Development on Infected Citrus over Generations Increases Vector Infection by ‘Candidatus Liberibacter Asiaticus in Diaphorina citri’

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
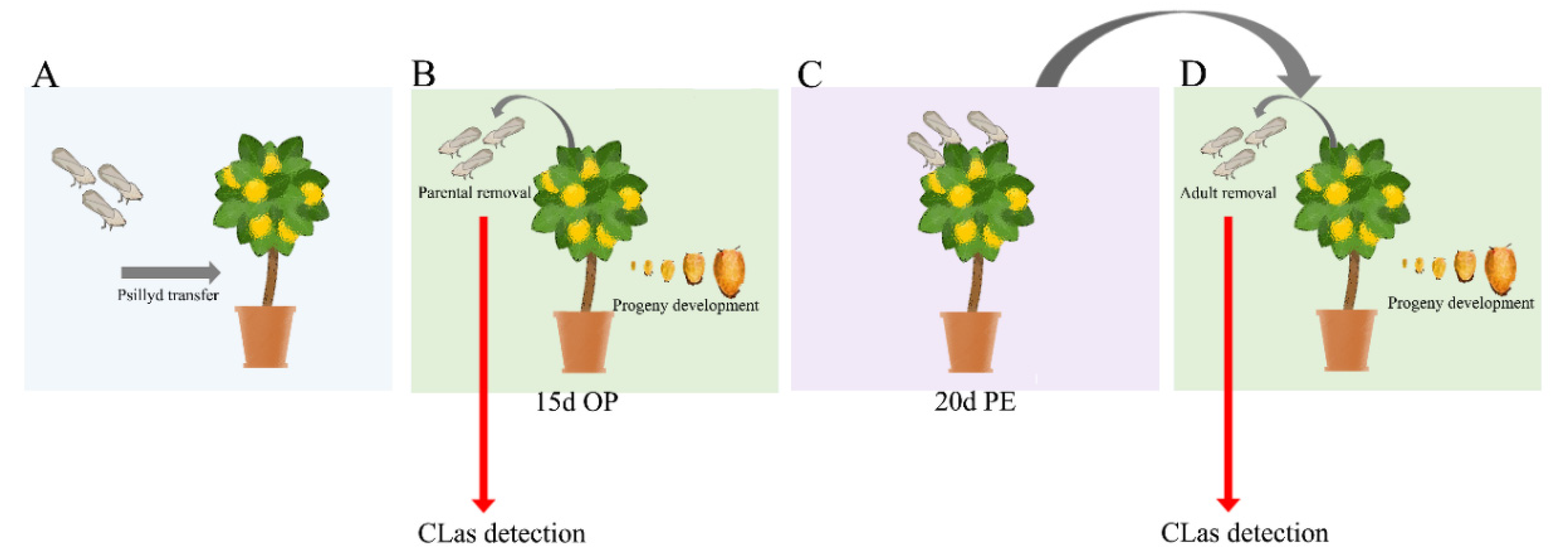
2.2. CLas Acquisition Bioassay over Multiple ACP Generations
2.3. Nucleic Acid Extraction and CLas Detection by qPCR in Psyllid Samples
2.4. Molecular Standard Curve
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zheng, Z.; Chen, J.C.; Deng, X. Historical Perspectives, Management, and Current Research of Citrus HLB in Guangdong Province of China, Where the Disease has been Endemic for Over a Hundred Years. Phytopathology 2018, 108, 1224–1236. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Hall, D.G.; Hosseinzadeh, S.; Heck, M.L. The quest for a non-vector psyllid: Natural variation in acquisition and transmission of the huanglongbing pathogen “Candidatus Liberibacter asiaticus” by Asian citrus psyllid isofemale lines. PLoS ONE 2018, 13, e0195804. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.-D.; Shatters, R.G.; Hall, D. Localization of Candidatus Liberibacter asiaticus, Associated with Citrus Huanglongbing Disease, in its Psyllid Vector using Fluorescence in situ Hybridization. J. Phytopathol. 2011, 159, 726–734. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Shatters, R.G.; Lynch, C.; Hall, D.G. Detection and Relative Titer of Candidatus Liberibacter asiaticus in the Salivary Glands and Alimentary Canal of Diaphorina citri (Hemiptera: Psyllidae) Vector of Citrus Huanglongbing Disease. Ann. Entomol. Soc. Am. 2011, 104, 526–533. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Ramos, J.E.; Hall, D.G.; Dawson, W.; Shatters, R.G. Acquisition, Replication and Inoculation of Candidatus Liberibacter asiaticus following Various Acquisition Periods on Huanglongbing-Infected Citrus by Nymphs and Adults of the Asian Citrus Psyllid. PLoS ONE 2016, 11, e0159594. [Google Scholar] [CrossRef] [PubMed]
- Canale, M.C.; Tomaseto, A.F.; Haddad, M.D.L.; Lopes, J.R.S.; Filho, H.D.C. Latency and Persistence of “Candidatus Liberibacter asiaticus” in its Psyllid Vector, Diaphorina citri Kuwayama (Hemiptera: Liviidae). Phytopathology 2017, 107, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Levy, L.; Hartung, J. Quantitative Distribution of “Candidatus Liberibacter asiaticus” in Citrus Plants with Citrus Huanglongbing. Phytopathology 2009, 99, 139–144. [Google Scholar] [CrossRef]
- Gottwald, T.R. Current Epidemiological Understanding of Citrus Huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef]
- Lee, J.A.; Halbert, S.E.; Dawson, W.O.; Robertson, C.J.; Keesling, J.E.; Singer, B.H. Asymptomatic spread of huanglongbing and implications for disease control. Proc. Natl. Acad. Sci. USA 2015, 112, 7605–7610. [Google Scholar] [CrossRef]
- Canale, M.C.; Komada, K.M.A.; Lopes, J.R.S. Latency and incubation of “Candidatus Liberibacter asiaticus” in citrus after vector inoculation. Trop. Plant Pathol. 2019, 1–7. [Google Scholar] [CrossRef]
- Shapiro, L.R.; Seidl-Adams, I.; De Moraes, C.M.; Stephenson, A.G.; Mescher, M.C. Dynamics of short- and long-term association between a bacterial plant pathogen and its arthropod vector. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rashed, A.; Nash, T.D.; Paetzold, L.; Workneh, F.; Rush, C.M. Transmission Efficiency of “Candidatus Liberibacter solanacearum” and Potato Zebra Chip Disease Progress in Relation to Pathogen Titer, Vector Numbers, and Feeding Sites. Phytopathology 2012, 102, 1079–1085. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sisterson, M.S. Effects of Insect-Vector Preference for Healthy or Infected Plants on Pathogen Spread: Insights from a Model. J. Econ. Entomol. 2008, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.G.; Moulton, K.M. Transmission Rates of “Candidatus Liberibacter asiaticus” to Greenhouse Seedlings by Laboratory Colonies of Asian Citrus Psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2018, 111, 2546–2552. [Google Scholar] [CrossRef]
- Coletta-Filho, H.D.; Daugherty, M.P.; Ferreira, C.; Lopes, J.R.S. Temporal Progression of “Candidatus Liberibacter asiaticus” Infection in Citrus and Acquisition Efficiency by Diaphorina citri. Phytopathology 2014, 104, 416–421. [Google Scholar] [CrossRef][Green Version]
- Swallow, W.H. Relative Mean Squared Error and Cost Considerations in Choosing Group Size for Group Testing to Estimate Infection Rates and Probabilities of Disease Transmission. Phytopathology 1987, 77, 1376. [Google Scholar] [CrossRef]
- Swift, M.L. GraphPad Prism, Data Analysis, and Scientific Graphing. J. Chem. Inf. Comput. Sci. 1997, 37, 411–412. [Google Scholar] [CrossRef]
- Inoue, H.; Ohnishi, J.; Ito, T.; Tomimura, K.; Miyata, S.; Iwanami, T.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2009, 155, 29–36. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission Parameters for Candidatus Liberibacter asiaticus by Asian Citrus Psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef]
- Fisher, T.W.; Vyas, M.; He, R.; Nelson, W.; Cicero, J.M.; Willer, M.; Kim, R.; Kramer, R.; May, G.A.; Crow, J.A.; et al. Comparison of Potato and Asian Citrus Psyllid Adult and Nymph Transcriptomes Identified Vector Transcripts with Potential Involvement in Circulative, Propagative Liberibacter Transmission. Pathogens 2014, 3, 875–907. [Google Scholar] [CrossRef]
- Kruse, A.; Fattah-Hosseini, S.; Saha, S.; Johnson, R.; Warwick, E.; Sturgeon, K.; Mueller, L.; MacCoss, M.J.; Shatters, R.G.; Heck, M.L. Combining ’omics and microscopy to visualize interactions between the Asian citrus psyllid vector and the Huanglongbing pathogen Candidatus Liberibacter asiaticus in the insect gut. PLoS ONE 2017, 12, e0179531. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M.; Fattah-Hosseini, S.; Levy, A.; Cilia, M. Morphological abnormalities and cell death in the Asian citrus psyllid (Diaphorina citri) midgut associated with Candidatus Liberibacter asiaticus. Sci. Rep. 2016, 6, 33418. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Hijaz, F.; Ebert, T.A.; Rogers, M.E. A Plant Bacterial Pathogen Manipulates its Insect Vector’s Energy Metabolism. Appl. Environ. Microbiol. 2016, 83, e03005-16. [Google Scholar] [CrossRef] [PubMed]
- Nehela, Y.; Killiny, N. “Candidatus Liberibacter asiaticus” and its Vector, Diaphorina citri, Augment the Tricarboxylic Acid Cycle of Their Host via the γ-Aminobutyric Acid Shunt and Polyamines Pathway. Mol. Plant Microbe Interact. 2019, 32, 413–427. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Ammar, E.-D.; Hall, D.G.; Shatters, R.G.; Lapointe, S.L. Prolonged phloem ingestion by Diaphorina citri nymphs compared to adults is correlated with increased acquisition of citrus greening pathogen. Sci. Rep. 2018, 8, 10352. [Google Scholar] [CrossRef]
- Mann, M.; Fattah-Hosseini, S.; Ammar, E.-D.; Stange, R.; Warrick, E.; Sturgeon, K.; Shatters, R.G.; Heck, M.L. Diaphorina citri Nymphs Are Resistant to Morphological Changes Induced by “Candidatus Liberibacter asiaticus” in Midgut Epithelial Cells. Infect. Immun. 2018, 86, e00889-17. [Google Scholar] [CrossRef]
- Jain, M.; Munoz, A.; Gabriel, D.W. Concomitant Loss of the Glyoxalase System and Glycolysis Makes the Uncultured Pathogen “Candidatus Liberibacter asiaticus” an Energy Scavenger. Appl. Environ. Microbiol. 2017, 83, e01670-17. [Google Scholar] [CrossRef]
- Lu, Z.; Killiny, N. Huanglongbing pathogen Candidatus Liberibacter asiaticus exploits the energy metabolism and host defence responses of its vector Diaphorina citri. Physiol. Entomol. 2017, 42, 319–335. [Google Scholar] [CrossRef]
- Mann, R.S.; Pelz-Stelinski, K.; Hermann, S.L.; Tiwari, S.; Stelinski, L.L. Sexual Transmission of a Plant Pathogenic Bacterium, Candidatus Liberibacter asiaticus, between Conspecific Insect Vectors during Mating. PLoS ONE 2011, 6, e29197. [Google Scholar] [CrossRef]
- Kelley, A.J.; Pelz-Stelinski, K.S. Maternal Contribution of Candidatus Liberibacter asiaticus to Asian Citrus Psyllid (Hemiptera: Liviidae) Nymphs through Oviposition Site Inoculation and Transovarial Transmission. J. Econ. Entomol. 2019, 112, 2565–2568. [Google Scholar] [CrossRef]
- Wu, F.; Huang, J.; Xu, M.; Fox, E.G.P.; Beattie, G.A.C.; Holford, P.; Cen, Y.; Deng, X. Host and environmental factors influencing “Candidatus Liberibacter asiaticus” acquisition in Diaphorina citri. Pest Manag. Sci. 2018, 74, 2738–2746. [Google Scholar] [CrossRef] [PubMed]
- Sengoda, V.G.; Buchman, J.L.; Henne, D.C.; Pappu, H.R.; Munyaneza, J.E. “Candidatus Liberibacter solanacearum” titer over time in Bactericera cockerelli (Hemiptera: Triozidae) after acquisition from infected potato and tomato plants. J. Econ. Entomol. 2013, 106, 1964–1972. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.-D.; Ramsey, J.S.; Mahney, J.; Hall, D.G.; Cilia, M. Effect of host switch between Murraya and citrus plants on acquisition and transmission of huanglongbing (citrus greening) bacterium by the Asian citrus psyllid Diaphorina citri. In Proceedings of the 6th International Research Conference on HLB, Riverside, CA, USA, 12–15 March 2019; p. 212. [Google Scholar]
- Wu, F.; Qureshi, J.A.; Huang, J.; Fox, E.G.P.; Deng, X.; Wan, F.; Liang, G.; Cen, Y. Host Plant-Mediated Interactions between “Candidatus Liberibacter asiaticus” and its Vector Diaphorina citri Kuwayama (Hemiptera: Liviidae). J. Econ. Entomol. 2018, 111, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, D.M.; Pacheco, I.D.S.; Alves, G.R.; Granato, L.M.; Rashidi, M.; Turner, D.; Levy, A.; Machado, M.A. Friend or foe? Relationship between “Candidatus Liberibacter asiaticus” and Diaphorina citri. Trop. Plant Pathol. 2020, 1–13. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Chavez, J.D.; Johnson, R.; Hosseinzadeh, S.; Mahoney, J.E.; Mohr, J.P.; Robison, F.; Zhong, X.; Hall, D.G.; MacCoss, M.; et al. Protein interaction networks at the host–microbe interface in Diaphorina citri, the insect vector of the citrus greening pathogen. R. Soc. Open Sci. 2017, 4, 160545. [Google Scholar] [CrossRef]
- Login, F.H.; Heddi, A. Insect immune system maintains long-term resident bacteria through a local response. J. Insect Physiol. 2013, 59, 232–239. [Google Scholar] [CrossRef]
- Ratzka, C.; Gross, R.; Feldhaar, H. Endosymbiont Tolerance and Control within Insect Hosts. Insects 2012, 3, 553–572. [Google Scholar] [CrossRef]
- Coy, M.R.; Stelinski, L.L. Great Variability in the Infection Rate of “Candidatus Liberibacter Asiaticus” in Field Populations of Diaphorina citri (Hemiptera: Liviidae) in Florida. Fla. Entomol. 2015, 98, 356–357. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Generations | Total Number of Samples | Number of ACP per Sample | Number of Positive Samples (Frequency) | p (×100) (%) | Number of CLas Cells per 100 ng of DNA |
|---|---|---|---|---|---|
| Parental | 15 | 5 | 6 (0.4) | 9.71 | 4.79 × 100d |
| F1 | 33 | 5 | 25 (0.75) | 24.21 | 1.40 × 101c |
| F2 | 38 | 5 | 33 (0.86) | 32.51 | 6.85 × 101b |
| F4 | 15 | 5 | 15 (1.0) | - | 5.39 × 103a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza Pacheco, I.; Manzano Galdeano, D.; Spotti Lopes, J.R.; Machado, M.A. Development on Infected Citrus over Generations Increases Vector Infection by ‘Candidatus Liberibacter Asiaticus in Diaphorina citri’. Insects 2020, 11, 469. https://doi.org/10.3390/insects11080469
de Souza Pacheco I, Manzano Galdeano D, Spotti Lopes JR, Machado MA. Development on Infected Citrus over Generations Increases Vector Infection by ‘Candidatus Liberibacter Asiaticus in Diaphorina citri’. Insects. 2020; 11(8):469. https://doi.org/10.3390/insects11080469
Chicago/Turabian Stylede Souza Pacheco, Inaiara, Diogo Manzano Galdeano, João Roberto Spotti Lopes, and Marcos Antonio Machado. 2020. "Development on Infected Citrus over Generations Increases Vector Infection by ‘Candidatus Liberibacter Asiaticus in Diaphorina citri’" Insects 11, no. 8: 469. https://doi.org/10.3390/insects11080469
APA Stylede Souza Pacheco, I., Manzano Galdeano, D., Spotti Lopes, J. R., & Machado, M. A. (2020). Development on Infected Citrus over Generations Increases Vector Infection by ‘Candidatus Liberibacter Asiaticus in Diaphorina citri’. Insects, 11(8), 469. https://doi.org/10.3390/insects11080469