The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Melon Crop
2.4. Flower Margin Composition and Growth
2.5. Visitor Sampling
2.6. Melon Productivity and Quality
2.7. Statistical Analysis
3. Results
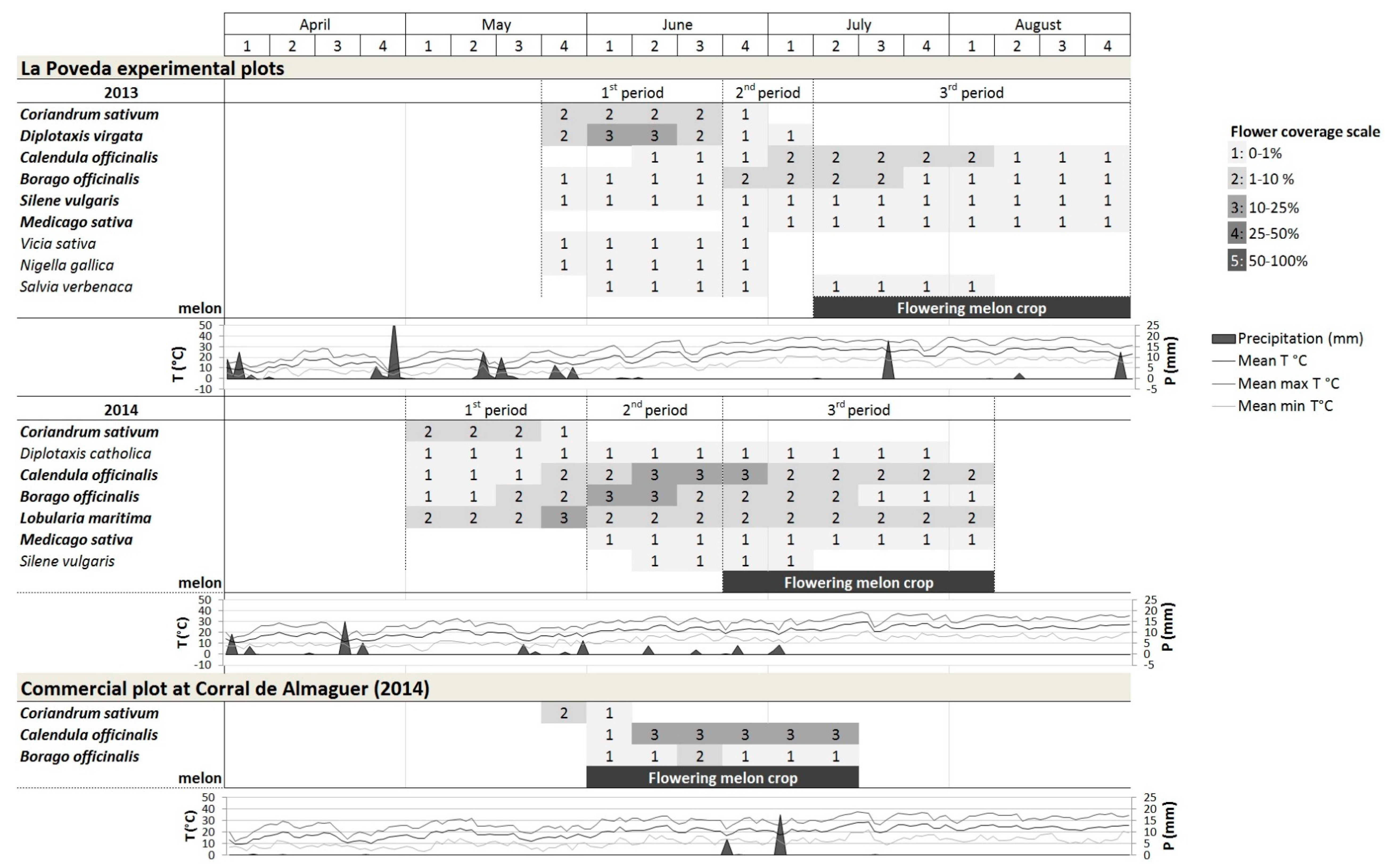
3.1. Bloom Period and Flower Coverage in the Flower Strips
3.1.1. La Poveda Experimental Plots
3.1.2. Commercial Field at Corral de Almaguer
3.2. Visitors
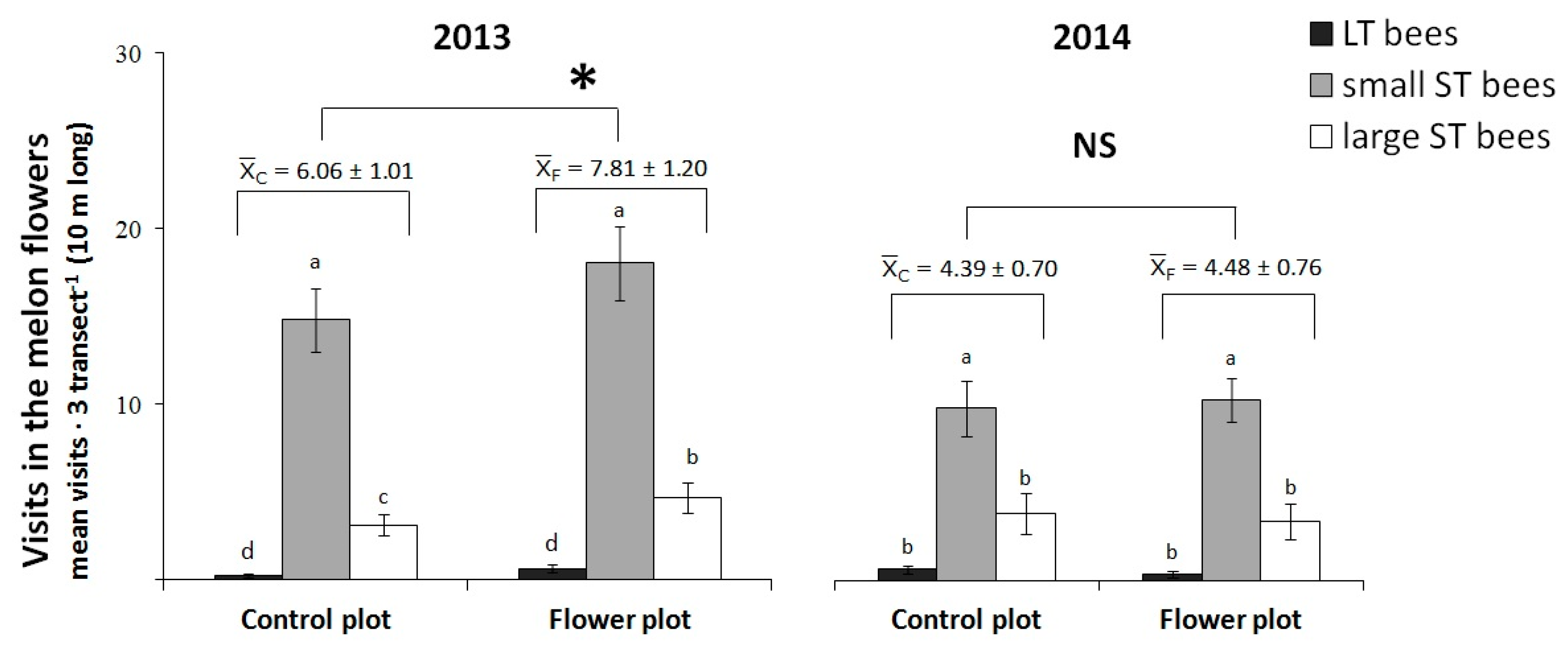
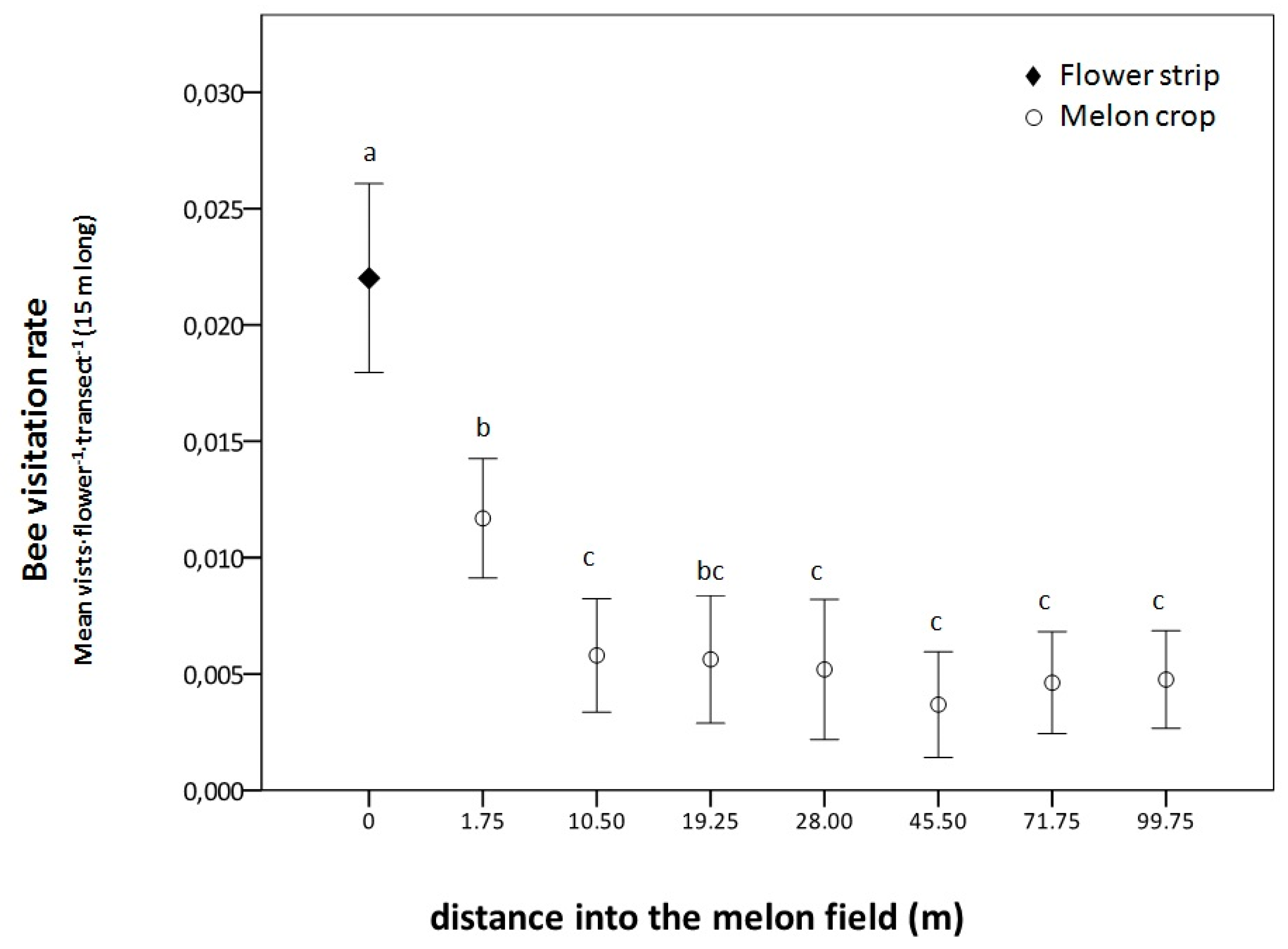
3.2.1. La Poveda Experimental Plots
3.2.2. Commercial Field at Corral de Almaguer
3.3. Melon Quality and Production
4. Discussion
4.1. Selection of Suitable Flowering Plants
4.2. Visits to the Melon Crop and the Role of the Flower Strip as Competitor or Facilitator
4.3. Melon Productivity and Quality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Fontaine, C.; Dajoz, I.; Meriguet, J.; Loreau, M. Functional diversity of plant—Pollinator interaction webs enhances the persistence of plant communities. PLoS Biol. 2006, 4, e1. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Aizen, M.A.; Klein, A.M.; Cunningham, S.A.; Harder, L.D. Global growth and stability of agricultural yield decrease with pollinator dependence. Proc. Natl. Acad. Sci. USA 2011, 108, 5909–5914. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. Long-Term Global Trends in Crop Yield and Production Reveal No Current Pollination Shortage but Increasing Pollinator Dependency. Curr. Biol. 2008, 18, 1572–1575. [Google Scholar] [CrossRef]
- Breeze, T.D.; Vaissière, B.E.; Bommarco, R.; Petanidou, T.; Seraphides, N.; Kozák, L.; Scheper, J.; Biesmeijer, J.C.; Kleijn, D.; Gyldenkærne, S.; et al. Agricultural Policies Exacerbate Honeybee Pollination Service Supply-Demand Mismatches Across Europe. PLoS ONE 2014, 9, e82996. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Cusser, S.; Neff, J.L.; Jha, S. Natural land cover drives pollinator abundance and richness, leading to reductions in pollen limitation in cotton agroecosystems. Agric. Ecosyst. Environ. 2016, 226, 33–42. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Carvalheiro, L.G.; Leonhardt, S.D.; Aizen, M.A.; Blaauw, B.R.; Isaacs, R.; Kuhlmann, M.; Kleijn, D.; Klein, A.M.; Kremen, C.; et al. From research to action: Enhancing crop yield through wild pollinators. Front. Ecol. Environ. 2014, 12, 439–447. [Google Scholar] [CrossRef]
- EC Biodiversity Strategy—Environment—European Commission. Available online: https://ec.europa.eu/environment/nature/biodiversity/strategy/index_en.htm (accessed on 14 August 2017).
- EC European Commission, Agriculture and Rural Development. Greening. Available online: https://ec.europa.eu/agriculture/direct-support/greening_en (accessed on 1 December 2016).
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- BOE Agencia Estatal Boletín Oficial del Estado. Real Decreto 1378/2018. Available online: https://www.boe.es/diario_boe/txt.php?id=BOE-A-2018-15349 (accessed on 30 November 2018).
- Morrison, J.; Izquierdo, J.; Hernández, E.; González-andújar, J.L. Agriculture, Ecosystems and Environment The role of fi eld margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Carvell, C.; Bourke, A.F.G.; Osborne, J.L.; Heard, M.S. Effects of an agri-environment scheme on bumblebee reproduction at local and landscape scales. Basic Appl. Ecol. 2015, 16, 519–530. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Ekroos, J.; Dänhardt, J.; Andersson, G.K.S.; Olsson, O.; Smith, H.G. Sown flower strips in southern Sweden increase abundances of wild bees and hoverflies in the wider landscape. Biol. Conserv. 2015, 184, 51–58. [Google Scholar] [CrossRef]
- Scheper, J.; Bommarco, R.; Holzschuh, A.; Potts, S.G.; Riedinger, V.; Roberts, S.P.M.; Rundlöf, M.; Smith, H.G.; Steffan-Dewenter, I.; Wickens, J.B.; et al. Local and landscape-level floral resources explain effects of wildflower strips on wild bees across four European countries. J. Appl. Ecol. 2015, 52, 1165–1175. [Google Scholar] [CrossRef]
- M’Gonigle, L.K.; Ponisio, L.C.; Cutler, K.; Kremen, C. Habitat restoration promotes pollinator persistence and colonization in intensively managed agriculture. Ecol. Appl. 2015, 25, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Tuell, J.K.; Fiedler, A.K.; Landis, D.; Isaacs, R. Visitation by Wild and Managed Bees (Hymenoptera: Apoidea) to Eastern U.S. Native Plants for Use in Conservation Programs. Environ. Entomol. 2008, 37, 707–718. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Balzan, M.V.; Bocci, G.; Moonen, A.-C. Augmenting flower trait diversity in wildflower strips to optimise the conservation of arthropod functional groups for multiple agroecosystem services. J. Insect Conserv. 2014, 18, 713–728. [Google Scholar] [CrossRef]
- Campbell, A.J.; Biesmeijer, J.C.; Varma, V.; Wäckers, F.L. Realising multiple ecosystem services based on the response of three beneficial insect groups to floral traits and trait diversity. Basic Appl. Ecol. 2012, 13, 363–370. [Google Scholar] [CrossRef]
- Carreck, N.L.; Williams, I.H. Food for insect pollinators on farmland: Insect visits to flowers of annual seed mixtures. J. Insect Conserv. 2002, 6, 13–23. [Google Scholar] [CrossRef]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biol. Control 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Carrié, R.J.G.; George, D.R.; Wäckers, F.L. Selection of floral resources to optimise conservation of agriculturally-functional insect groups. J. Insect Conserv. 2012, 16, 635–640. [Google Scholar] [CrossRef]
- Barbir, J.; Badenes-Pérez, F.R.; Fernández-Quintanilla, C.; Dorado, J. The attractiveness of flowering herbaceous plants to bees (Hymenoptera: Apoidea) and hoverflies (Diptera: Syrphidae) in agro-ecosystems of Central Spain. Agric. For. Entomol. 2015, 17, 20–28. [Google Scholar] [CrossRef]
- Barbir, J.; Azpiazu, C.; Badenes-Pérez, F.R.; Fernández-Quintanilla, C.; Dorado, J. Functionality of Selected Aromatic Lamiaceae in Attracting Pollinators in Central Spain. J. Econ. Entomol. 2016, 4, 1–8. [Google Scholar] [CrossRef]
- M’Gonigle, L.K.; Williams, N.M.; Lonsdorf, E.; Kremen, C. A Tool for Selecting Plants When Restoring Habitat for Pollinators. Conserv. Lett. 2017, 10, 105–111. [Google Scholar] [CrossRef]
- Amy, C.; Noël, G.; Hatt, S.; Uyttenbroeck, R.; Van De Meutter, F.; Genoud, D.; Francis, F. Flower strips in wheat intercropping system: Effect on pollinator abundance and diversity in Belgium. Insects 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Haaland, C.; Naisbit, R.E.; Bersier, L.-F. Sown wildflower strips for insect conservation: A review. Insect Conserv. Divers. 2011, 4, 60–80. [Google Scholar] [CrossRef]
- Kohler, F.; Verhulst, J.; van Klink, R.; Kleijn, D. At what spatial scale do high-quality habitats enhance the diversity of forbs and pollinators in intensively farmed landscapes? J. Appl. Ecol. 2008, 45, 753–762. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2014, 189, 164–170. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Saraiva, A.M.; Giannini, T.C. Establishing Knowledge Management System for Ecological Interactions. The case of crop pollinators. In Pollination Services to Agriculture: Sustaining and Enhancing a Key Ecosystem Service; Gemmill-Herren, B., Ed.; Routledge: New York, NY, USA, 2016; pp. 74–94. [Google Scholar]
- Balzan, M.V.; Bocci, G.; Moonen, A.C. Utilisation of plant functional diversity in wildflower strips for the delivery of multiple agroecosystem services. Entomol. Exp. Appl. 2016, 158, 304–319. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Flower plantings increase wild bee abundance and the pollination services provided to a pollination-dependent crop. J. Appl. Ecol. 2014, 51, 890–898. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F. Getting more power from your flowers: Multi-functional flower strips enhance pollinators and pest control agents in apple orchards. Insects 2017, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Quinn, N.F.; Brainard, D.C.; Szendrei, Z. Floral Strips Attract Beneficial Insects but Do Not Enhance Yield in Cucumber Fields. J. Econ. Entomol. 2017, 110, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Williams, N.M.; Gaines, H.; Ascher, J.S.; Kremen, C. Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J. Appl. Ecol. 2008, 45, 793–802. [Google Scholar] [CrossRef]
- Ganser, D.; Mayr, B.; Albrecht, M.; Knop, E. Wildflower strips enhance pollination in adjacent strawberry crops at the small scale. Ecol. Evol. 2018, 8, 11775–11784. [Google Scholar] [CrossRef]
- Hodgkiss, D.; Brown, M.J.F.; Fountain, M.T. The effect of within-crop floral resources on pollination, aphid control and fruit quality in commercial strawberry. Agric. Ecosyst. Environ. 2019, 275, 112–122. [Google Scholar] [CrossRef]
- Garbach, K.; Long, R.F. Determinants of field edge habitat restoration on farms in California’s Sacramento Valley. J. Environ. Manag. 2017, 189, 134–141. [Google Scholar] [CrossRef] [PubMed]
- MAPA Ministerio de Agricultura Y Pesca, Alimentación y Medio Ambiente. Anuario de Estadística 2018. Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/2018/default.aspx?parte=3&capitulo=07&grupo=6&seccion=21 (accessed on 25 November 2019).
- Bomfim, I.; Freitas, B.; de Aragão, F.; Walters, S. Pollination in cucurbit crops. In Handbook of Cucurbits: Growth, Cultural Practices, and Physiology; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 181–200. [Google Scholar]
- Tschoeke, P.H.; Oliveira, E.E.; Dalcin, M.S.; Silveira-Tschoeke, M.C.A.C.; Santos, G.R. Diversity and flower-visiting rates of bee species as potential pollinators of melon (Cucumis melo L.) in the Brazilian Cerrado. Sci. Hortic. 2015, 186, 207–216. [Google Scholar] [CrossRef]
- Rodrigo Gómez, S.; Ornosa, C.; Selfa, J.; Guara, M.; Polidori, C. Small sweat bees (Hymenoptera: Halictidae) as potential major pollinators of melon (Cucumis melo) in the Mediterranean. Entomol. Sci. 2016, 19, 55–66. [Google Scholar] [CrossRef]
- SIAR Sistema de Información Agroclimática Para el Regadío. Ministerio de Agricultura Y Pesca, Alimentación Y Medio. Ambiente. Available online: http://eportal.magrama.gob.es/websiar/Inicio.aspx (accessed on 15 November 2015).
- Viñuela, E.; Adan, A.; Rodríguez, J.; Hernando, S.; Dorado, J.; Fernández-Quintanilla, C.; Canomanuel, G.; Fereres, A. Provision of ecological infrastructures to increase pollinators and other beneficial organisms in rainfed crops in Central Spain. IOBC/wrps Bull. 2012, 79, 229–233. [Google Scholar]
- Azpiazu, C.; Morales, I.; Adán, Á.; Medina, P.; Fereres, A. Identifying a suitable annual floral mixture and its relative attractiveness to pollinators in Central Spain. IOBC-WPRS Bull. 2017, 122, 79–82. [Google Scholar]
- Flora ibérica Flora ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares. Available online: http://www.floraiberica.es/ (accessed on 25 January 2013).
- Braun-Blanquet, J.; Lalucat Jo, J.; de Bolòs, O. Fitosociología: Bases para el Estudio de las Comunidades Vegetales. Spanish Edition of Pflanzensoziologie: Grundzüge der Vegetationskunde; Blume: Madrid, Spain, 1979; ISBN 8472141748. [Google Scholar]
- Michener, C.D. The Bees of the World, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; ISBN 9780801885730. [Google Scholar]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Schaffers, A.P.; Peeters, T.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Atlas Hymenoptera Atlas Hymenoptera. Available online: http://www.atlashymenoptera.net (accessed on 3 May 2017).
- Ortiz-Sánchez, F.J. Lista actualizada de las especies de abejas en Espana (Hymenoptera: Apoidea: Apiformes). Boletín la Soc. Entomológica Aragon. 2011, 49, 265–281. [Google Scholar]
- Cabello, M.J.; Castellanos, M.T.; Romojaro, F.; Martínez-Madrid, C.; Ribas, F. Yield and quality of melon grown under different irrigation and nitrogen rates. Agric. Water Manag. 2009, 96, 866–874. [Google Scholar] [CrossRef]
- Walters, S.A.; Taylor, B.H. Efects of honey bee pollination on pumpkin fruit and seed yield. HortScience 2006, 41, 370–373. [Google Scholar] [CrossRef]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B.; Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 74, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Goonewardene, L.A. The use of MIXED models in the analysis of animal experiments with repeated measures data. Can. J. Anim. Sci. 2004, 84, 1–11. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Rosa García, R.; Miñarro, M. Role of floral resources in the conservation of pollinator communities in cider-apple orchards. Agric. Ecosyst. Environ. 2014, 183, 118–126. [Google Scholar] [CrossRef]
- Blüthgen, N.; Klein, A.-M. Functional complementarity and specialisation: The role of biodiversity in plant—Pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortiz-Sánchez, F.J. How Bees Respond Differently to Field Margins of Shrubby and Herbaceous Plants in Intensive Agricultural Crops of the Mediterranean Area. Insects 2020, 11, 26. [Google Scholar] [CrossRef]
- Bugg, R.L.; Colfer, R.G.; Chaney, W.E.; Smith, H.A.; Cannon, J. Flower Flies (Syrphidae) and Other Biological Control Agents for Aphids in Vegetable Crops. UC ANR 2008, 8285, 1–25. [Google Scholar]
- Gorenflo, A.; Diekötter, T.; Van Kleunen, M.; Wolters, V.; Jauker, F. Contrasting pollination efficiency and effectiveness among flower visitors of Malva sylvestris, Borago officinalis and Onobrychis viciifolia. J. Pollinat. Ecol. 2017, 21, 62–70. [Google Scholar]
- Colley, M.R.; Luna, J.M. Relative Attractiveness of Potential Beneficial Insectary Plants to Aphidophagous Hoverflies (Diptera: Syrphidae). Environ. Entomol. 2000, 29, 1054–1059. [Google Scholar] [CrossRef]
- Pineda, A.; Marcos-García, M.Á. Use of selected flowering plants in greenhouses to enhance aphidophagous hoverfly populations (Diptera: Syrphidae). Ann. la Soc. Entomol. Fr (N.S.). 2008, 44, 487–492. [Google Scholar] [CrossRef]
- Gumbert, A. Color choices by bumble bees (Bombus terrestris): Innate preferences and generalization after learning. Behav. Ecol. Sociobiol. 2000, 48, 36–43. [Google Scholar] [CrossRef]
- Carreck, N.L.; Williams, I.H. Observations on two commercial flower mixtures as food sources for beneficial insects in the UK. J. Agric. Sci. 1997, 128, 397–403. [Google Scholar] [CrossRef]
- Pisanty, G.; Klein, A.M.; Mandelik, Y. Do wild bees complement honeybee pollination of confection sunflowers in Israel? Apidologie 2014, 45, 235–247. [Google Scholar] [CrossRef]
- Hicks, D.M.; Ouvrard, P.; Baldock, K.C.R.; Baude, M.; Goddard, M.A.; Kunin, W.E.; Mitschunas, N.; Memmott, J.; Morse, H.; Nikolitsi, M.; et al. Food for pollinators: Quantifying the nectar and pollen resources of urban flower meadows. PLoS ONE 2016, 11, e0158117. [Google Scholar] [CrossRef]
- Polidori, C.; Rubichi, A.; Barbieri, V.; Trombino, L.; Donegana, M. Floral resources and nesting requirements of the ground-nesting social bee, Lasioglossum malachurum (hymenoptera: Halictidae), in a Mediterranean semiagricultural landscape. Psyche A J. Entomol. 2010, 2010. [Google Scholar] [CrossRef]
- Fowler, R.E.; Rotheray, E.L.; Goulson, D. Floral abundance and resource quality influence pollinator choice. Insect Conserv. Divers. 2016, 9, 481–494. [Google Scholar] [CrossRef]
- Goulson, D. Foraging Strategies of Insects for Gathering Nectar an Pollen and Implications for Plant Ecology and Evolution. Perspect. Plant Ecol. Evol. Syst. 1999, 2, 185–209. [Google Scholar] [CrossRef]
- Scheper, J.; Holzschuh, A.; Kuussaari, M.; Potts, S.G.; Rundlöf, M.; Smith, H.G.; Kleijn, D. Environmental factors driving the effectiveness of European agri-environmental measures in mitigating pollinator loss—A meta-analysis. Ecol. Lett. 2013, 16, 912–920. [Google Scholar] [CrossRef]
- Heard, M.S.; Carvell, C.; Carreck, N.L.; Rothery, P.; Osborne, J.L.; Bourke, A.F.G. Landscape context not patch size determines bumble-bee density on flower mixtures sown for agri-environment schemes. Biol. Lett. 2007, 3, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, L.G.; Seymour, C.L.; Veldtman, R.; Nicolson, S.W. Pollination services decline with distance from natural habitat even in biodiversity-rich areas. J. Appl. Ecol. 2010, 47, 810–820. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do sown flower strips boost wild pollinator abundance and pollination services in a spring-flowering crop? A case study from UK cider apple orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef]
- Williams, N.M.; Minckley, R.L.; Silveira, F.A. Variation in native bees used for detecting community changes. Conserv. Ecol. 2001, 5, 7. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Floral Nectaries Location | Height (cm) 1 | La Poveda | Corral de Almaguer |
|---|---|---|---|---|---|
| Calendula officinalis L. | Compositae | open | 20–50 | 2013, 2014 | 2014 |
| Coriandrum sativum L. | Umbelliferae | open | 40–60 | 2013, 2014 | 2014 |
| Nigella gallica L. | Ranunculaceae | open | 10–40 | 2013 | 2014 |
| Lobularia marítima L. | Cruciferae | open | 2–20 | 2014 | - |
| Borago officinalis L. | Boraginaceae | partly concealed | 30–70 | 2013, 2014 | 2014 |
| Diplotaxis virgata Cav. | Cruciferae | partly concealed | 50–100 | 2013 | - |
| Diplotaxis catholica L. | Cruciferae | partly concealed | <80 | 2014 | 2014 |
| Medicago sativa L. | Leguminosae | concealed | 30–70 | 2013, 2014 | 2014 |
| Salvia verbenaca L. | Labiatae | concealed | 5–60 | 2013 | 2014 |
| Silene vulgaris Moench. | Caryophyllaceae | concealed | 24–80 | 2013, 2014 | 2014 |
| Vicia sativa L. | Leguminosae | concealed | <80 | 2013 | 2014 |
| Beetles | |||
| Bruchidae | Spermophagussp. | Curculionidae | Apion sp.1 |
| Anthaxia anatolica Chevrolat 1837 1 | Dasytidae | Enicopus calcaratus Kiesenwetter 1859 1 | |
| Cerambycidae | Agapanthia cardui (Linnaeus 1767) 1 | Psilothrix viridicoerulea (Geoffroy 1785) 1 | |
| Agapanthia annularis (Olivier 1795) 1 | Dermestidae | Attagenus fasciatus (Thunberg 1795) 1 | |
| Certallum ebulinum (Linnaeus 1767) 1 | Meloidae | Cerocoma schaefferi (Linnaeus 1758) 1 | |
| Cetoniidae | Oxythyrea funesta (Poda 1761) 1 | Nitidulidae | Meligetes sp.1 |
| Tropinota hirta (Poda 1761) 1 | Oedemeridae | Oedemera podagrariae (Linnaeus 1767) 1 | |
| Chrysomelidae | Altica sp. 1 | Oedemera simplex (Linnaeus 1767) 1 | |
| Clytra sp. 1 | Tenebrionidae | Heliotaurus ruficollis (Fabricius 1781) 1,2 | |
| Coccinellidae | Coccinella septempunctata Linnaeus 1758 1,2 | ||
| Hoverflies | |||
| Syrphidae | Ceriana vespiformis (Latreille 1804) 1 | Scaeva sp.1,2 | |
| Episyrphus balteatus (De Geer, 1776) 1,2 | Sphaerophoria rueppellii (Weidemann, 1820) 1 | ||
| Eristalis tenax (Linnaeus, 1758) 1,2 | Sphaerophoria scripta (Linnaeus, 1758) 1,2 | ||
| Eupeodes corollae (Fabricius, 1794) 2 | |||
| Long-Tongue (L-T) bees | |||
| Apidae | Amegilla quadrifasciata (de Villers, 1789) 1,2 | Eucera notata Lepeletier, 1841 1 | |
| Anthophora agama Radoszkowski, 1869 1 | Habropoda zonatula Smith, 1854 1 | ||
| Anthophora atroalba Lepeletier, 1841 1 | Xylocopa violacea (Linnaeus, 1758) 1,2 | ||
| Anthophora fulvitarsis Brullé, 1832 1 | Megachilidae | Anthidium florentinum(Fabricius, 1775)1 | |
| Apis melliferaLinnaeus, 17581,2 | Coelioxys echinata Förster, 1853 *,1 | ||
| Bombus terrestris (Linnaeus, 1758) 1,2 | Hoplitis antigae (Pérez, 1895) * 1 | ||
| Ceratina chalcitesGermar, 1839 *,1 | Hoplitis sp.*,1 | ||
| Ceratina cucurbitina(Rossi, 1792) *,1,2 | Megachile pilidens Alfken, 1924 1,2 | ||
| Ceratina nigrolabiata Friese, 1896 1 | Megachile rotundata (Fabricius 1787) 1 | ||
| Eucera elongatula Vachal, 1907 1,2 | Megachile versicolor Smith, 1844 1 | ||
| Small Short-Tongue (S-T) bees | |||
| Andrenidae | Andrena bicolorFabricius, 1775 2 | Halictus maculatusSmith, 18481 | |
| Andrena djelfensis Pérez, 1895 1,2 | Halictus seladonius (Fabricius, 1794) 1 | ||
| Andrena tenuistriata Pérez, 1895 1 | Halictus tetrazonius Klug in Germar, 1817 1 | ||
| Andrena sp.11 | Halictus sp. 1 1 | ||
| Andrena sp.2 1 | Halictus sp. 2 1 | ||
| Panurgus calcaratus (Scopoli, 1763) 1 | Lasioglossum discum (Smith 1853) 1 | ||
| Panurgus canescens Latreille, 18111 | Lasioglossum leucozonium (Schrank, 1781) 1,2 | ||
| Panurgus sp. 1 | Lasioglossum clypeare (Schenck 1853) 1 | ||
| Halictidae | Ceylalictus variegatus (Fabricius, 1798) 1 | Lasioglossum malachurum(Kirby, 1802) 1,2 | |
| Halictus crenicornis Blüthgen 1923 1 | Lasioglossum minutulum(Schenck 1853) 1,2 | ||
| Halictus gemmeus Dours, 1872 1 | Sphecodes croaticus Meyer 1922 1 | ||
| Large Short-Tongue (S-T) bees | |||
| Andrenidae | Andrena albopunctata ssp. melona Warncke, 1967 1,2 | Halictus quadricinctus (Fabricius, 1776) 1,2 | |
| Andrena bicolorata (Rossi, 1790) 1 | Halictus rubicundus(Christ, 1791)1 | ||
| Andrena bimaculata (Kirby, 1802) 1 | Halictus scabiosae(Rossi, 1790)1,2 | ||
| Andrena carbonaria (Linnaeus, 1767) 1,2 | Halictus tridivisus Blüthgen, 1924 1 | ||
| Andrena flavipesPanzer, 17991 | Lasioglossum aegyptiellum (Strand, 1909) 1 | ||
| Andrena florea Fabricius, 1793 1 | Lasioglossum albocinctum(Lucas 1846) | ||
| Andrena nigroaenea (Kirby, 1802) 1 | Lasioglossum pygmaeum(Schenck, 1853)1,2 | ||
| Andrena ovatula (Kirby, 1802) 1 | Pseudapis bispinosa (Brullé, 1832) 1,2 | ||
| Andrena thoracica (Fabricius, 1775) 1,2 | Pseudapis diversipes (Latreille 1806) 1 | ||
| Panurgus banksianus (Kirby, 1802) 1 | Sphecodes albilabris (Fabricius, 1793) 1 | ||
| Halictus asperulus Pérez, 1895 1 | Sphecodes gibbus (Linnaeus, 1758) 1 | ||
| Halictidae | Halictus consobrinus (Perez, 1895) | Sphecodes gibbus (Linnaeus, 1758) 1 | |
| Halictus crenicornis Blüthgen, 1923 1,2 | Melittidae | Dasypoda visnaga (Rossi, 1790) 2 | |
| Halictus fulvipes(Klug, 1817)1 | |||
| Year | 1st Bloom Period | 2nd Bloom Period | 3rd Bloom Period | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2013 | df | F | p | df | F | p | df | F | p |
| Visitor groups (V) | 471.2 | 32.15 | <0.001 | 439.1 | 79.88 | <0.001 | 222.9 | 68.46 | <0.001 |
| Plant species (S) | 3101.5 | 5.70 | 0.001 | 339.9 | 35.89 | <0.001 | 326.2 | 141.33 | <0.001 |
| V × S | 1271.9 | 19.55 | <0.001 | 1239.0 | 7.26 | <0.001 | 622.9 | 20.94 | <0.001 |
| N flowers | 1141.8 | 37.01 | <0.001 | 141.5 | 10.18 | 0.002 | 190.5 | 70.38 | <0.001 |
| 2014 | |||||||||
| Visitor groups (V) | 448.4 | 3.95 | 0.007 | 437.4 | 18.17 | <0.001 | 259.8 | 39.21 | <0.001 |
| Plant species (S) | 372.8 | 4.03 | 0.010 | 346.8 | 57.99 | <0.001 | 364.3 | 24.46 | <0.001 |
| V × S | 1248.3 | 3.06 | 0.003 | 1237.3 | 18.48 | <0.001 | 659.8 | 10.47 | <0.001 |
| N flowers | 1135.9 | 5.24 | 0.024 | 1110.0 | 24.02 | <0.001 | 196.8 | 17.99 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azpiazu, C.; Medina, P.; Adán, Á.; Sánchez-Ramos, I.; del Estal, P.; Fereres, A.; Viñuela, E. The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop. Insects 2020, 11, 66. https://doi.org/10.3390/insects11010066
Azpiazu C, Medina P, Adán Á, Sánchez-Ramos I, del Estal P, Fereres A, Viñuela E. The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop. Insects. 2020; 11(1):66. https://doi.org/10.3390/insects11010066
Chicago/Turabian StyleAzpiazu, Celeste, Pilar Medina, Ángeles Adán, Ismael Sánchez-Ramos, Pedro del Estal, Alberto Fereres, and Elisa Viñuela. 2020. "The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop" Insects 11, no. 1: 66. https://doi.org/10.3390/insects11010066
APA StyleAzpiazu, C., Medina, P., Adán, Á., Sánchez-Ramos, I., del Estal, P., Fereres, A., & Viñuela, E. (2020). The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop. Insects, 11(1), 66. https://doi.org/10.3390/insects11010066