Lethal and Sublethal Effects of Methyl Benzoate on the Predatory Bug Nesidiocoris tenuis





Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Reagents
2.2. Lethal Effects of MB on N. tenuis
2.3. Sublethal Effects of MB on N. tenuis
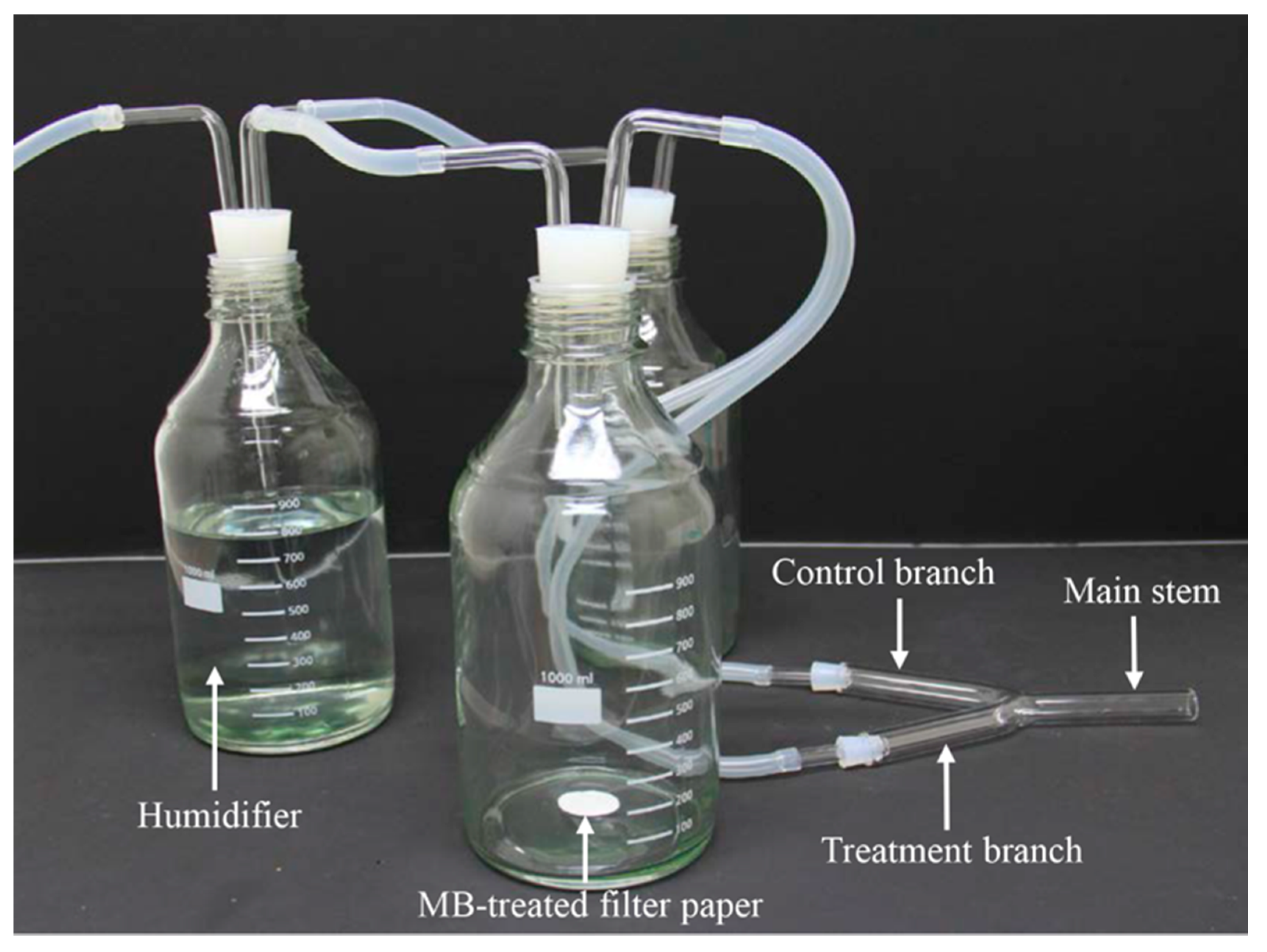
2.4. Y-tube Olfactometer Behavioral Assay
2.5. Statistical Analysis
3. Results
3.1. Lethal Effects of MB on N. tenuis
3.2. Sublethal Effects of MB on N. tenuis
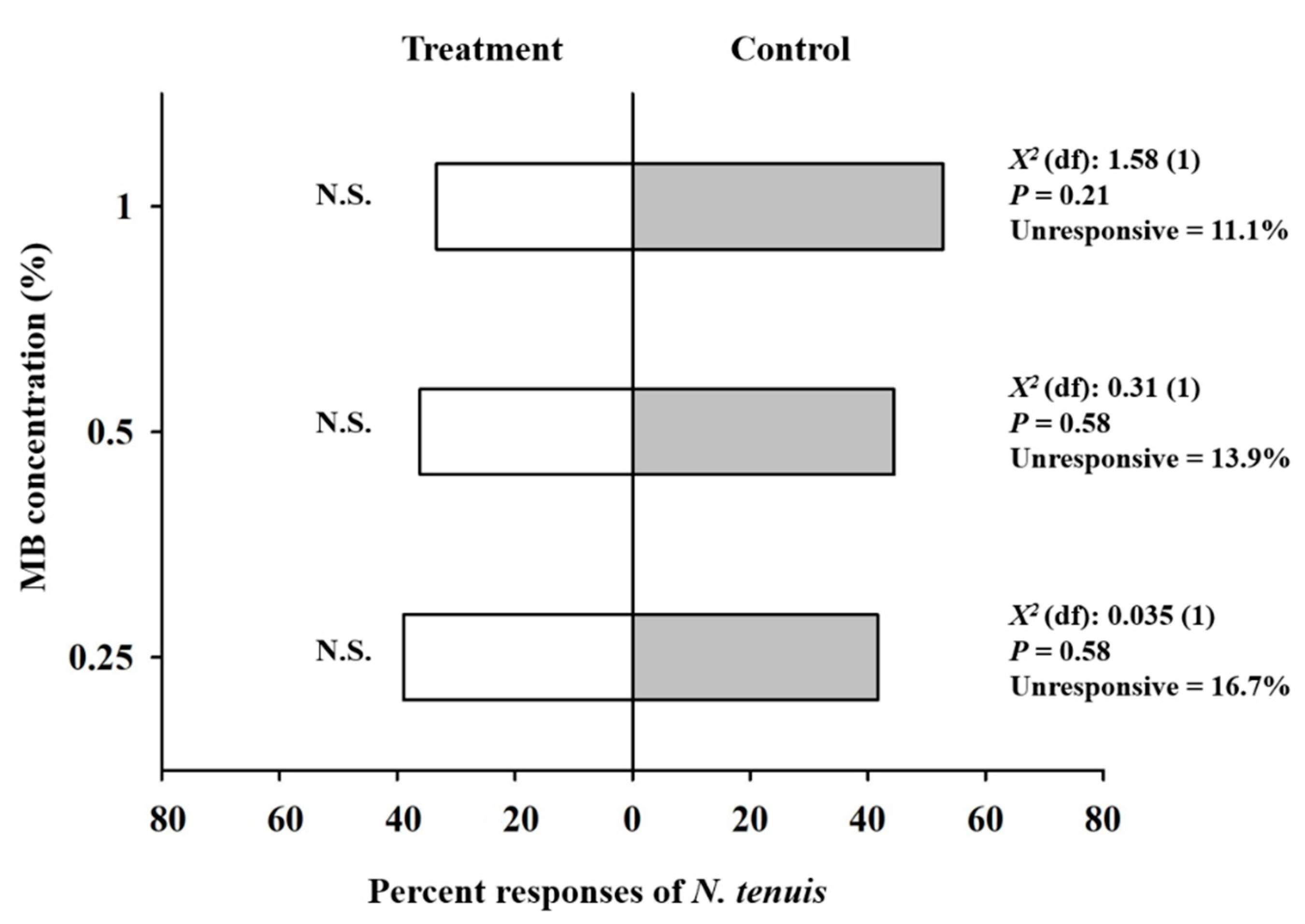
3.3. Olfactory Responses of N. tenuis to MB
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Roditakis, E.; Fytrou, N.; Staurakaki, M.; Vontas, J.; Tsagkarakou, A. Activity of flonicamid on the sweet potato whitely Bemisia tabaci (Homoptera: Aleyrodidae) and its natural enemies. Pest Manag. Sci. 2014, 70, 1460–1467. [Google Scholar] [CrossRef]
- Zappalà, L.; Siscaro, G.; Biondi, A.; Mollá, O.; González-Cabrera, J.; Urbaneja, A. Efficacy of sulphur on Tuta absoluta and its side effects on the predator Nesidiocoris tenuis. J. Appl. Entomol. 2012, 136, 401–409. [Google Scholar] [CrossRef]
- Martinou, A.F.; Seraphides, N.; Stavrinides, M.C. Lethal and behavioral effects of pesticides on the insect predator Macrolophus pygmaeus. Chemosphere 2014, 96, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Mills, N.J.; Beers, E.H.; Shearer, P.W.; Unruh, T.R.; Amarasekare, K.G. Comparative analysis of pesticide effects on natural enemies in western orchards: A synthesis of laboratory bioassay data. Biol. Control 2016, 102, 17–25. [Google Scholar] [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef]
- Candolfi, M.P.; Barrett, P.J.; Campbell, R.; Foster, N.; Grandy, M.-C.; Lewis, P.A.; Oomen, R.; Schmuck, H.V. Guidance document on regulatory testing and risk assessment procedures for plant protection products with non-target artrhropods. In Proceedings of the ESCORT 2 Workshop (European Standard Characteristics of Non-Target Arthropod Regulatory Testing), Wageningen, The Netherlands, 21–23 March 2000. [Google Scholar]
- Bengochea, P.; Budia, F.; Viñuela, E.; Medina, P. Are kaolin and copper treatments safe to the olive fruit fly parasitoid Psyttalia concolor? J. Pest Sci. 2014, 87, 351–359. [Google Scholar] [CrossRef]
- Biondi, A.; Zappalà, L.; Stark, J.D.; Desneux, N. Do Biopesticides affect the demographic traits of a parasitoid wasp and its biocontrol services through sublethal effects? PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Wanumen, A.C.; Carvalho, G.A.; Medina, P.; Vinuela, E.; Adan, A. Residual acute toxicity of some modern insecticides toward two mirid predators of tomato pests. J. Econ. Entomol. 2016, 109, 1079–1085. [Google Scholar] [CrossRef]
- Planes, L.; Catalán, J.; Tena, A.; Porcuna, J.L.; Jacas, J.A.; Izquierdo, J.; Urbaneja, A. Lethal and sublethal effects of spirotetramat on the mealybug destroyer, Cryptolaemus montrouzieri. J. Pest Sci. 2013, 86, 321–327. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Lacasa, A.; Arnó, J.; Castañé, C.; Alomar, O. Life history parameters for Nesidiocoris tenuis (Reuter) (Het., Miridae) under different temperature regimes. J. Appl. Entomol. 2009, 133, 125–132. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Release rate for a pre-plant application of Nesidiocoris tenuis for Bemisia tabaci control in tomato. BioControl 2012, 57, 809–817. [Google Scholar] [CrossRef]
- Perdikis, D.; Arvaniti, K. Nymphal development on plant vs. leaf with and without prey for two omnivorous predators: Nesidiocoris tenuis (Reuter, 1895) (Hemiptera: Miridae) and Dicyphus errans (Wolff, 1804) (Hemiptera: Miridae). Entomol. Gen. 2016, 297–306. [Google Scholar] [CrossRef]
- Campos, M.R.; Biondi, A.; Adiga, A.; Guedes, R.N.C.; Desneux, N. From the western palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. J. Pest Sci. 2017, 90, 787–796. [Google Scholar] [CrossRef]
- Zappalà, L.; Biondi, A.; Alma, A.; Al-Jboory, I.J.; Arnò, J.; Bayram, A.; Chailleux, A.; El-Arnaouty, A.; Gerling, D.; Guenaoui, Y.; et al. Natural enemies of the South American moth, Tuta absoluta, in Europe, North Africa and Middle East, and their potential use in pest control strategies. J. Pest Sci. 2013, 86, 635–647. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.-H.; Desneux, N. Ecology, Worldwide Spread, and Management of the invasive south american tomato pinworm, Tuta absoluta: Past, present, and future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Alomar, O.; Riudavets, J.; Castañe, C. Macrolophus caliginosus in the biological control of Bemisia tabaci on greenhouse melons. Biol. Control 2006, 36, 154–162. [Google Scholar] [CrossRef]
- Mollá, O.; Biondi, A.; Alonso-Valiente, M.; Urbaneja, A. A comparative life history study of two mirid bugs preying on Tuta absoluta and Ephestia kuehniella eggs on tomato crops: Implications for biological control. BioControl 2014, 59, 175–183. [Google Scholar] [CrossRef]
- Calvo, J.; Bolckmans, K.; Stansly, P.A.; Urbaneja, A. Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato. BioControl 2009, 54, 237–246. [Google Scholar] [CrossRef]
- Urbaneja-Bernat, P.; Alonso, M.; Tena, A.; Bolckmans, K.; Urbaneja, A. Sugar as nutritional supplement for the zoophytophagous predator Nesidiocoris tenuis. BioControl 2013, 58, 57–64. [Google Scholar] [CrossRef]
- Ahn, Y.-J.; Kim, Y.-J.; Yoo, J.-K. Toxicity of the herbicide glufosinate-ammonium to predatory insects and mites of Tetranychus urticae (Acari: Tetranychidae) Under Laboratory Conditions. J. Econ. Entomol. 2001, 94, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-I.; Lee, S.-G.; Park, H.-M.; Ahn, Y.-J. Toxicity of plant essential oils to Tetranychus urticae (Acari : Tetranychidae) and Phytoseiulus persimilis (Acari : Phytoseiidae). J. Econ. Entomol. 2004, 97, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.-G.; Choi, B.-R.; Park, H.-M.; Park, C.-G.; Ahn, Y.-J. Fumigant toxicity of plant essential oils to Thrips palmi (Thysanoptera: Thripidae) and Orius strigicollis (Heteroptera: Anthocoridae). J. Econ. Entomol. 2009, 99, 1733–1738. [Google Scholar] [CrossRef]
- Galvan, T.L.; Koch, R.L.; Hutchison, W.D. Toxicity of commonly used insecticides in sweet corn and soybean to multicolored asian lady beetle (Coleoptera: Coccinellidae). J. Econ. Entomol. 2005, 98, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Grutter, T.; Changeux, J.P. Nicotinic receptors in wonderland. Trends Biochem. Sci. 2001, 26, 459–463. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Naheed, N.; Abbaskhan, A.; Musharraf, S.G.; Siddiqui, H. Atta-ur-Rahman Phenolic and other constituents of fresh water fern Salvinia molesta. Phytochemistry 2008, 69, 1018–1023. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, A. A floral fragrance methyl benzoate, is an efficient green pesticide. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mostafiz, M.M.; Jhan, P.K.; Shim, J.K.; Lee, K.Y. Methyl benzoate exhibits insecticidal and repellent activities against Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). PLoS ONE 2018, 1–14. [Google Scholar] [CrossRef]
- Mostafiz, M.M.; Hassan, E.; Shim, J.K.; Lee, K.Y. Insecticidal efficacy of three benzoate derivatives against Aphis gossypii and its predator Chrysoperla carnea. Ecotoxicol. Environ. Saf. 2019, 184, 109653. [Google Scholar] [CrossRef]
- Mostafiz, M.M.; Shim, J.K.; Hwang, H.S.; Bunch, H.; Lee, K.Y. Acaricidal effects of methyl benzoate against Tetranychus urticae Koch (Acari: Tetranychidae) on common crop plants. Pest Manag. Sci. 2020, 76, 2347–2354. [Google Scholar] [CrossRef] [PubMed]
- Fytrou, N.; Ilias, A.; Sklivakis, J.; Tsagkarakou, A. Lethal and sublethal effects of selected insecticides on commercially available natural enemies of whiteflies. IOBC-WPRS Bull. 2017, 125, 19–27. [Google Scholar]
- Bacci, L.; Crespo, A.L.; Galvan, T.L.; Pereira, E.J.; Picanço, M.C.; Silva, G.A.; Chediak, M. Toxicity of insecticides to the sweetpotato whitefly (Hemiptera: Aleyrodidae) and its natural enemies. Pest Manag. Sci. 2007, 63, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Van de Veire, M.; Tirry, L. Side effects of pesticides on four species of beneficials used in IPM in glasshouse vegetable crops: “worst case” laboratory tests. IOBC-WPRS Bull. 2003, 26, 41–50. [Google Scholar]
- Rahman, M.M.; Lim, U.T. Females of Riptortus pedestris (Hemiptera: Alydidae) in reproductive diapause are more responsive to synthetic aggregation pheromone. J. Econ. Entomol. 2016, 109, 2082–2089. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Shekar, C.; Krishna, V. Field studies on efficacy of novel insecticides against Helicoverpa armigera (Hubner) infesting on Chickpea. J. Entomol. Zool. Stud. 2014, 2, 286–289. [Google Scholar]
- SAS Institute Inc. Base SAS 9.4 Procedures Guide, Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Systat Software Inc. Systat.; Systat Software: San Jose, CA, USA, 2013. [Google Scholar]
- Van De Veirk, M. Laboratory test method to evaluate the effect of 31 pesticides on the predatory bug, Orius laevigatus (Heteroptera: Anthocoridae). Entomophaga 1996, 41, 235–243. [Google Scholar] [CrossRef]
- Hassan, S.A.; Bigler, F.; Bogenschütz, H.; Boller, E.; Brun, J.; Calis, J.N.M.; Coremans-Pelseneer, J.; Duso, C.; Grove, A.; Heimbach, U.; et al. Results of the sixth joint pesticide testing programme of the IOBC/WPRS-working group pesticides and beneficial organisms. Entomophaga 1994, 39, 107–119. [Google Scholar] [CrossRef]
- Campolo, O.; Puglisi, I.; Barbagallo, R.N.; Cherif, A.; Ricupero, M.; Biondi, A.; Palmeri, V.; Baglieri, A.; Zappalà, L. Side effects of two citrus essential oil formulations on a generalist insect predator, plant and soil enzymatic activities. Chemosphere 2020. [Google Scholar] [CrossRef]
- Ziaei Madbouni, M.A.; Samih, M.A.; Qureshi, J.A.; Biondi, A.; Namvar, P. Compatibility of insecticides and fungicides with the zoophytophagous mirid predator Nesidiocoris tenuis. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Treatment | Laboratory Condition (Treated Tomato Leaf) | Greenhouse Condition (Treated Tomato Plant) |
|---|---|---|
| Acetamiprid | 80.0 ± 3.8a | 66.7 ± 4.4a |
| MB 1% | 17.8 ± 4.4b | 13.3 ± 3.3b |
| MB 0.5% | 8.9 ± 2.2bc | 6.7 ± 1.7bc |
| MB 0.25% | 4.4 ± 2.2bc | 3.3 ± 1.7bc |
| Negative control | 0.0 ± 0.0c | 0.0 ± 0.0c |
| Treatment | Mean Number B. tabaci Eggs/Treatment | Mean Number of B. tabaci Eggs Consumed Per N. tenuis Adult/3 Days | Reduction in Feeding Rate Relative to Control (%) |
|---|---|---|---|
| Acetamiprid | 212.6 ± 10.1 | 68.0 ± 5.3c | 45.4 |
| MB 1% | 208.3 ± 15.5 | 96.3 ± 4.9b | 22.7 |
| MB 0.5% | 205.3 ± 6.5 | 112.3 ± 4.1ab | 9.8 |
| MB 0.25% | 206.7 ± 13.4 | 116.0 ± 2.8ab | 6.9 |
| Negative control | 211.7 ± 8.6 | 124.6 ± 3.7a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostafiz, M.M.; Hassan, E.; Shim, J.-K.; Lee, K.-Y. Lethal and Sublethal Effects of Methyl Benzoate on the Predatory Bug Nesidiocoris tenuis. Insects 2020, 11, 377. https://doi.org/10.3390/insects11060377
Mostafiz MM, Hassan E, Shim J-K, Lee K-Y. Lethal and Sublethal Effects of Methyl Benzoate on the Predatory Bug Nesidiocoris tenuis. Insects. 2020; 11(6):377. https://doi.org/10.3390/insects11060377
Chicago/Turabian StyleMostafiz, Md Munir, Errol Hassan, Jae-Kyoung Shim, and Kyeong-Yeoll Lee. 2020. "Lethal and Sublethal Effects of Methyl Benzoate on the Predatory Bug Nesidiocoris tenuis" Insects 11, no. 6: 377. https://doi.org/10.3390/insects11060377
APA StyleMostafiz, M. M., Hassan, E., Shim, J.-K., & Lee, K.-Y. (2020). Lethal and Sublethal Effects of Methyl Benzoate on the Predatory Bug Nesidiocoris tenuis. Insects, 11(6), 377. https://doi.org/10.3390/insects11060377