Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Genomic DNA Extraction
2.2. Mitogenome Sequencing, Assembly and Annotation
2.3. Sequence Analyses
2.4. Sequence Alignments and Phylogenetic Analyses
3. Results and Discussion
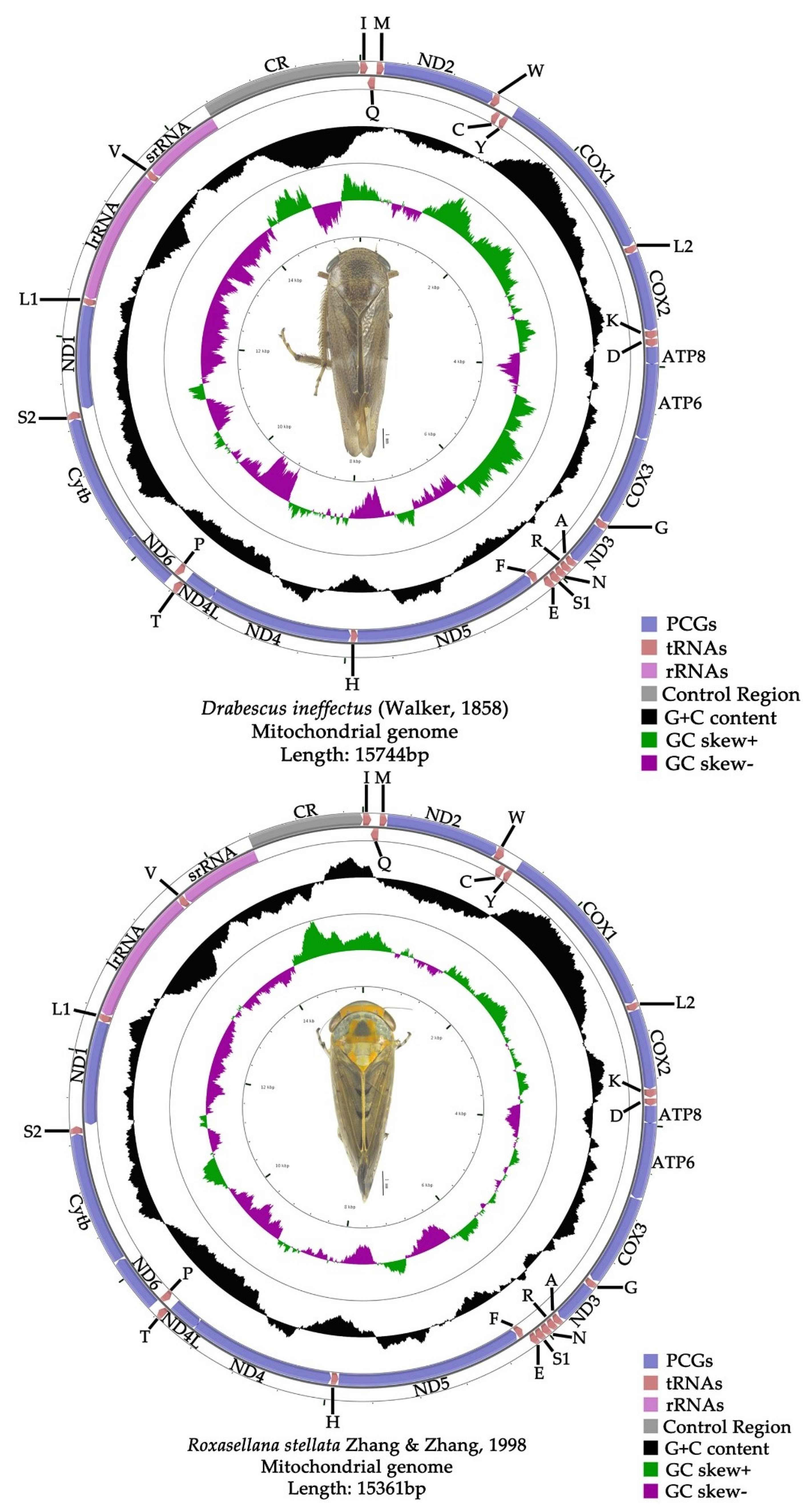
3.1. Mitogenome Organization and Nucleotide Composition
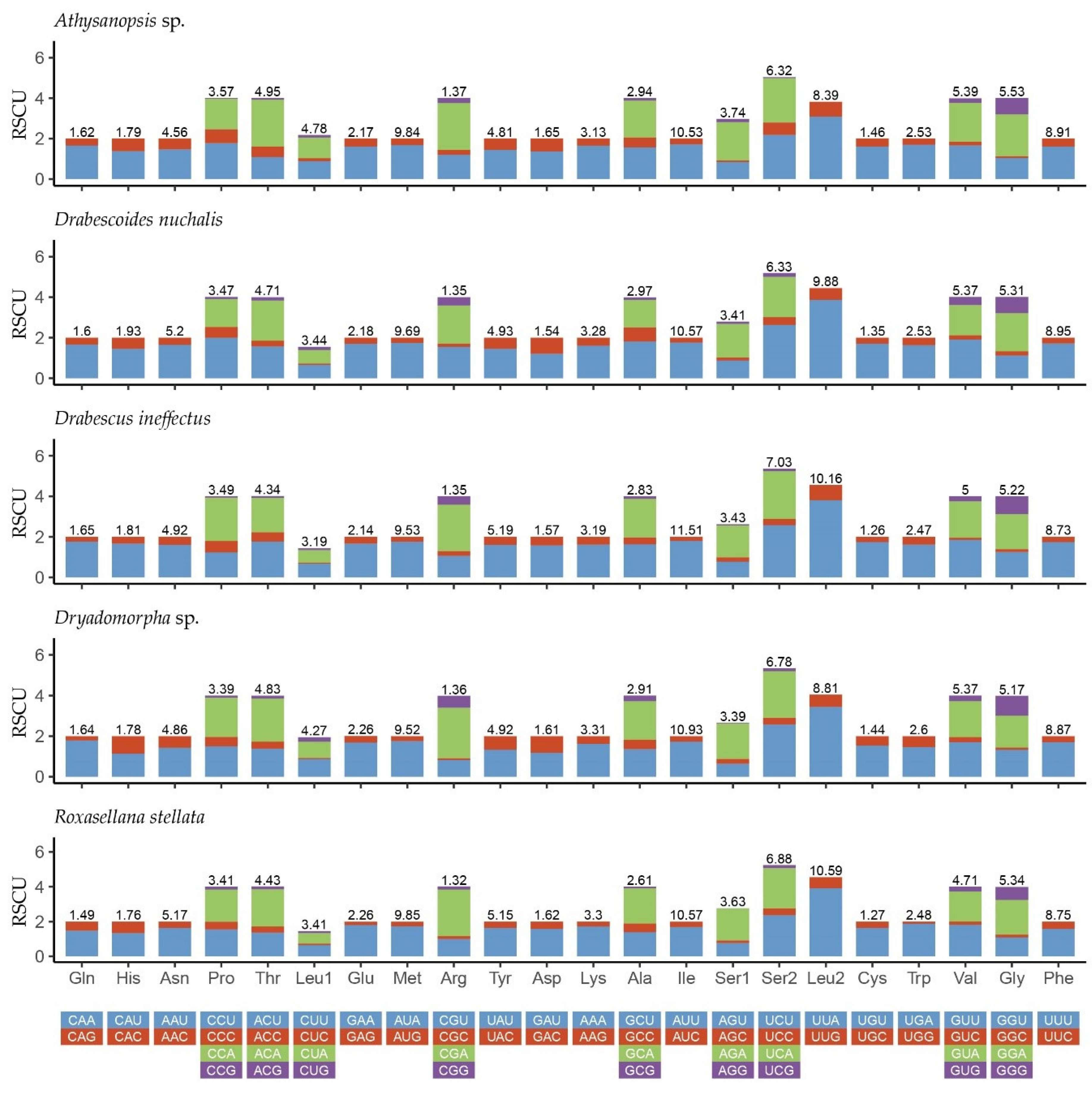
3.2. Protein-Coding Genes and Codon Usage
3.3. Gene Overlaps and Intergenic Spacers
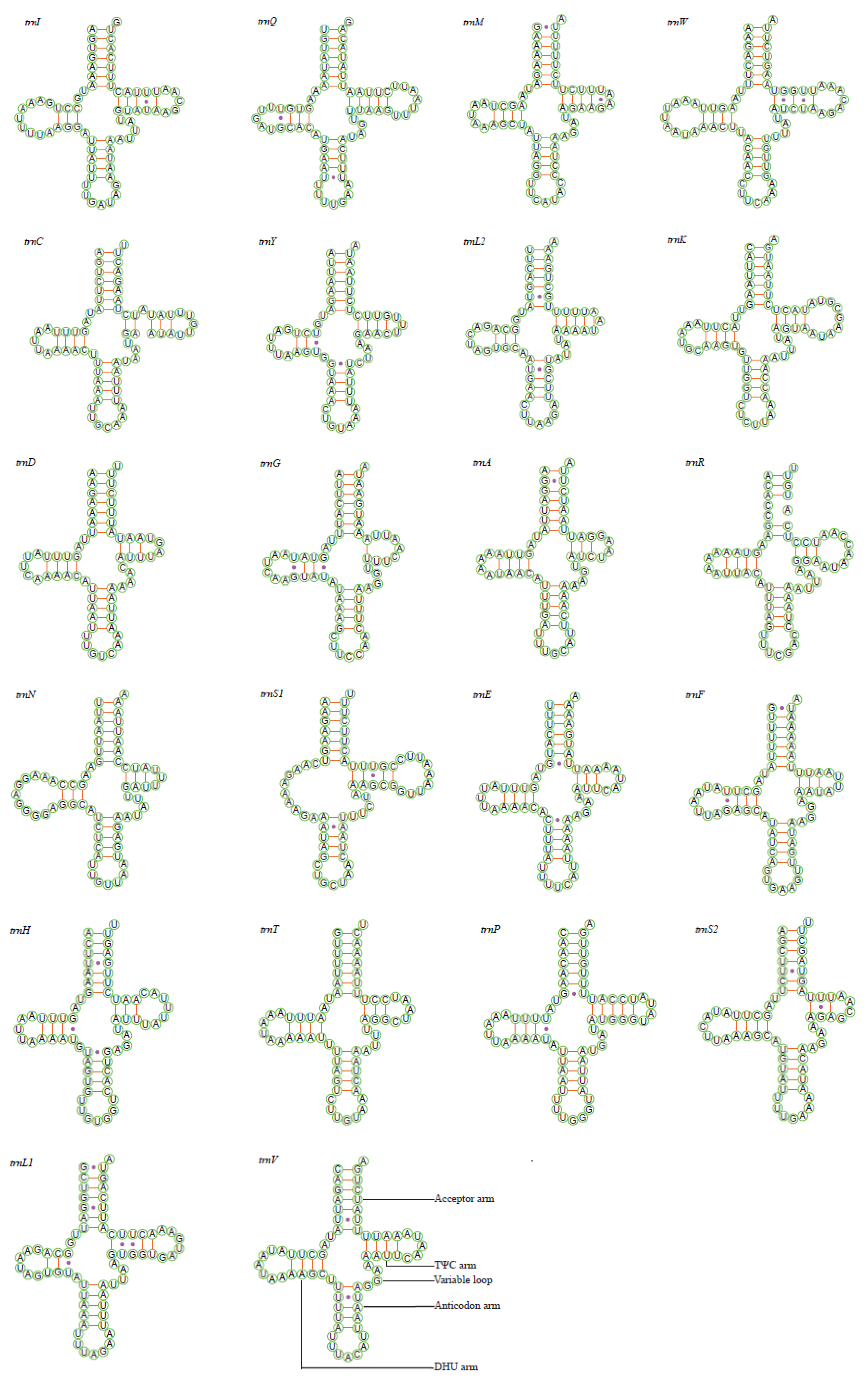
3.4. Transfer and Ribosomal RNA Genes
3.5. A + T-Control Region
3.6. Nucleotide Diversity and Evolutionary Rate Analysis
3.7. Phylogenetic Relationships
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zahniser, J.N.; Dietrich, C.H. A review of the tribes of Deltocephalinae (Hemiptera: Auchenorrhyncha: Cicadellidae). Eur. J. Taxon. 2013, 45, 1–121. [Google Scholar] [CrossRef]
- Linnavuori, R.E.; Al-Ne’amy, K.T. Revision of the African Cicadellidae (Subfamily Selenocephalinae) (Homoptera, Auchenorrhyncha). Acta Zool. Fenn. 1983, 168, 1–103. [Google Scholar]
- Zhang, Y.L.; Webb, M.D. A revised classification of the Asian and Pacific Selenocephaline leafhoppers (Homoptera: Cicadellidae). Bull. Nat. Hist. Mus. Entomol. Series 1996, 65, 1–103. [Google Scholar]
- Xu, D.L.; Zhang, Y.L. Review of Indonesian species of the leafhopper genus Drabescus Stål (Hemiptera: Cicadellidae: Deltocephalinae) with description of two new species. Zootaxa 2018, 4524, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.L.; Zhang, Y.L. A new species of the Oriental leafhopper genus Kutara Distant (Hemiptera: Cicadellidae: Deltocephalinae) from Vietnam. Zootaxa 2020, 4772, 585–592. [Google Scholar] [CrossRef]
- Lu, L.; Webb, M.D.; Zhang, Y.L. A new species of the leafhopper genus Drabescus Stål (Hemiptera: Cicadellidae: Deltocephalinae) from China, with a checklist and key to species. Zootaxa 2017, 4612, 237–246. [Google Scholar] [CrossRef]
- Dmitriev, D.A. Nymphs of some species of the tribes Drabescini and Paraboloponini with a proposed synonymy of Paraboloponini with Drabescini (Hemiptera: Cicadellidae: Deltocephalinae). Orient. Insects 2004, 38, 235–244. [Google Scholar] [CrossRef]
- Zahniser, J.N.; Dietrich, C.H. Phylogeny of the leafhopper subfamily Deltocephalinae (Insecta: Auchenorrhyncha: Cicadellidae) and related subfamilies based on morphology. Syst. Biodivers. 2008, 6, 1–24. [Google Scholar] [CrossRef]
- Zahniser, J.N.; Dietrich, C.H. Phylogeny of the leafhopper subfamily Deltocephalinae (Hemiptera: Cicadellidae) based on molecular and morphological data with a revised family-group classification. Syst. Entomol. 2010, 35, 489–511. [Google Scholar] [CrossRef]
- Dietrich, C.H.; Allen, J.M.; Lemmon, A.R.; Lemmon, E.M.; Takiya, D.M.; Evangelista, O.; Walden, K.K.; Grady, P.G.; Johnson, K.P. Anchored hybrid enrichment-based phylogenomics of leafhoppers and treehoppers (Hemiptera: Cicadomorpha: Membracoidea). Insect Syst. Evol. 2017, 1, 57–72. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Hewitt, G.M. Insect mitochondrial control region: A review of its structure, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 1997, 25, 99–120. [Google Scholar] [CrossRef]
- Wolstenholme, D.R. Genetic novelties in mitochondrial genomes of multicellular animals. Curr. Opin. Genet. Dev. 1992, 2, 918–925. [Google Scholar] [CrossRef]
- Wolstenholme, D.R. Animal mitochondrial DNA: Structure and evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Li, H.; Cai, W.Z. Insufficient power of mitogenomic data in resolving the auchenorrhynchan monophyly. Zool. J. Linn. Sco-Lond. 2017, 183, 776–790. [Google Scholar] [CrossRef]
- Wu, Y.F.; Dai, R.H.; Zhan, H.P.; Qu, L. Complete mitochondrial genome of Drabescoides nuchalis (Hemiptera: Cicadellidae). Mitochondrial DNA Part A 2015, 27, 3626–3627. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Leavengood, J.M., Jr.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.G.; Cai, W.Z. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef]
- Su, T.J.; He, B.; Li, K.; Liang, A.P. Comparative analysis of the mitochondrial genomes of oriental spittlebug tribe Cosmoscartini: Insights into the relationships among closely related taxa. BMC Genomics 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Mao, M.; Yang, X.S.; Bennett, G. The complete mitochondrial genome of Entylia carinata (Hemiptera: Membracidae). Mitochondrial DNA Part B 2016, 1, 662–663. [Google Scholar] [CrossRef]
- Hu, K.; Yuan, F.; Dietrich, C.H.; Yuan, X.Q. Structural features and phylogenetic implications of four new mitogenomes of Centrotinae (Hemiptera: Membracidae). Int. J. Biol. Macromol. 2019, 139, 1018–1027. [Google Scholar] [CrossRef]
- Wang, J.J.; Xing, J.C. Complete mitochondrial genome of Abrus expansivus (Hemiptera: Cicadellidae: Deltocephalinae) from China. Mitochondrial DNA Part B 2019, 4, 197–198. [Google Scholar] [CrossRef]
- Song, N.; Cai, W.Z.; Li, H. Deep-level phylogeny of Cicadomorpha inferred from mitochondrial genomes sequenced by NGS. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Yu, P.F.; Wang, M.X.; Cui, L.; Chen, X.X.; Han, B.Y. The complete mitochondrial genome of Tambocerus sp. (Hemiptera: Cicadellidae). Mitochondrial DNA Part A 2017, 28, 133–134. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.M.; Zhang, C.N.; Dietrich, C.H.; Zhang, Y.L.; Dai, W. Characterization of the complete mitochondrial genomes of Maiestas dorsalis and Japananus hyalinus (Hemiptera: Cicadellidae) and comparison with other Membracoidea. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Yang, X.S.; Bennett, G. The complete mitochondrial genome of Macrosteles quadrilineatus (Hemiptera: Cicadellidae). Mitochondrial DNA Part B 2017, 2, 173–175. [Google Scholar] [CrossRef]
- Du, Y.M.; Dietrich, C.H.; Dai, W. Complete mitochondrial genome of Macrosteles quadrimaculatus (Matsumura) (Hemiptera: Cicadellidae: Deltocephalinae) with a shared tRNA rearrangement and its phylogenetic implications. Int. J. Biol. Macromol. 2018, 122, 1027–1034. [Google Scholar] [CrossRef]
- Xing, J.C.; Wang, J.J. Complete mitochondrial genome of Paralaevicephalus gracilipenis (Hemiptera: Cicadellidae: Deltocephalinae) from China. Mitochondrial DNA Part B 2019, 4, 1372–1373. [Google Scholar] [CrossRef]
- Du, Y.M.; Dai, W.; Dietrich, C.H. Mitochondrial genomic variation and phylogenetic relationships of three groups in the genus Scaphoideus (Hemiptera: Cicadellidae: Deltocephalinae). Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef][Green Version]
- Zhong, L.K.; Li, H.X.; Yu, X.F.; Yang, M.F. Complete mitochondrial genome sequence of Cicadella viridis (Hemiptera: Cicadellidae: Cicadellinae). Mitochondrial DNA Part B 2019, 4, 1287–1288. [Google Scholar] [CrossRef]
- Wang, J.J.; Dai, R.H. Complete mitochondrial genome of Taharana fasciana (Insecta, Hemiptera: Cicadellidae) and comparison with other Cicadellidae insects. Genetica 2017, 145, 593–602. [Google Scholar] [CrossRef]
- Dai, R.H.; Wang, J.J.; Yang, M.F. The complete mitochondrial genome of the leafhopper Idioscopus clypealis (Hemiptera: Cicadellidae: Idiocerinae). Mitochondrial DNA Part B 2018, 3, 32–33. [Google Scholar] [CrossRef]
- Wang, J.J.; Yang, M.F.; Dai, R.H.; Li, H.; Wang, X.Y. Characterization and phylogenetic implications of the complete mitochondrial genome of Idiocerinae (Hemiptera: Cicadellidae). Int. J. Biol. Macromol. 2018, 120, 2366–2372. [Google Scholar] [CrossRef]
- Wang, J.J.; Yang, M.F.; Dai, R.H.; Li, H. Complete mitochondrial genome of Evacanthus heimianus (Hemiptera: Cicadellidae: Evacanthinae) from China. Mitochondrial DNA Part B 2019, 4, 284–285. [Google Scholar] [CrossRef]
- Wang, J.J.; Wu, Y.F.; Dai, R.H.; Yang, M.F. Comparative mitogenomes of six species in the subfamily Iassinae (Hemiptera: Cicadellidae) and phylogenetic analysis. Int. J. Biol. Macromol. 2020, 149, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Li, D.F.; Li, H.; Yang, M.F.; Dai, R.H. Structural and phylogenetic implications of the complete mitochondrial genome of Ledra auditura. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Dai, R.H.; Li, H.; Zhan, H.P. Characterization of the complete mitochondrial genome of Japanagallia spinosa and Durgades nigropicta (Hemiptera: Cicadellidae: Megophthalminae). Biochem. Syst. Ecol. 2017, 74, 33–41. [Google Scholar] [CrossRef]
- Shi, R.; Yu, X.F.; Yang, M.F. Complete mitochondrial genome of Ghauriana sinensis (Hemiptera: Cicadellidae: Typhlocybinae). Mitochondrial DNA Part B 2020, 5, 1367–1368. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Haeseler, A.V.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Garey, J.R.; Wolstenholme, D.R. Platyhelminth mitochondrial DNA: Evidence for early evolutionary origin of a tRNAserAGN that contains a dihydrouridine arm replacement loop, and of serine-specifying AGA and AGG codons. J. Mol. Evol. 1989, 28, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.Z.; Yan, H.B.; Guo, A.J.; Zhu, X.Q.; Wang, Y.C.; Shi, W.G.; Chen, H.T.; Zhan, F.; Zhang, S.H.; Fu, B.Q.; et al. Complete mitochondrial genomes of Taenia multiceps, T. hydatigena and T. pisiformis: Additional molecular markers for a tapeworm genus of human and animal health significance. BMC Genomics 2010, 11, 447. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superfamily | Family | Subfamily | Species | Accession Number | Reference |
|---|---|---|---|---|---|
| Cicadoidea | Cicadidae | Cicadinae | Cryptotympana atrata | JQ910980 | [25] |
| Cercopoidea | Cercopidae | Cercopinae | Cosmoscarta dorsimacula | NC_040115 | [26] |
| Membracoidea | Membracidae | Smiliinae | Entylia carinata | NC_033539 | [27] |
| Centrotinae | Centrotus cornutus | KX437728 | [16] | ||
| Leptocentrus albolineatus | MK746137 | [28] | |||
| Tricentrus brunneus | NC_044708 | [28] | |||
| Cicadellidae | Deltocephalinae | Abrus expansivus | NC_045238 | [29] | |
| Norvellina sp. | KY039131 | [30] | |||
| Paramacrosteles nigromaculatus | NC_045270 | Direct Submission | |||
| Tambocerus sp. | KT827824 | [31] | |||
| Exitianus sp. | KX437722 | [16] | |||
| Exitianus indicus | KY039128 | [30] | |||
| Nephotettix cincticeps | NC_026977 | Direct Submission | |||
| Cicadula sp. | KX437724 | [16] | |||
| Alobaldia tobae | KY039116 | [30] | |||
| Maiestas dorsalis | NC_036296 | [32] | |||
| Athysanopsis sp. | KX437726 | [16] | |||
| Drabescoides nuchalis | NC_028154 | [17] | |||
| Drabescus ineffectus | MT527188 | This study | |||
| Roxasellana stellata | MT527187 | This study | |||
| Dryadomorpha sp. | KX437736 | [16] | |||
| Macrosteles quadrilineatus | KY645960 | [33] | |||
| Macrosteles quadrimaculatus | NC_039560 | [34] | |||
| Hishimonus phycitis | KX437727 | [16] | |||
| Hishimonoides recurvatis | KY364883 | Unpublished | |||
| Japananus hyalinus | NC_036298 | [32] | |||
| Orosius orientalis | KY039146 | [30] | |||
| Paralaevicephalus gracilipenis | MK450366 | [35] | |||
| Psammotettix sp.1 | KX437725 | [16] | |||
| Psammotettix sp.2 | KX437742 | [16] | |||
| Yanocephalus yanonis | NC_036131 | [30] | |||
| Phlogotettix sp.1 | KY039135 | [30] | |||
| Phlogotettix sp.2 | KX437721 | [16] | |||
| Scaphoideus maai | KY817243 | [36] | |||
| Scaphoideus nigrivalveus | KY817244 | [36] | |||
| Scaphoideus varius | KY817245 | [36] | |||
| Cicadellinae | Cicadella viridis | MK335936 | [37] | ||
| Homalodisca vitripennis | NC_006899 | Direct Submission | |||
| Coelidiinae | Olidiana ritcheriina | NC_045207 | Direct Submission | ||
| Taharana fasciana | KY886913 | [38] | |||
| Eurymelinae | Idioscopus clypealis | NC_039642 | [39] | ||
| Populicerus populi | MH492318 | [40] | |||
| Evacanthinae | Evacanthus heimianus | MG813486 | [41] | ||
| Sophonia linealis | KX437723 | [16] | |||
| Iassinae | Batracomorphus lateprocessus | MG813489 | [42] | ||
| Krisna rufimarginata | NC_046068 | [42] | |||
| Ledrinae | Ledra auditura | MK387845 | [43] | ||
| Tituria pyramidata | NC_046701 | Direct Submission | |||
| Megophthalminae | Durgades nigropicta | KY123686 | [44] | ||
| Japanagallia spinosa | NC_035685 | [44] | |||
| Typhlocybinae | Mitjaevia protuberanta | NC_047465 | Unpublished | ||
| Ghauriana sinensis | MN699874 | [45] | |||
| Limassolla lingchuanensis | NC_046037 | Unpublished |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Yu, T.; Zhang, Y. Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications. Insects 2020, 11, 534. https://doi.org/10.3390/insects11080534
Xu D, Yu T, Zhang Y. Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications. Insects. 2020; 11(8):534. https://doi.org/10.3390/insects11080534
Chicago/Turabian StyleXu, Deliang, Tinghao Yu, and Yalin Zhang. 2020. "Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications" Insects 11, no. 8: 534. https://doi.org/10.3390/insects11080534
APA StyleXu, D., Yu, T., & Zhang, Y. (2020). Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications. Insects, 11(8), 534. https://doi.org/10.3390/insects11080534