Use of Molecular Gut Content Analysis to Decipher the Range of Food Plants of the Invasive Spotted Lanternfly, Lycorma delicatula



Abstract

1. Introduction
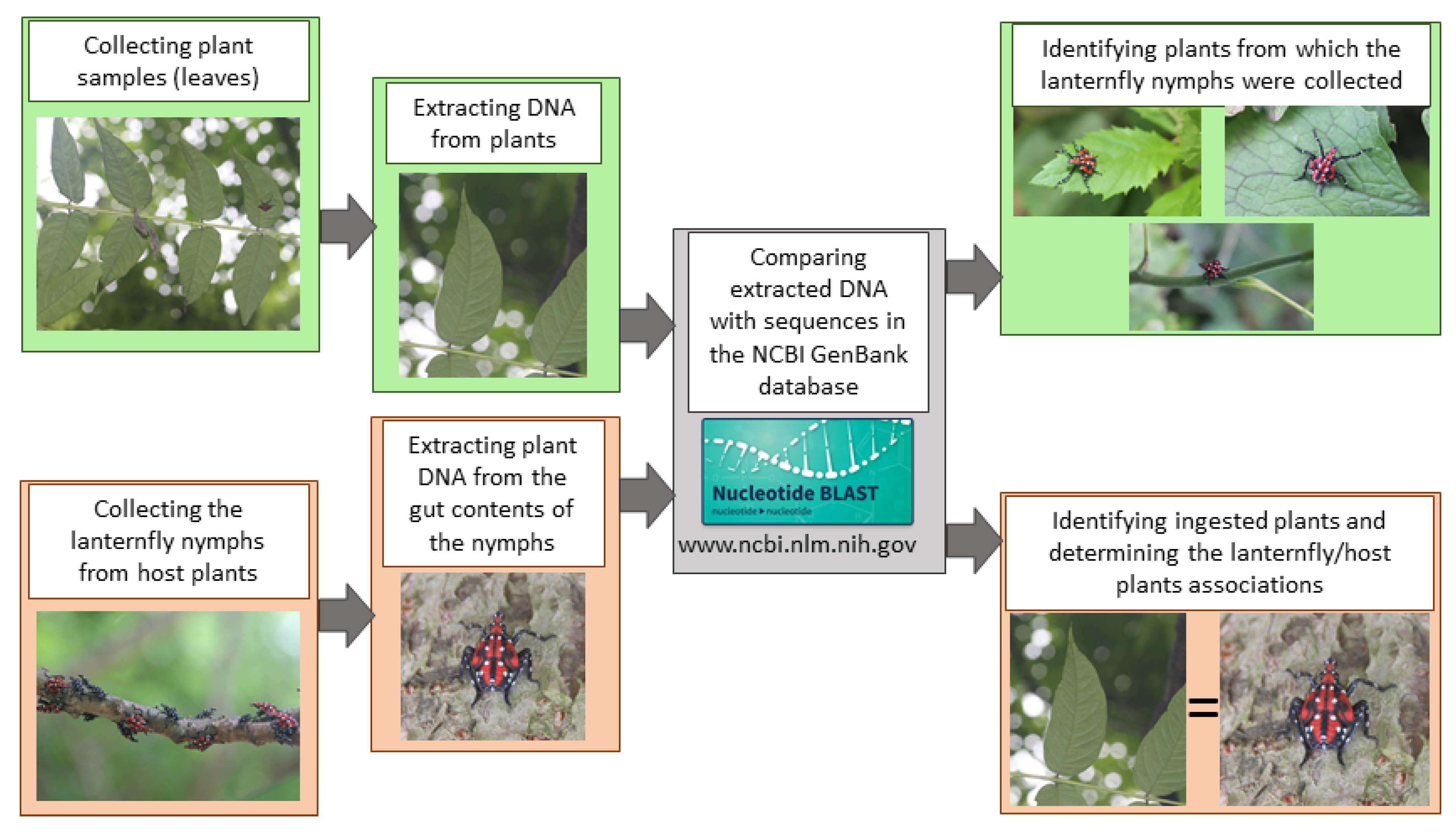
2. Materials and Methods
2.1. Insect and Plant Collection
2.2. DNA Extraction, PCR, and Sequencing
2.3. Data Analysis
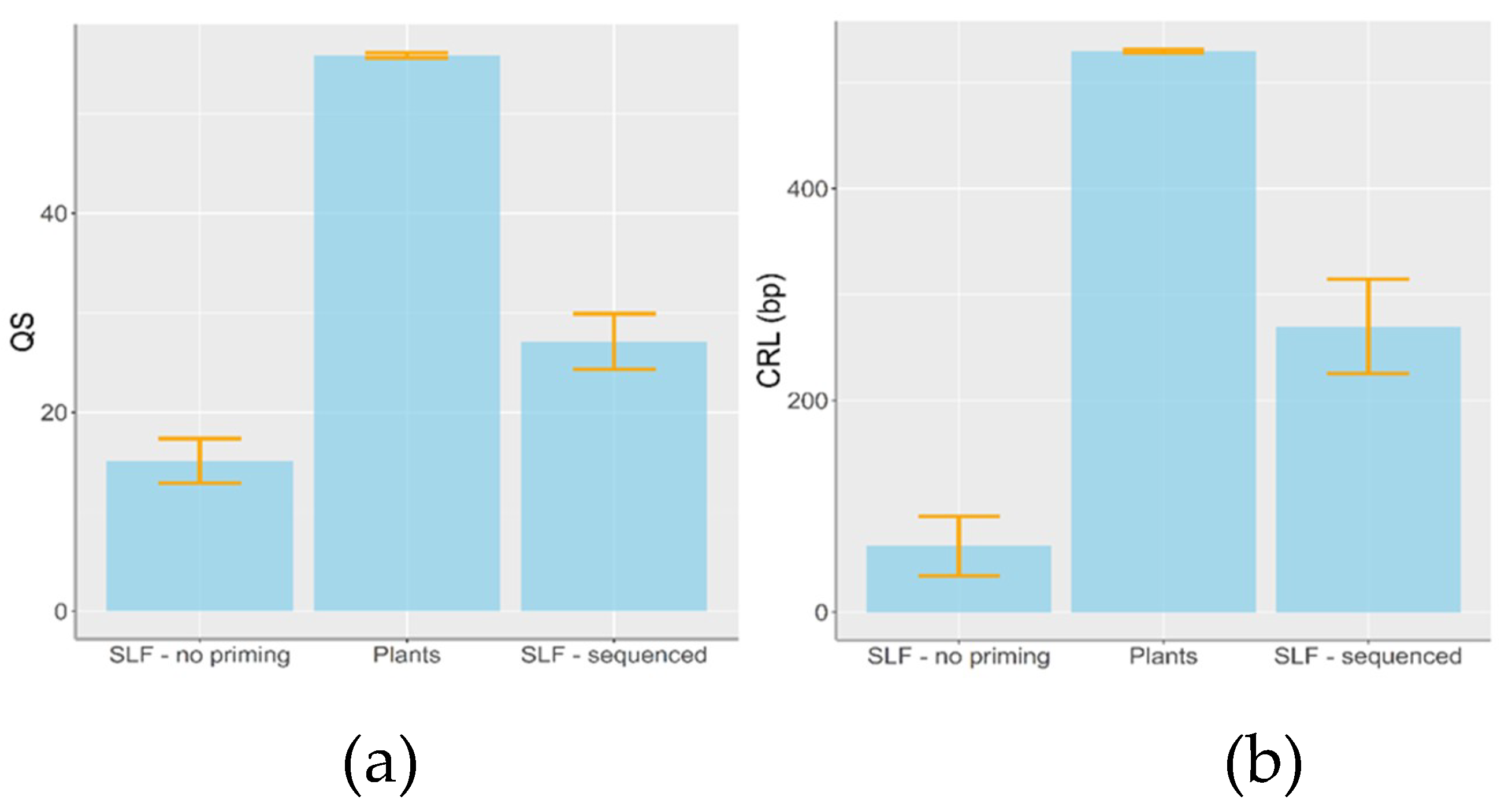
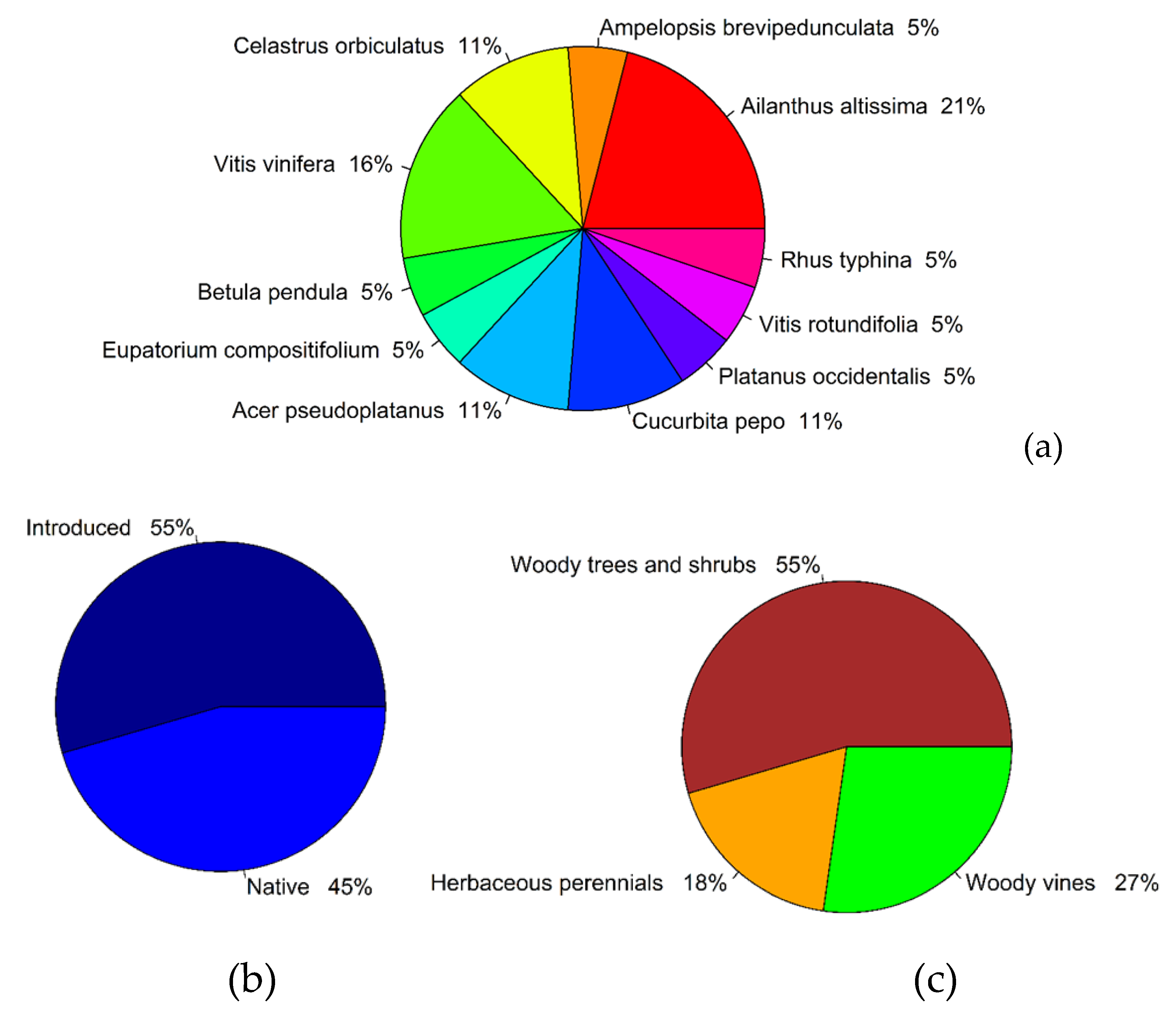
3. Results
4. Discussion
4.1. Plant DNA Detection from the Gut Contents of L. delicatula
4.2. Potential Implications for Host Plant Usage by L. delicatula
4.3. Practical Applications, Methodological Recommendations, and Additional Future Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.G.; Lee, E.H.; Seo, Y.M.; Kim, N.Y. Cyclic behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on host plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The first New World record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Dara, S.K.; Barringer, L.; Arthurs, S.P. Lycorma delicatula (Hemiptera: Fulgoridae): A new invasive pest in the United States. J. Integr. Pest Manag. 2015, 6, 20. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, Y.L.; Leskey, T.C. A review of biology and management of Lycorma delicatula (Hemiptera: Fulgoridae), an emerging global invasive species. J. Asia-Pac. Entomol. 2019, 22, 589–596. [Google Scholar] [CrossRef]
- Liu, H.; Hartlieb, R.J. Spatial Distribution of Lycorma delicatula (Hemiptera: Fulgoridae) Egg Masses on Tree-Of-Heaven, Black Walnut, and Siberian Elm in North America. J. Econ. Entomol. 2019. [Google Scholar] [CrossRef]
- Wakie, T.T.; Neven, L.G.; Yee, W.L.; Lu, Z. The establishment risk of Lycorma delicatula (Hemiptera: Fulgoridae) in the United States and globally. J. Econ. Entomol. 2020, 113, 306–314. [Google Scholar] [CrossRef]
- Jung, J.M.; Jung, S.; Byeon, D.H.; Lee, W.H. Model-Based prediction of potential distribution of the invasive insect pest, spotted lanternfly Lycorma delicatula (Hemiptera: Fulgoridae), by using CLIMEX. J. Asia-Pac. Biodivers. 2017, 10, 532–538. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Wickham, J.; Cleary, K.; Spichiger, S.E.; Zhang, L.; Baker, J.; Carrillo, D. Discovery of three kairomones in relation to trap and lure development for spotted lanternfly (Hemiptera: Fulgoridae). J. Econ. Entomol. 2019, 112, 671–682. [Google Scholar] [CrossRef]
- Liu, H. Oviposition substrate selection, egg mass characteristics, host preference, and life history of the spotted lanternfly (Hemiptera: Fulgoridae) in North America. Environ. Entomol. 2019, 48, 1452–1468. [Google Scholar] [CrossRef]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Al Homaidan, A.A.; Al Farhan, A.H.; Khan, H.A. DNA barcoding of arid wild plants using rbcL gene sequences. Genet. Mol. Res. 2012, 11, 1934–1941. [Google Scholar] [CrossRef]
- De la Cadena, G.; Papadopoulou, A.; Maes, J.M.; Gómez-Zurita, J. Evaluation of bias on the assessment of diet breadth of herbivorous insects using molecular methods. Insect Sci. 2017, 24, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Robledo, C.; Erickson, D.L.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Tropical plant–Herbivore networks: Reconstructing species interactions using DNA barcodes. PLoS ONE 2013, 8, e52967. [Google Scholar] [CrossRef] [PubMed]
- Avanesyan, A. Plant DNA detection from grasshopper gut contents: A step-By-Step protocol, from tissues preparation to obtaining plant DNA sequences. Appl. Plant Sci. 2014, 2, 1300082. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; McNeill, M.R.; Ma, J.; Qin, X.; Tu, X.; Cao, G.; Wang, G.; Nong, X.; Zhang, Z. Gut transcriptome analysis shows different food utilization efficiency by the grasshopper Oedaleous asiaticus (Orthoptera: Acrididae). J. Econ. Entomol. 2017, 110, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gravel, D.; He, F. Seeing is believing? Comparing plant–Herbivore networks constructed by field co-occurrence and DNA barcoding methods for gaining insights into network structures. Ecol. Evol. 2019, 9, 1764–1776. [Google Scholar] [CrossRef] [PubMed]
- Latif, M.A.; Omar, M.Y.; Tan, S.G.; Siraj, S.S.; Ali, M.E.; Rafii, M.Y. Food assimilated by two sympatric populations of the brown planthopper Nilaparvata lugens (Delphacidae) feeding on different host plants contaminates insect DNA detected by RAPD-PCR analysis. Genet. Mol. Res. 2012, 11, 30–41. [Google Scholar] [CrossRef]
- Hereward, J.P.; DeBarro, P.J.; Walter, G.H. Resolving multiple host use of an emergent pest of cotton with microsatellite data and chloroplast markers (Creontiades dilutus Stål; Hemiptera, Miridae). Bull. Entomol. Res. 2013, 103, 611–618. [Google Scholar] [CrossRef]
- Cooper, W.R.; Horton, D.R.; Unruh, T.R.; Garczynski, S.F. Gut content analysis of a phloem-Feeding insect, Bactericera cockerelli (Hemiptera: Triozidae). Environ. Entomol. 2016, 45, 938–944. [Google Scholar] [CrossRef]
- Baker, T.C.; Smyers, E.C.; Urban, J.M.; Meng, Z.; Damadaram, K.P.; Myrick, A.J.; Cooperband, M.F.; Domingue, M.J. Progression of seasonal activities of adults of the spotted lanternfly, Lycorma delicatula, during the 2017 season of mass flight dispersal behavior in eastern Pennsylvania. J. Asia-Pac. Entomol. 2019, 22, 705–713. [Google Scholar] [CrossRef]
- Avanesyan, A.; Culley, T.M. Feeding preferences of Melanoplus femurrubrum grasshoppers on native and exotic grasses: Behavioral and molecular approaches. Entomol. Exp. Appl. 2015, 157, 152–163. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O.; Bermingham, E. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef] [PubMed]
- Sundari, N.; Papuangan, N. Amplification and Analysis of Rbcl Gene (Ribulose-1, 5-Bisphosphate Carboxylase) of Clove in Ternate Island. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Malang, Indonesia, 2019; Volume 276, No. 1; p. 012061. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, No. 41; pp. 95–98. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. Available online: http://www.R-project.org/ (accessed on 20 December 2018).
- Avanesyan, A.; Maugel, T.K.; Lamp, W. External morphology and developmental changes of tarsal tips and mouthparts of the invasive spotted lanternfly, Lycorma delicatula. PLoS ONE 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Bao, W.F.; Yang, F.; Xu, B.; Yang, Y.Z. The specific host plant DNA detection suggests a potential migration of Apolygus lucorum from cotton to mungbean fields. PLoS ONE 2017, 12, e0177789. [Google Scholar] [CrossRef] [PubMed]
- Cooperband, M.F.; Mack, R.; Spichiger, S.E. Chipping to destroy egg masses of the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae). J. Insect Sci. 2018, 18, 7. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Hulme, P.E. How strongly do interactions with closely-Related native species influence plant invasions? Darwin’s naturalization hypothesis assessed on Mediterranean islands. J. Biogeogr. 2006, 33, 1116–1125. [Google Scholar] [CrossRef]
- Avanesyan, A. Should I eat or should I go? Acridid grasshoppers and their novel host plants: Implications for biotic resistance. Plants 2018, 7, 83. [Google Scholar] [CrossRef]
- Wang, Q.; Bao, W.; Zhang, Q.; Fu, X.; Yang, Y.; Lu, Y. Host plant use of a polyphagous mirid, Apolygus lucorum: Molecular evidence from migratory individuals. Ecol. Evol. 2019, 9, 11518–11528. [Google Scholar] [CrossRef]
- Jurado-Rivera, J.A.; Vogler, A.P.; Reid, C.A.; Petitpierre, E.; Gómez-Zurita, J. DNA barcoding insect–Host plant associations. Proc. R. Soc. B Biol. Sci. 2008, 276, 639–648. [Google Scholar] [CrossRef]
- Wang, Q.; Bao, W.; Yang, F.; Yang, Y.; Lu, Y. A PCR-Based analysis of plant DNA reveals the feeding preferences of Apolygus lucorum (Heteroptera: Miridae). Arthropod-Plant Interact. 2018, 12, 567–574. [Google Scholar] [CrossRef]
- Cooper, W.R.; Horton, D.R.; Wildung, M.R.; Jensen, A.S.; Thinakaran, J.; Rendon, D.; Nottingham, L.B.; Beers, E.H.; Wohleb, C.H.; Hall, D.G.; et al. Host and Non-Host ‘Whistle Stops’ for Psyllids: Molecular Gut Content Analysis by High-Throughput Sequencing Reveals Landscape-Level Movements of Psylloidea (Hemiptera). Environ. Entomol. 2019, 48, 554–566. [Google Scholar] [CrossRef]
- Hereward, J.P.; Walter, G.H. Molecular interrogation of the feeding behaviour of field captured individual insects for interpretation of multiple host plant use. PLoS ONE 2012, 7, e44435. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Moon, H.; Sherratt, T.N.; Lee, S.I.; Jablonski, P.G. Multiple lines of anti-Predator defence in the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae). Biol. J. Linn. Soc. 2016, 120, 115–124. [Google Scholar] [CrossRef]
- Song, S.; Kim, S.; Kwon, S.W.; Lee, S.I.; Jablonski, P.G. Defense sequestration associated with narrowing of diet and ontogenetic change to aposematic colours in the spotted lanternfly. Sci. Rep. 2018, 8, 16831. [Google Scholar] [CrossRef] [PubMed]
- Myrick, A.J.; Baker, T.C. Analysis of Anemotactic Flight Tendencies of the Spotted Lanternfly (Lycorma delicatula) during the 2017 Mass Dispersal Flights in Pennsylvania. J. Insect Behav. 2019, 32, 11–23. [Google Scholar] [CrossRef]
- Whitaker, M.R.; Baker, C.C.; Salzman, S.M.; Martins, D.J.; Pierce, N.E. Combining stable isotope analysis with DNA metabarcoding improves inferences of trophic ecology. PloS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Hagler, J.R. It’s Gut Check Time! A Universal Food Immunomarking Technique for Studying Arthropod Feeding Activities. Ann. Entomol. Soc. Am. 2019, 112, 211–219. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Description | Unique Plant Species 1 | Nymphs Gut Contents Sequenced 2 |
|---|---|---|---|
| Location 1 | Private territory (backyard); 7 trees; ~ 12 × 8 m2 -area | Lonicera maackii | Yes |
| Vitis acerifolia | Yes | ||
| Celastrus orbiculatus | Yes | ||
| Rhus typhina | Yes | ||
| Location 2 | Private forest territory; 8 trees; ~ 10 × 10 m2 -area | Ailanthus altissima | Yes |
| Erechtites hieraciifolius | No | ||
| Rubus caesius | No | ||
| Acer platanoides | No | ||
| Acer distylum | Yes | ||
| Phytolacca dioica | No | ||
| Eupatorium serotinum | No | ||
| Location 3 | Private territory; 1 trap tree | Acer rubrum | Yes |
| Location 4 | Private forest territory; randomly chosen tree (preffered host plant) | Ailanthus altissima | Yes |
| # | Instar | Location 1 | Plants at Field Location 2 | Ingested Plant | |||
|---|---|---|---|---|---|---|---|
| Best Match 3 | Match % | Origin 4 | Plant Life Form 4 | ||||
| 1 | 3 | L1 | Lonicera maackii | Vitis vinifera | 100 | introduced | woody vine |
| 2 | 3 | L1 | Vitis acerifolia | Vitis vinifera | 100 | introduced | woody vine |
| 3 | 4 | L1 | Vitis acerifolia | Celastrus orbiculatus | 98.06 | introduced | woody vine |
| 4 | 3 | L1 | Celastrus orbiculatus | Ampelopsis brevipedunculata | 94 | introduced | shrub |
| 5 | 4 | L1 | Rhus typhina | Betula pendula | 100% | introduced | tree |
| 6 | 4 | L1 | Rhus typhina | Eupatorium compositifolium | 85.19 | native | herbaceous perennial |
| 7 | 3 | L1 | Rhus typhina | Vitis vinifera | 100 | introduced | woody vine |
| 8 | 4 | L1 | Rhus typhina | Acer pseudoplatanus | 100 | introduced | tree |
| 9 | 3 | L1 | Rhus typhina | Cucurbita pepo | 96.70% | introduced | herbaceous perennial |
| 10 | 4 | L1 | Rhus typhina | Acer pseudoplatanus | 99% | introduced | tree |
| 11 | 4 | L1 | Rhus typhina | Platanus occidentalis | 95.45% | native | tree |
| 12 | 4 | L2 | Ailanthus altissima | Ailanthus altissima | 99% | introduced | tree |
| 13 | 4 | L2 | Acer distylum | Rhus typhina | 84% | native | tree |
| 14 | 3 | L3 | Acer rubrum | Ailanthus altissima | 96.32% | introduced | tree |
| 15 | 4 | L3 | Acer rubrum | Celastrus orbiculatus | 100% | introduced | woody vine |
| 16 | 4 | L3 | Acer rubrum | Vitis rotundifolia | 95.22% | native | woody vine |
| 17 | 3 | L3 | Acer rubrum | Cucurbita pepo | 85% | introduced | herbaceous perennial |
| 18 | 4 | L4 | Ailanthus altissima | Ailanthus altissima | 99% | introduced | tree |
| 19 | 4 | L4 | Ailanthus altissima | Ailanthus altissima | 98% | introduced | tree |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avanesyan, A.; Lamp, W.O. Use of Molecular Gut Content Analysis to Decipher the Range of Food Plants of the Invasive Spotted Lanternfly, Lycorma delicatula. Insects 2020, 11, 215. https://doi.org/10.3390/insects11040215
Avanesyan A, Lamp WO. Use of Molecular Gut Content Analysis to Decipher the Range of Food Plants of the Invasive Spotted Lanternfly, Lycorma delicatula. Insects. 2020; 11(4):215. https://doi.org/10.3390/insects11040215
Chicago/Turabian StyleAvanesyan, Alina, and William O. Lamp. 2020. "Use of Molecular Gut Content Analysis to Decipher the Range of Food Plants of the Invasive Spotted Lanternfly, Lycorma delicatula" Insects 11, no. 4: 215. https://doi.org/10.3390/insects11040215
APA StyleAvanesyan, A., & Lamp, W. O. (2020). Use of Molecular Gut Content Analysis to Decipher the Range of Food Plants of the Invasive Spotted Lanternfly, Lycorma delicatula. Insects, 11(4), 215. https://doi.org/10.3390/insects11040215