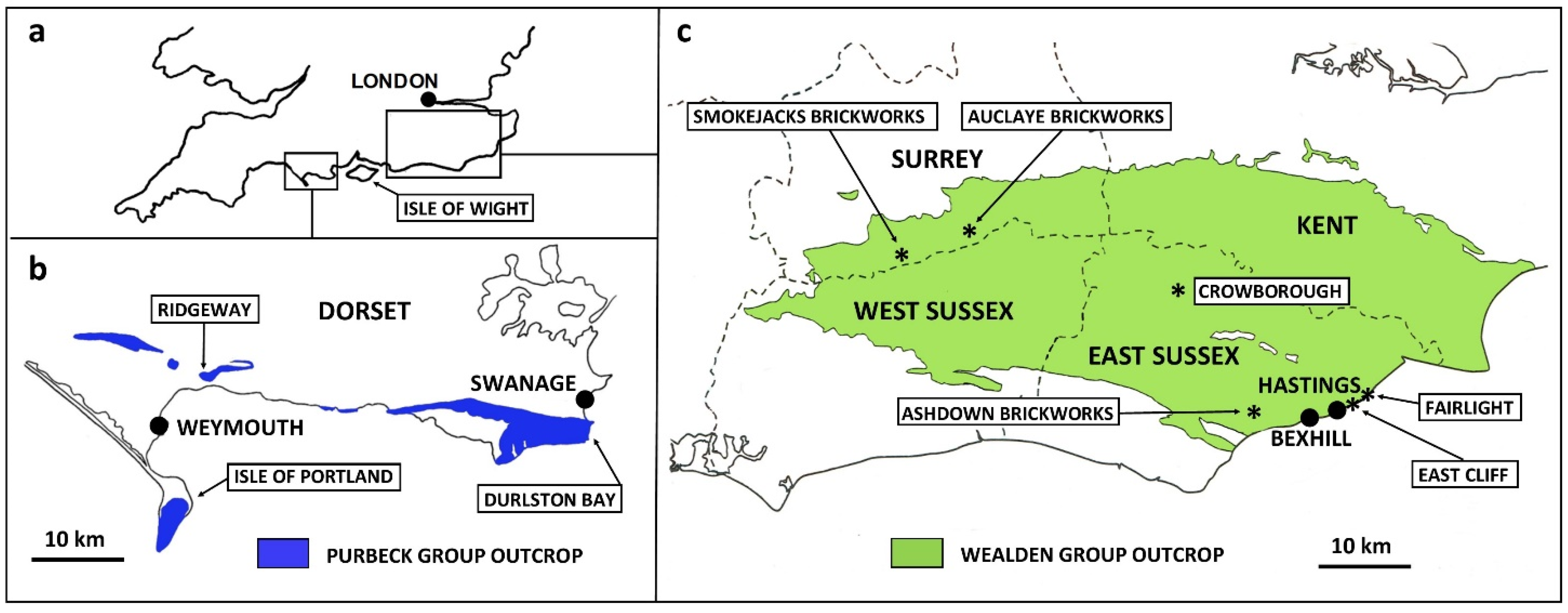
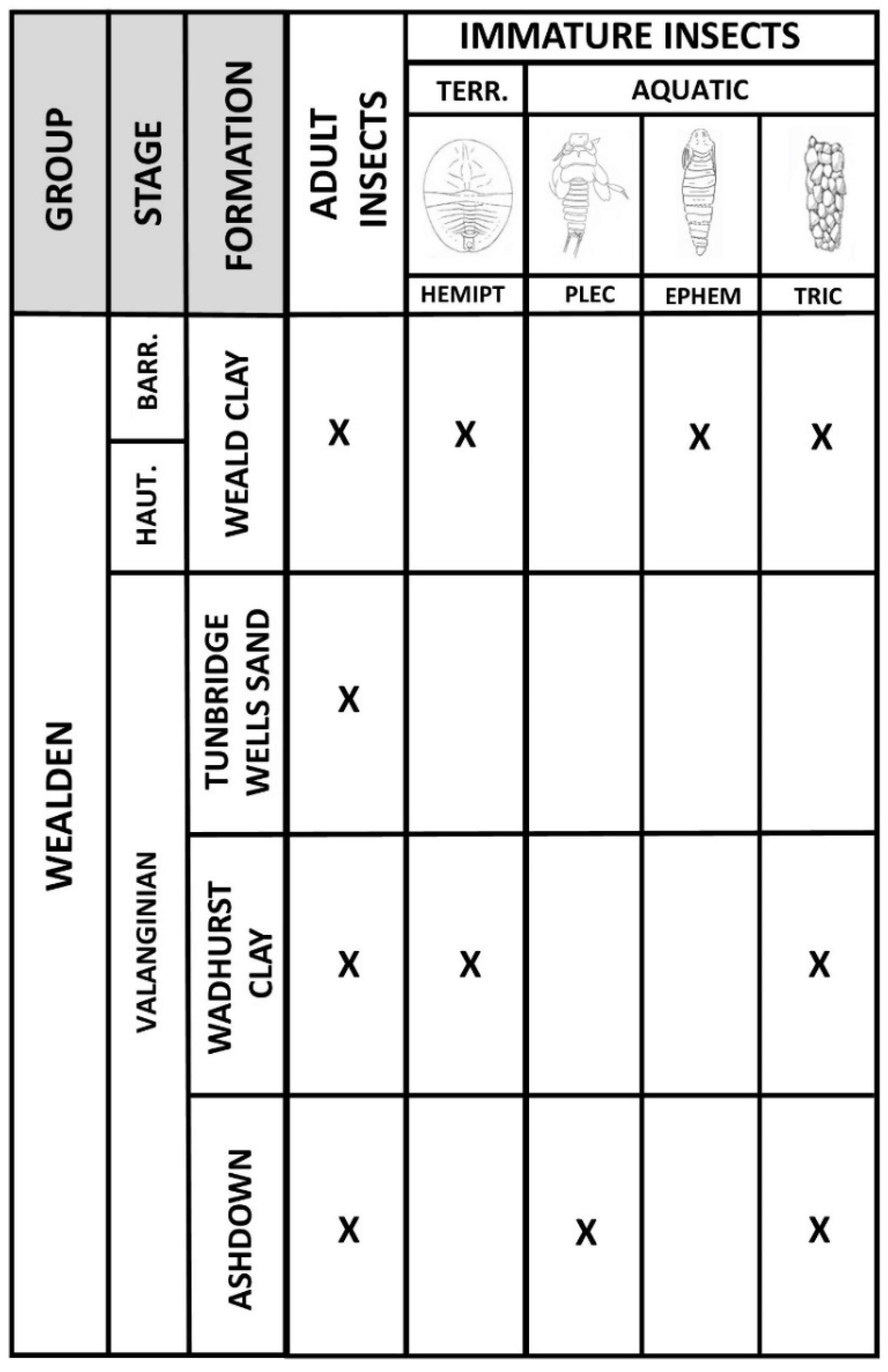
1. Introduction and Geological Setting
2. Materials and Methods
3. Results
3.1. Immature Terrestrial Insects
3.1.1. Introduction
3.1.2. Hemipteran Body Fossils
3.1.3. Trace Fossils
3.2. Immature Aquatic Insects—Purbeck Group
3.2.1. Introduction
3.2.2. Taxa Present
- Ephemeroptera (mayflies). No nymphal remains of Ephemeroptera have been reported; however, a subimaginal wing (Durlophlebia radleyi Sinitshenkova and Coram, 2002 [35]) from the Lulworth Formation of Durlston Bay represents the sole record of this order in the Purbeck. A mayfly subimago, popularly known as a ‘dun’, is a winged intermediate stage between nymph and adult that is capable of flight.
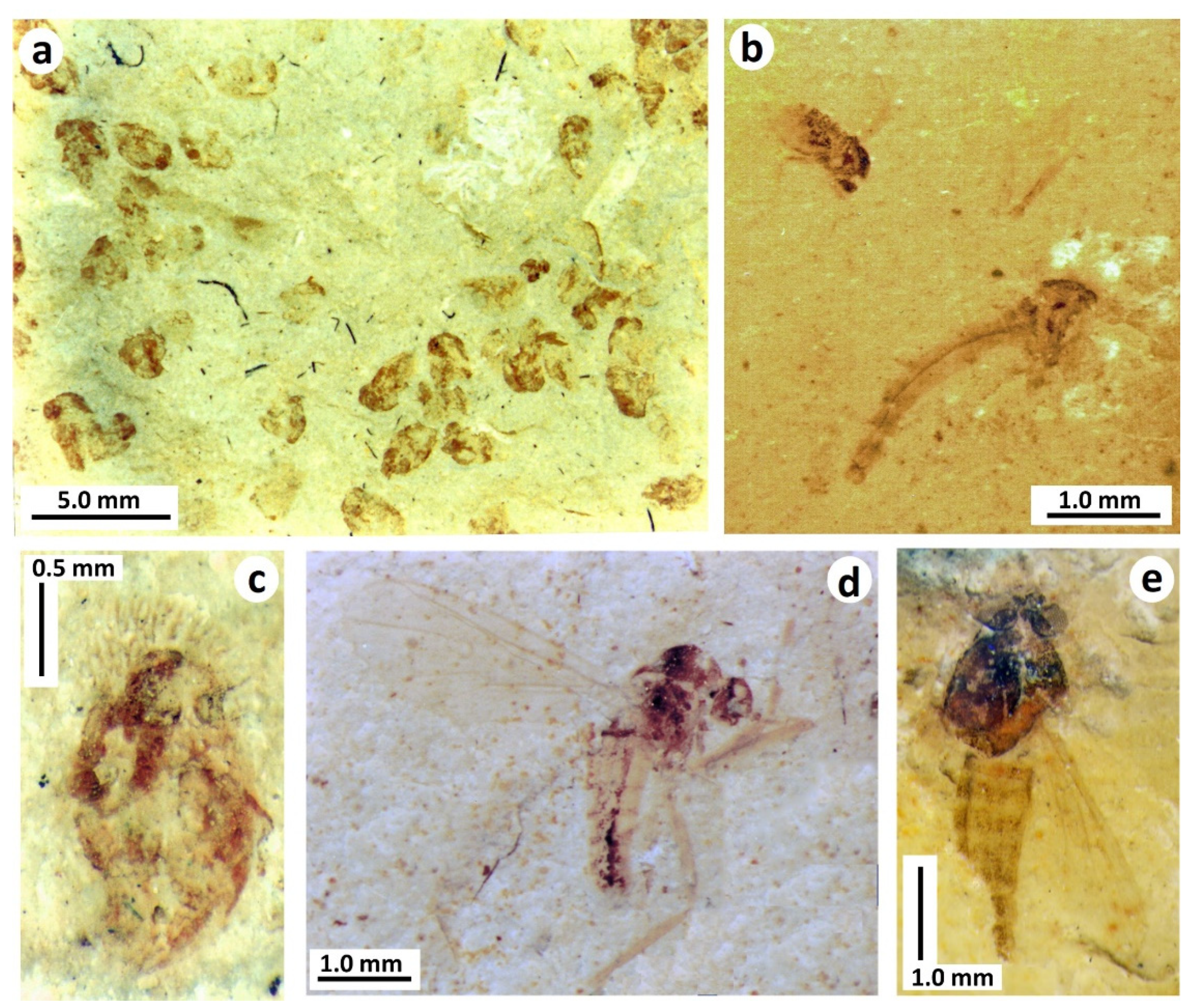
- Odonata (damselflies and dragonflies). Fossils of Odonata, mostly adult wings, are conspicuous and diverse elements of the Purbeck entomofauna, and include representatives of the two extant suborders Zygoptera and Epiprocta (including Anisoptera, or ‘true’ dragonflies), along with rare examples of extinct Archizygoptera and Tarsophlebioptera [17]. Fossils of nymphs (or their ecdysed cuticles) are much less common, and are usually indistinctly preserved, although often fairly intact [16,17] (Figure 8a,b). Among extant Odonata, the Purbeck fossils most resemble those of Anisoptera (which dominate the preserved adult fauna), being relatively wide-bodied and lacking evidence of the prominent caudal lamellae seen in Zygoptera. There are at least two morphotypes of nymph present, although it cannot be determined at present which taxa known from adult material they might belong to. Parallel discontinuous tracks observed rarely on bedding surfaces in a Durlston Formation horizon (DB175; Figure 8c) have been tentatively attributed, on the basis of their size and the presence of immature remains in the same bed, to odonatan naiads [30].
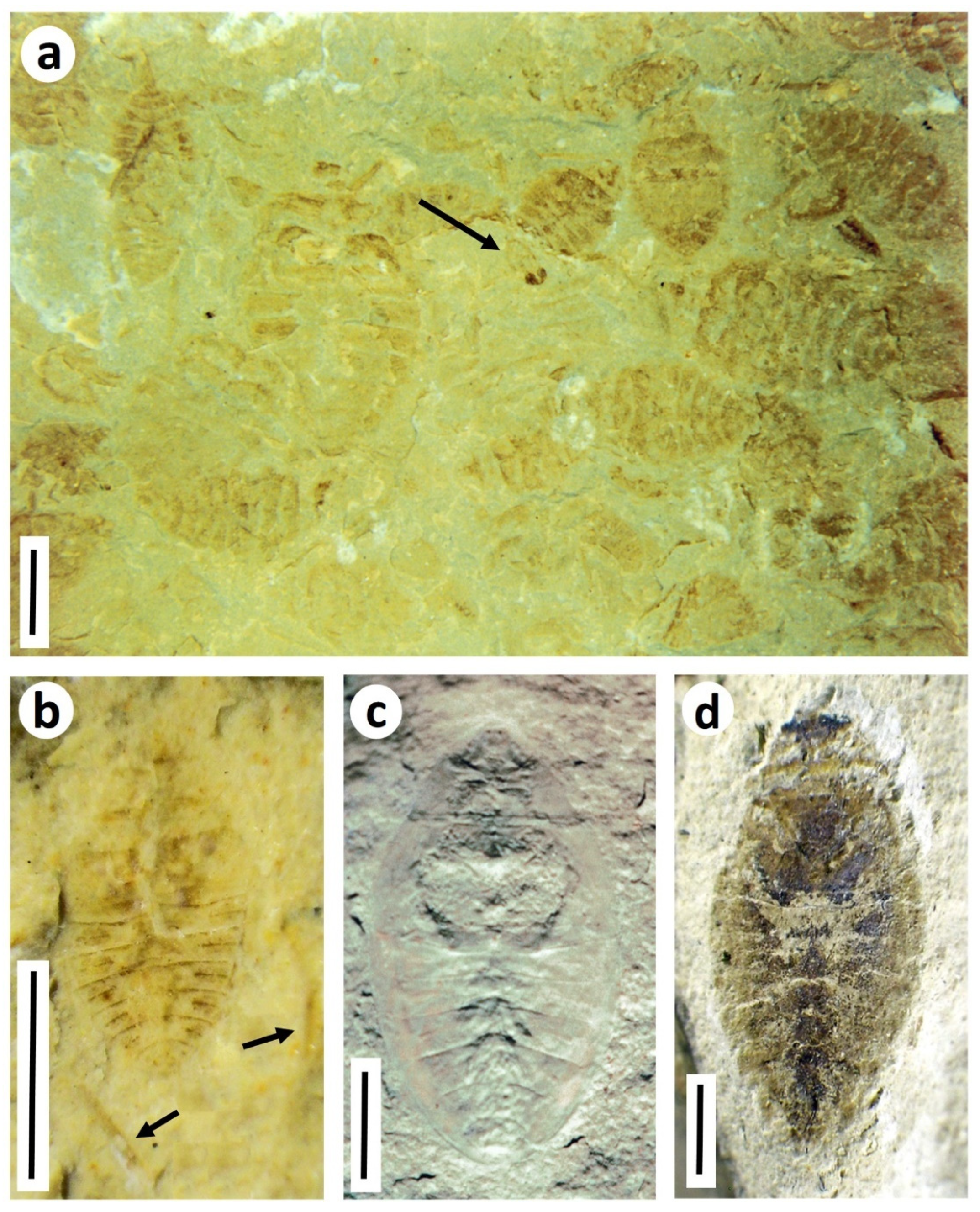
- Hemiptera (true bugs). Aquatic hemipteran nymphs can be commonly preserved in certain Purbeck horizons, occasionally massed on bedding planes (Figure 9a), reflecting mass mortality events or hydrodynamic concentration of ecdysed cuticles. All well-preserved specimens can be referred to Nepidium stolones Westwood, 1854 [36], winged adults of which commonly occur alongside the nymphs. Here we supplement the line drawings of selected life stages in Coram and Jarzembowski; Figure 6 [3] with photographs of specimens (Figure 9b–d). Nepidium Westwood, 1854 is of uncertain heteropteran family affinities, although well-preserved individuals show a distinctive pair of elongate ‘rowing legs’ in common with recent Notonectidae (backswimmers) and Corixidae (water boatmen).
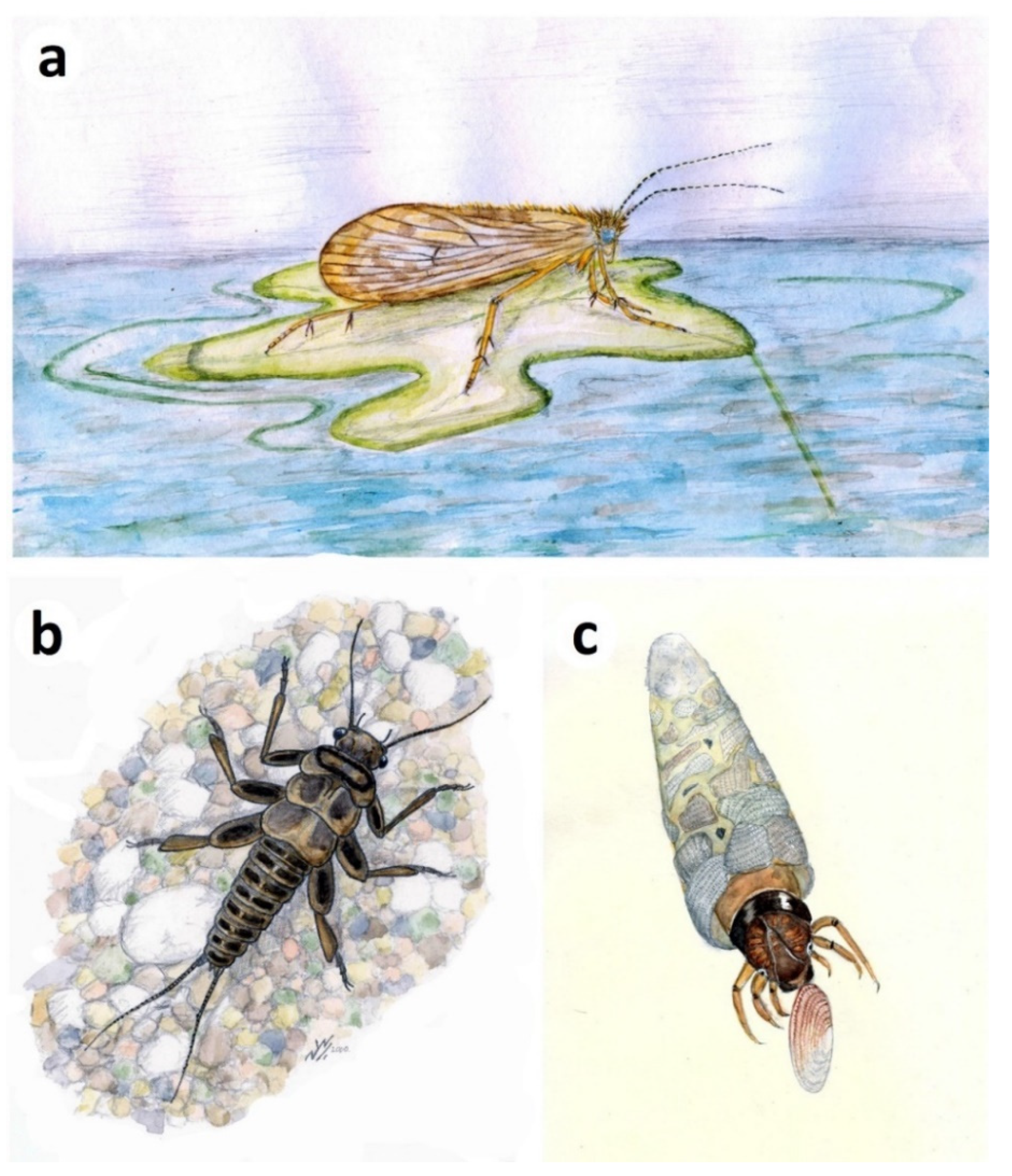
- Trichoptera (caddisflies). Larval cases, mostly composed of ostracod valves (igen. Ostracindusia Vialov, 1973 [37]) occur low in the Lulworth Formation of Durlston Bay and the Isle of Portland [3] (Figure 10a). Indistinct cases are also known from the Durlston Formation, but these appear to be composed of faecal pellets (Coprindusia Ivanov, 2006 [38]). Occasional fossils of exquisitely-preserved but crumpled-winged adult caddisflies from both Lulworth and Durlston formations are interpreted as newly pupated individuals which failed to emerge at the water surface [17,39,40]. Some, in fact, show what appear to be remnants of the pupal cases at the rear of the abdomen (Figure 10b). Eleven species of Trichoptera have been described from the Purbeck on the basis of adult wings, the fossils being numerically dominated by members of the stem genus Purbimodus Sukatsheva and Jarzembowski, 2001 [41], which survived into the Wealden. The presumed teneral caddisflies, despite their often fine preservation, cannot be linked to known taxa because the crumpled wings obscure important venational details. In other respects, however, they are morphologically indistinguishable. Intraspecific variation in choice of case building material in response to local availability has been documented in recent caddisflies [42], raising the possibility, as suggested by Coram and Jarzembowski [3], that a single opportunistic Purbeck species may have produced cases assignable to two different ichnogenera, making use of whatever case-building materials were available (ostracods being largely absent from the horizon with faecal-pellet cases).
- Diptera (true flies). Small aquatic pupae of nematoceran Diptera can be abundant at certain levels (Figure 11a–c). They can also be accompanied by adult flies which usually have indistinct crumpled wings, suggesting that these were newly pupated individuals that failed to emerge from the water, as suggested for the caddisflies above. Their taxonomic identity is uncertain, however, at least some and perhaps all are likely to be chironomids or chaoborids since both families are also known from well-preserved adult specimens that presumably emerged locally (Figure 11d,e). A chironomid identity is probably more likely since recent chaoborids, in contrast to chironomids, are intolerant of elevated salinities [43] and thus are less likely to have thrived in the saline Purbeck waters the pupae are preserved in (see discussion below). The total number of preserved immature dipteran species cannot be determined at present, but overall diversity may have been low since there is limited morphological diversity evident. Nevertheless, other nematoceran families known from adult material could be present, including small Limoniidae (craneflies, common in the Purbeck) and Ceratopogonidae (biting midges, known from a single Purbeck wing [44]). The pupa in Figure 11c uniquely bears hair-like respiratory filaments on its head, which, along with its general body proportions most closely resemble Simuliidae (black flies) among recent Diptera with aquatic development. Simuliidae, which can tolerate brackish salinities [45], have a fossil record extending back to the Jurassic [46] although no adult wings have to date been recognised in the Purbeck.
3.3. Immature Aquatic Insects—Wealden Group
3.3.1. Taxa Present
- Plecoptera (stoneflies). The lower Wealden has produced a unique plecopteran nymph (Figure 12b). Ecdyoperla fairlightensis Sinitshenkova, 1998 [12] was recovered from the Fairlight clay facies in the lower Ashdown Formation of Fairlight, near Hastings (Figure 1 and Figure 3). It is the only European Cretaceous representative of this order recorded to date and the family placement is problematic.
- Ephemeroptera (mayflies). The upper Weald Clay Formation of Smokejacks brickworks, Surrey, has produced a unique mayfly nymph, Schistonotorum wallisi Jarzembowski and Wang, 2019 [51]. Adult wings are unknown. Sedimentary burrows of the ichnogenus Rhizocorallium Zenker, 1836 [52] from the lower Wealden Wadhurst Clay Formation have been attributed to burrowing mayfly nymphs [53].
- Trichoptera (caddisflies). The lower Wealden river-channel deposits of the upper Ashdown (Sand) Formation at Hastings [54] have produced a diverse trichopteran indusifauna characterised by the extinct ichnogenus Conchindusia Vialov and Sukatsheva, 1976 [55], utilising ‘conchostracan’ (clam shrimp/spinicaudatan) carapaces (Figure 12c), and for which there appears to be no exact modern analogue. A comparatively minor channel deposit, the Northiam Sandstone, in the overlying Wadhurst Clay Formation has recently yielded an indusifauna reported below. Caddisfly cases also occur sporadically in the upper Wealden, small numbers being found in the Weald Clay Formation; amongst these may be mentioned the ichnogenus Piscindusia Jarzembowski, 1995 [50], also reported from scour fills in the Wadhurst Clay Formation and the Vectis Formation (coeval with the youngest Weald Clay) of the Isle of Wight [4], as there appears to be no modern analogue incorporating fish scales/bones in case construction, any more than clam shrimps [4].
3.3.2. Northiam Sandstone Indusifauna
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.; Ogg, G. Geologic Time Scale 2020, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; p. 1390. [Google Scholar]
- Hopson, P.M.; Wilkinson, I.P.; Woods, M.A. A Stratigraphical Framework for the Lower Cretaceous of England; British Geological Survey Research Report RR/08/03; British Geological Survey: Nottingham, UK, 2008. [Google Scholar]
- Coram, R.; Jarzembowski, E.A. Diversity and ecology of fossil insects in the Dorset Purbeck succession, southern England. In Life and Environments in Purbeck Times; Milner, A.R., Batten, D.J., Eds.; Palaeontological Association: London, UK, 2002; pp. 257–268. [Google Scholar]
- Jarzembowski, E.A. Insects. In English Wealden Fossils; Batten, D.J., Ed.; Palaeontological Association: London, UK, 2011; pp. 138–173. [Google Scholar]
- Ogg, J.G.; Hasenyager, R.W.; Wimbledon, W.A. Jurassic-Cretaceous boundary: Portland-Purbeck magnetostratigraphy and possible correlation to the Tethyan faunal realm. Geobios 1994, 17, 519–527. [Google Scholar] [CrossRef]
- Feist, M.; Lake, R.D.; Wood, C.J. Charophyte biostratigraphy of the Purbeck and Wealden of southern England. Palaeontology 1995, 38, 407–442. [Google Scholar]
- Schneider, A.C.; Heimhofer, U.; Heunisch, C.; Mutterlose, J. The Jurassic–Cretaceous boundary interval in non-marine strata of northwest Europe–new light on an old problem. Cretac. Res. 2018, 87, 42–54. [Google Scholar] [CrossRef]
- Allen, P. Purbeck-Wealden (early Cretaceous) climates. Proc. Geol. Assoc. 1998, 109, 197–236. [Google Scholar] [CrossRef]
- Clements, R.G. Type-section of the Purbeck Limestone Group, Durlston Bay, Swanage, Dorset. Proc. Dorset Nat. Hist. Arch. Soc. 1993, 114, 181–206. [Google Scholar]
- West, I.M. Evaporites and associated sediments of the basal Purbeck Formation (Upper Jurassic) of Dorset. Proc. Geol. Assoc. 1975, 86, 205–225. [Google Scholar] [CrossRef]
- Batten, D.J. Wealden geology. In English Wealden Fossils; Batten, D.J., Ed.; Palaeontological Association: London, UK, 2011; pp. 7–14. [Google Scholar]
- Sinitshenkova, N.D. The first European Cretaceous stonefly (Insecta: Perlida = Plecoptera). Cretac. Res. 1998, 19, 317–321. [Google Scholar] [CrossRef]
- Martin, J.H.; Camus, J.M. Whiteflies (Sternorrhyncha, Aleyrodidae) colonising ferns (Pteridophyta: Filicopsida, with descriptions of two new Trialeurodes and one new Metabemisia species from south-east Asia. Zootaxa 2001, 2, 1–19. [Google Scholar] [CrossRef]
- Martin, J.H.; Mifsud, D.; Rapisarda, C. The whiteflies (Hemiptera: Aleyrodidae) of Europe and the Mediterranean Basin. Bull. Entomol. Res. 2000, 90, 407–448. [Google Scholar] [CrossRef]
- Shcherbakov, D.E. The most primitive whiteflies (Hemiptera; Aleyrodidae; Bernaeinae subfam. nov.) from the Mesozoic of Asia and Burmese amber, with an overview of Burmese amber hemipterans. Bull. Nat. Hist. Mus. London Geol. 2000, 56, 29–37. [Google Scholar]
- Jarzembowski, E.A.; Coram, R.A. New fossil insect records from the Purbeck of Dorset and the Wealden of the Weald. Proc. Dorset Nat. Hist. Arch. Soc. 1997, 118, 119–124. [Google Scholar]
- Coram, R.A.; Jepson, J.E. Fossil Insects of the Purbeck Limestone Group of Southern England, 1st ed.; Siri Scientific Press: Manchester, UK, 2012; p. 144. [Google Scholar]
- Szwedo, J. The unity, diversity and conformity of bugs (Hemiptera) through time. Earth Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 109–128. [Google Scholar] [CrossRef]
- Handlirsch, A. Die Fossilen Insekten und die Phylogenie der Rezenten Formen, 1st ed.; Wilhelm Engelmann: Leipzig, Germany, 1908; p. 1430. [Google Scholar]
- Whalley, P.E.S.; Jarzembowski, E.A. Fossil insects from the Lithographic Limestone of Montsech (late Jurassic-early Cretaceous), Lérida Province, Spain. Bull. Brit. Mus. Nat. Hist. Geol. 1985, 38, 381–412. [Google Scholar]
- Popov, Y.A.; (Russian Academy of Sciences, Moscow, Russia). Personal communication, 1996.
- Jarzembowski, E.A. A boring beetle from the Wealden of the Weald. In Evolutionary Paleobiology of Behaviour and Coevolution; Boucot, A., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 373–376. [Google Scholar]
- Labandeira, C.C.; LePage, B.A.; Johnson, A.H. A Dendroctonus bark engraving (Coleoptera: Scolytidae) from a middle Eocene Larix (Coniferales: Pinaceae): Early or delayed colonization? Amer. J. Bot. 2001, 88, 2026–2039. [Google Scholar] [CrossRef]
- Needham, J.E. Forests of the Dinosaurs, 1st ed.; Hobnob Press: Salisbury, UK, 2011; p. 221. [Google Scholar]
- Lockwood, J.; Shears-Ozeki, C.; Martill, D.; Peaker, A. Life after Death: Subaerial and Subterranean Invertebrate Necrophagous Activity in Dinosaur Bone from the Wealden Group (Early Cretaceous), Isle of Wight, UK; University of Portsmouth: Portsmouth, UK; University of Liverpool: Liverpool, UK, 2016. [Google Scholar]
- Jarzembowski, E.A. Early Cretaceous insects from southern England. Modern Geol. 1984, 9, 71–93. [Google Scholar]
- Williams, D.D.; Feltmate, B.W. Aquatic Insects, 1st ed.; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Riek, E.F. The marine caddisfly Family Chathamiidae (Trichoptera). J. Aust. Entomol. Soc. 1976, 15, 405–419. [Google Scholar] [CrossRef]
- Haq, B.U.; Hardenbol, J.; Vail, P.R. Chronology of fluctuating sea levels since the Triassic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef]
- Coram, R.A. Palaeoecology And Palaeoenvironmental Significance of Early Cretaceous Fossil Insects from the Purbeck Limestone Group, Dorset, UK. Ph.D. Thesis, University of Reading, Reading, UK, 2005. [Google Scholar]
- Coram, R.A.; Radley, J.D. Revisiting climate change and palaeoenvironments in the Purbeck Limestone Group (Tithonian–Berriasian) of Durlston Bay, southern UK. Proc. Geol. Assoc. 2021, 132, 392–404. [Google Scholar] [CrossRef]
- Ponomarenko, A.G.; Coram, R.A.; Jarzembowski, E.A. New beetles (Insecta: Coleoptera) from the Berriasian Purbeck Limestone Group, Dorset, UK. Cretac. Res. 2005, 26, 277–281. [Google Scholar] [CrossRef]
- Popov, Y.A.; Coram, R.A.; Jarzembowski, E.A. Fossil heteropteran bugs from the Purbeck Limestone Group of Dorset. Proc. Dorset Nat. Hist. Arch. Soc. 1999, 120, 73–76. [Google Scholar]
- Burne, R.V.; Bauld, J.; De Deckker, P. Saline lake charophytes and their geological significance. J. Sedim. Petrol. 1980, 50, 281–293. [Google Scholar]
- Sinitshenkova, N.D.; Coram, R.A. The first mayfly from the Lower Cretaceous of southern England (Insecta: Ephemerida = Ephemeroptera). Cretac. Res. 2002, 23, 461–463. [Google Scholar] [CrossRef][Green Version]
- Westwood, J.O. Contributions to fossil entomology. Quart. J. Geol. Soc. Lond. 1854, 10, 378–396. [Google Scholar] [CrossRef]
- Vialov, O.S. Klasifikatsiya vikopnikh khatinok ruchainikiv (Trichoptera). Dopovidi Akad. Nauk. Ukr. Radyanskoi Sotsialistichnoi Resp. B 1973, B7, 585–588. [Google Scholar]
- Ivanov, V.D. Larvae of caddisflies (Insecta: Trichoptera) from the Mesozoic of Siberia. Paleontol. J. 2006, 40, 178–189. [Google Scholar] [CrossRef]
- Coram, R.; Jarzembowski, E.A.; Ross, A.J. New records of Purbeck fossil insects. Proc. Dorset Nat. Hist. Arch. Soc. 1995, 116, 146–150. [Google Scholar]
- Coram, R.A. Taphonomy and ecology of Purbeck fossil insects. Acta Zool. Crac. 2003, 46, 311–318. [Google Scholar]
- Sukatsheva, I.D.; Jarzembowski, E.A. Fossil caddisflies (Insecta: Trichoptera) from the Early Cretaceous of southern England II. Cretac. Res. 2001, 22, 685–694. [Google Scholar] [CrossRef]
- Otto, C.; Svensson, B.S. The significance of case material selection for the survival of caddis larvae. J. Anim. Ecol. 1980, 49, 855–865. [Google Scholar] [CrossRef]
- Grueber, W.B.; Bradley, T.J. The evolution of increased salinity tolerance in larvae of Aedes mosquitos: A phylogenetic analysis. Physiol. Zool. 1994, 67, 566–579. [Google Scholar] [CrossRef]
- Borkent, A.; Coram, R.A.; Jarzembowski, E.A. The oldest fossil biting midge (Diptera: Ceratopogonidae) from the Purbeck Limestone Group (Lower Cretaceous) of southern Great Britain. Polish J. Entomol. 2013, 82, 273–279. [Google Scholar] [CrossRef][Green Version]
- Zinchenko, T.D.; Golovatyuk, L.V. Salinity tolerance of macroinvertebrates in stream waters (review). Arid Ecosyt. 2013, 3, 113–121. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Sukhomlin, E.B. The first record of Late Cretaceous blackflies (Diptera: Simuliidae) in Asia (Yantardakh, Taimyr). Palentol. J. 2016, 50, 997–1000. [Google Scholar] [CrossRef]
- Coram, R.; Jarzembowski, E.A. Insect-bearing horizons in the type Purbeck and new Purbeck/Wealden flies (Diptera). Proc. Dorset Nat. Hist. Arch. Soc. 1998, 119, 135–140. [Google Scholar]
- Hill, C.R. A plant with flower-like organs from the Wealden of the Weald (Lower Cretaceous), southern England. Cretac. Res. 1996, 17, 27–38. [Google Scholar] [CrossRef]
- Jarzembowski, B.; Watson, N.; Jarzembowski, E. Wealden insects. An artist’s impression. Part III. Deposits 2017, 49, 10–11. [Google Scholar]
- Jarzembowski, E.A. Fossil caddisflies (Insecta: Trichoptera) from the Early Cretaceous of southern England. Cretac. Res. 1995, 16, 695–703. [Google Scholar] [CrossRef]
- Jarzembowski, E.A.; Wang, B. The first fossil mayfly nymph (Insecta: Ephemeroptera) from the British Isles. Proc. Geol. Assoc. 2019, 130, 673–676. [Google Scholar] [CrossRef]
- Zenker, J.C. Historisch-Topographisches Taschenbuch von Jena und Seiner Umgebung Besonders in Seiner Naturwissenschaftlicher und Medicinischer Beziehung; Wackenholder: Jena, Germany, 1836; p. 338. [Google Scholar]
- Pollard, J.E.; Radley, J.D. Trace fossils. In English Wealden Fossils; Batten, D.J., Ed.; Palaeontological Association: London, UK, 2011; pp. 652–676. [Google Scholar]
- Jarzembowski, E.A.; Sukatsheva, I.D.; Wang, B.; Zhang, H.; Zheng, D. A tubiculous perspective: New caddisfly cases (Insecta: Trichoptera) from the Lower Cretaceous of southern England and comparison with the Chinese indusifauna. Cretac. Res. 2016, 61, 44–56. [Google Scholar] [CrossRef]
- Vialov, O.S.; Sukatsheva, I.D. Iskopaemye domiki lichinok rucheinikov (Insecta, Trichoptera) Iikh ziachenie dlya stratigrafii. Tr. Sovmest. Sov. Mong. Paleontol. Ekspeditsiya 1976, 3, 169–232. [Google Scholar]
- Akinlotan, O. Multi-proxy approach to palaeoenvironmental modelling: The English Lower Cretaceous Weald Basin. Geol. J. 2017, 53, 316–335. [Google Scholar] [CrossRef]
- Allen, P. Wealden of the Weald: A new model. Proc. Geol. Assoc. 1975, 86, 389–437. [Google Scholar] [CrossRef]
- Hayward, R. The Geology of Part of the Wadhurst Clay Formation and Part of the Tunbridge Wells Formation at the Ashdown Brick Works, Bexhill, Sussex. BSc. Dissertation, University of Greenwich, London, UK, 1996. [Google Scholar]
- Berry, E.W. A new type of caddis case from the Lower Eocene of Tennessee. Proc. US Nat. Mus. 1927, 71, 1–4. [Google Scholar] [CrossRef]
- Sukatsheva, I.D.; Vassilenko, D.V. Caddisflies from Chernovskie Kopi (Jurassic/Cretaceous of Transbaikalia). Zoosymposia 2011, 5, 434–438. [Google Scholar] [CrossRef]
- Batten, D.J.; Austen, P.A. The Wealden of south-east England. In English Wealden Fossils; Batten, D.J., Ed.; Palaeontological Association: London, UK, 2011; pp. 15–51. [Google Scholar]
- Hartmann, J.D.W. System der Erd- und Flusschnecken der Schweiz, mit vergleichender Aufzählung aller auch in den benachbarten Ländern, Deutschland, Frankreich und Italien sich vorfindenen Arten. Neue Alp. 1821, 1, 194–268. [Google Scholar]
- Berger, V.Y.; Gorbushin, A.M. Tolerance and resistance in gastropod mollusks Hydrobia ulvae and H. ventrosa from the White Sea to abiotic environmental factors. Russ. J. Marine Biol. 2001, 27, 314–319. [Google Scholar] [CrossRef]
- Oertli, H.J. The Venice System for the classification of marine waters according to salinity. Pubbl. Stn. Zool. Napoli 1964, 33, 611. [Google Scholar]
- Sutcliffe, D.W. Salinity fluctuations and the fauna in a salt marsh with special reference to aquatic insects. Trans. Nat. Hist. Soc. Northumb. Durham 1961, 14, 37–56. [Google Scholar]
- Hädicke, C.W.; Rédei, D.; Kment, P. The diversity of feeding habits recorded for water boatmen (Heteroptera: Corixoidea) world-wide with implications for evaluating information on the diet of aquatic insects. Eur. J. Entomol. 2017, 114, 147–159. [Google Scholar] [CrossRef]
- Radley, J.D. Distribution and palaeoenvironmental significance of molluscs in the late Jurassic-early Cretaceous Purbeck Formation of Dorset, southern England. In Life and Environments in Purbeck Times; Milner, A.R., Batten, D.J., Eds.; Palaeontological Association: London, UK, 2002; pp. 41–52. [Google Scholar]
- Shadrin, N.V.; Anufriieva, E.V.; Belyakov, V.P.; Bazhora, A.I. Chironomidae larvae in hypersaline waters of the Crimea: Diversity, distribution, abundance and production. Europ. Zool. J. 2017, 84, 61–72. [Google Scholar] [CrossRef]
- Edwards, H.M. Note sur deux Crustacés fossils de l’Order des Isopodes. Ann. Sci. Nat. Zool. 1843, 20, 326–329. [Google Scholar]
- Morter, A.A. Purbeck-Wealden Beds Mollusca and their relationship to ostracods biostratigraphy, stratigraphical correlation and palaeoecology in the Weald and adjacent areas. Proc. Geol. Assoc. 1984, 95, 217–234. [Google Scholar] [CrossRef]
- Ponomarenko, A.G.; Aristov, D.S.; Bashkuev, A.S.; Gubin, Y.M.; Khramov, A.V.; Lukashevich, E.D.; Popov, Y.A.; Pritykina, L.N.; Sinitsa, S.M.; Sinitshenkova, N.D.; et al. Upper Jurassic Lagerstätte Shar Teg, southwestern Mongolia. Paleontol. J. 2014, 48, 1573–1682. [Google Scholar] [CrossRef]
- Sinitshenkova, N.D. Ecological history of the aquatic insects. In History of Insects, 1st ed.; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 388–417. [Google Scholar]
- Alcocer, J.; Hammer, U.T. Saline lake ecosystems of Mexico. Aquat. Ecosyst. Health Manag. 1998, 1, 291–315. [Google Scholar] [CrossRef]
- Belmar, O.; Velasco, J.; Gutiérrez-Cánovas, C.; Mellado-Díaz, A.; Millán, A.; Wood, P.J. The influence of natural flow regimes on macroinvertebrate assemblages in a semiarid Mediterranean basin. Ecohydrology 2013, 6, 363–379. [Google Scholar] [CrossRef]
- Radley, J.D.; Allen, P. The Wealden (non-marine Lower Cretaceous) of the Weald Sub-basin, southern England. Proc. Geol. Assoc. 2012, 123, 245–318. [Google Scholar] [CrossRef]
- Feist, M. Charophytes. In English Wealden Fossils; Batten, D.J., Ed.; Palaeontological Association: London, UK, 2011; pp. 581–595. [Google Scholar]
- Masese, F.O.; Raburu, P.O. Improving the performance of the EPT Index to accommodate multiple stressors in Afrotropical streams. Afr. J. Aquat. Sci. 2017, 42, 219–233. [Google Scholar] [CrossRef]
- Kladarić, L.; Ćuk, R.; Dukić, I.; Popijač, A.; Ruždjak, A.M. Can Ephemeroptera, Plecoptera, Trichoptera (EPT) assemblages reflect nitrogen and phosphorus load in the riverine ecosystem? Nat. Croat. 2021, 30, 217–230. [Google Scholar] [CrossRef]
- Gouissi, F.M.; Samon, O.; Abahi, K.S.; Adje, D.D.; Tchaou, C.M.; Piami, Z.O.; Okoya, J.G.A.; Gnohossou, M.P. Relationship between macroinvertebrates and physico-chemical parameters to access water quality of the Affon River in Bénin. Adv. Entom. 2019, 7, 92–104. [Google Scholar] [CrossRef][Green Version]
- Hasiotis, S.T.; Kirkland, J.I.; Windscheffel, G.W.; Safris, C. Fossil caddisfly cases (Insecta: Trichoptera), Upper Jurassic Morrison Formation, fruita paleontological area, Colorado. Mod. Geol. 1998, 22, 493–502. [Google Scholar]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coram, R.A.; Jarzembowski, E.A. Immature Insect Assemblages from the Early Cretaceous (Purbeck/Wealden) of Southern England. Insects 2021, 12, 942. https://doi.org/10.3390/insects12100942
Coram RA, Jarzembowski EA. Immature Insect Assemblages from the Early Cretaceous (Purbeck/Wealden) of Southern England. Insects. 2021; 12(10):942. https://doi.org/10.3390/insects12100942
Chicago/Turabian StyleCoram, Robert A., and Edmund A. Jarzembowski. 2021. "Immature Insect Assemblages from the Early Cretaceous (Purbeck/Wealden) of Southern England" Insects 12, no. 10: 942. https://doi.org/10.3390/insects12100942
APA StyleCoram, R. A., & Jarzembowski, E. A. (2021). Immature Insect Assemblages from the Early Cretaceous (Purbeck/Wealden) of Southern England. Insects, 12(10), 942. https://doi.org/10.3390/insects12100942