Abstract
After the first record in 2008 in Southeast Italy, the alien invasive and quarantine pest Aleurocanthus spiniferus (orange spiny whitefly—OSW) has gradually spread throughout Europe, infesting several new host plants in addition to the known hosts. Molecular characterization of some Italian populations and a newly found Albanian population highlighted two different haplotypes invading Europe, belonging to one of the haplogroups previously recorded in China. A predator was recorded for the first time in several fields in Italy in association with OSW and other whitefly species. It was successively identified through a morpho-molecular characterization as a Nearctic member of the tribe Serangiini, the ladybird beetle, Delphastus catalinae. This predator represents a promising biocontrol agent to manage A. spiniferus outbreaks in Italy and other invaded countries.
1. Introduction
In Europe, more than 14,000 alien species have been recorded [1], half of which have become invasive. Their number is continuously rising [2] with a simultaneous increase in their diffusion rate [3]. Due to habitat fragmentation creating abundant and diverse niches, Italy is one of the most welcoming territories in Europe for foreign species. Many invasive insect species have been recently reported from Italy [4,5,6,7,8].
Aleurocanthus spiniferus (Quaintance) (Hemiptera: Aleyrodidae), the orange spiny whitefly (OSW), originating from China and South and Southeast Asia, is one of the serious pests infesting citrus [9]. Since its description [10], in the span of a century, OSW spread throughout Asia, Africa, Australia, and in the Pacific islands [11,12,13,14,15,16]. OSW was reported for the first time in the European and Mediterranean Plant Protection Organization (EPPO) areas after its first detection in the Lecce District (Apulia region, Southeast Italy) in 2008 [17]. Since then, OSW spread in the Apulia region, invading other municipalities neighboring Lecce [18] and, expanding northward, reached Brindisi and the districts of Bari and Taranto [15,19]. Although the spread of this pest was limited solely to the south-eastern area of Italy for about a decade, in June 2017 A. spiniferus was found in Salerno (Southwest Italy) [20]. Concurrently, OSW was also detected in the Balkan Peninsula including Croatia (2012), Montenegro (2013), and Greece (2016) [16,21,22].
OSW is considered one of the major threats to citrus production in Asia, Australasia, and the Nearctic zone [14,15,23]. The risk is mainly related to its high polyphagy as well as its self-spreading ability. OSW infests about 90 plant species belonging to 38 different plant families. In the area of the first European record, OSW was found on several hitherto unreported host-plants, among which some ornamentals were economically relevant: Hedera helix L., Laurus nobilis L., Punica granatum L., Malus spp., and Prunus spp. [15].
OSW infestations can weaken plants due to both direct and indirect damage ascribable to sap loss and the production of honeydew respectively. The remarkable amount of excreted honeydew encourages the growth of sooty mould, having negative effects on the photosynthetic process due to the copious soiling of the leaf surface [11,13].
Chemical control against OSW is not effective [24] and, in many cases, the frequent use of chemicals can adversely influence the natural enemy populations. The improper timing of treatments seems to be counter-productive, increasing the severity of infestation [25], probably due to side effects of pesticides on beneficial organisms.
The success of natural enemies against OSW in classical biological control (CBC) programs is widely recognized [11,26,27,28]. Several studies conducted in the native OSW territories highlighted the presence of a large group of natural enemies, including predators, parasitoids, and pathogens [29]. Predators recorded on A. spiniferus include species belonging to Diptera, Neuroptera, and a dozen ladybeetles [30]. However, most of the listed species have strongly polyphagous behavior [29] and OSW enemies also control the congeneric Aleurocanthus woglumi [15,30,31]. However, the complex of useful organisms that control OSW has been enriched because two other species native to the Palearctic region were recorded as preying on OSW in Italy: Clitostetus arcuatus (Rossi) (Coleoptera: Coccinellidae: Coccidulinae) [15], which is a specialist predator of whiteflies, and, sporadically, Oenopia conglobata (L.) (Porcelli, Pers. Comm.), which mainly preys on aphids (Hemiptera: Aphididae) and psyllids (Hemiptera: Psyllidae) [32,33].
Focusing on hymenopteran parasitoids, more than 10 wasp species were collected on OSW populations around the world [29,30,31]: most of them belong to the Aphelinidae family (Ablerus connectans Silvestri, Encarsia smithi (Silvestri), and Eretmocerus spp.), whereas only one Platygastridae (Amitus hesperidum Silvestri) was recorded. In the country of origin, some entomopathogenic fungi have been reported [29,34,35,36], but their role is still not well defined probably due to their poor specificity.
The recent OSW findings in several localities in Italy outline the relentless progress of its spread in the country [20]. Hence, we aimed to define biological and ethological aspects of OSW through: (1) providing an update of the distribution of OSW in the EPPO area, (2) revising the host plant list in the new areas of colonization, (3) evaluating the existence of genetic variability between populations from different areas and different host plants, and (4) finding and characterizing natural enemies in the newly infested areas with the object of evaluating the control of invasive populations of OSW.
2. Materials and Methods
2.1. Monitoring Activities
Since the first record in Southwest Italy (June 2017), monitoring activities were regularly conducted during the 2017–2019 to assess the presence and the spread of OSW in Campania [37]. Already known host plants were checked for the presence of all developmental stages in specialized and non-specialized orchards, private and urban gardens, ornamentals, and park areas. Similar inspections were completed of non-host plants close to infested plants, especially on wild plants or in abandoned fields, to evaluate the infestation of new plant species unrecorded as a suitable host. Unofficial monitoring was completed in places visited for other activities both in Italy and abroad.
Leaves infested by OSW were collected during the monitoring period, placed in sealed plastic bags in a refrigerated container, and carried to the laboratory. OSW samples were collected in eight localities from different host plants (Table 1). Specimens of different young developmental stages were removed with the help of a brush from the leaves, killed in absolute ethanol, and stored at −20 °C until analysis. Beetles, wasps, lacewings, and flies found on the A. spiniferus colonies were collected and placed on OSW-infested leaves in Petri dishes (25 ± 2 °C; 55% relative humidity, RH) to assess their role as natural enemies. Once we determined their ability or lack thereof against OSW, the inspected insects were collected or discarded, respectively. Collected specimens were treated as OSW specimens until analysis.

Table 1.
Information about the specimen involved in this study and respective haplotyping results and sequences accession numbers. FG, Foggia; NA, Naples.
2.2. Morpho-Molecular Characterization
Samples were collected on all different recorded host species and on different plants in eight localities to evaluate the genetic diversity. For each locality and host species, a maximum of five specimens were used for the molecular analysis (Table 1).
Aleurocanthus spiniferus DNAs were extracted from each specimen using a non-destructive Chelex 100 (Bio-Rad, Richmond, CA, USA) and proteinase-K-based method as reported by Gebiola et al. [38].
After DNA extraction, OSW samples were rinsed in deionized water, slide-mounted as described by Cioffi [15], and identified following the relevant taxonomic descriptions [31,39,40,41,42,43].
The mitochondrial gene cytochrome c oxidase subunit I (COI) was amplified using the primer pair AsFmik and AsR4mik [44] with the PCR profile reported by Uesugi [9].
For samples of Delphastus catalinae Horn 1895, due to the absorption of the solution during the DNA extraction process, the method described by Gebiola et al. [38] was slightly modified as follows. We used 6 μL of 20 mg/mL proteinase-K and 100 μL 5% Chelex 100 suspension to obtain DNA from single wild beetles collected in some fields in Campania and Sicily, feeding on OSW and Aleurothrixus floccosus (Mask.) colonies, respectively (Table 2). Samples supplied from two biofactories and commercialized as D. catalinae and D. pusillus (LeConte, 1852) (Table 2) were included in this study. Some samples were previously observed through the use of a Cryo-SEM (Hitachi (Tokyo, Japan) TM 3000 series). This methodology does not require critical point drying or metal coating, and the same observed sample can be later submitted to DNA extraction or/and conventional slide mounting.
Table 2.
Beetle specimens used in this study.
Once rinsed in deionized water, some beetle specimens were dissected and mounted on slides using balsam-phenol as a permanent medium; others were mounted on cards.
Extracted DNA was employed to amplify a portion of the mitochondrial gene COI using the forward primer C1-J-2183 with the reverse primer TL2-N-3014 [45] following the thermocycler conditions described in Gebiola et al. [38].
PCR products were checked on a 1.2% agarose gel stained with GelRED® (Biotium, Fremont, CA, USA) and directly sequenced. Sequences were assembled and edited by eye with Bioedit 7.2.5 [46], and were virtually translated into the corresponding amino acid chain to detect frame-shift mutations and stop codons, using EMBOSS Transeq (http://www.ebi.ac.uk/Tools/st/emboss_transeq/ (accessed 23 October 2019)). Edited sequences were checked against the GenBank database and were submitted to the GenBank database under accession numbers reported in Table 1 and Table 2.
Aleurocanthus spiniferus COI genetic distances and standard errors (SE) were calculated with MEGA 6 software [47] as uncorrected p-distance considering homologous sequences of OSW available in GenBank (accessed 23 October 2019).
The relationships between A. spiniferus specimens were also investigated using Statistical Parsimony in TCS 1.21 [48] on the COI dataset.
OSW phylogeny was reconstructed using maximum likelihood (ML) in RAxML 7.0.4 [49]. A GRT+G+I nucleotide substitution model was used, as selected by jModeltest [50]. ML branch support was based on 1000 rapid bootstrap pseudoreplicates, and clades were considered supported when bootstrap values were >70%. Homologous sequences available in GenBank were included in the alignment and the tree was rooted including the congeneric A. camelliae (Kanmiya & Kasai) sequences.
3. Results
3.1. Monitoring Activities
The survey results indicated that in the new areas of colonization, A. spiniferus was recorded on the main elective hosts Citrus spp. and on the already known host plants, Ceratonia siliqua L., Eriobotrya japonica (Thunb.) Lindl., Hedera helix L., Morus alba L., Prunus armeniaca L., Punica granatum L., Rosa spp., and Vitis vinifera L. OSW was collected from several additional host plants belonging to several botanical families that represent new associations (Table 3).
Table 3.
Additional host-plant species found infested by A. spiniferus in the present study.
In October and December 2017, findings in two different sites in Salerno of some small coleopteran belonging to the Coccinellidae revealed some small ladybeetles preying on A. spiniferus populations infesting leaves of C. limon and R. banksiae. Subsequent surveys in the same areas resulted in the collection of all ladybird developmental stages, from eggs to adults.
3.2. OSW Characterization
Mitochondrial COI sequencing revealed the presence of two haplotypes in the sampled A. spiniferus (Table 1). BLAST search revealed that both obtained haplotypes belong to mitochondrial haplogroup 2 [9] corresponding to the haplotypes H1 and H2 recently found in Greece, and Greece, Italy, and Montenegro, respectively [16].
Haplotype H1 was obtained from samples collected in Bari, Rome, Mattinata (Foggia), San Gennaro Vesuviano (Naples) and in four out five samples from Pescara. Haplotype H2 was detected in other samples from Bari and Pescara, and in all analyzed samples from Salerno, Portici (Naples), and Buthrotum (AL). The percentages of detection of haplotypes were 57% for H1 and 43% for H2. MEGA analyses highlighted that the mean intra-group distances existing between the haplotypes belonging to the haplogroup 2 was 0.31% (±0.002% SE), and 0.8% (±0.003%) in haplogroup 1. Focusing on haplogroup 2, the distance between H1 and H2 sequences recovered in this study was 0.6% (±0.003%), corresponding to four variable and parsimony-informative sites. The inter-group distance between haplogroups 1 and 2 was 12.1% (±0.017%).
Phylogenetic reconstruction resulted in an ML tree (Figure 1) where the two haplogroups were identified in two highly supported clades. The statistical parsimony with TCS yielded two separate networks, corresponding to the haplogroup 1 and haplogroup 2. The connection limit necessary to obtain a single network was 77 steps.
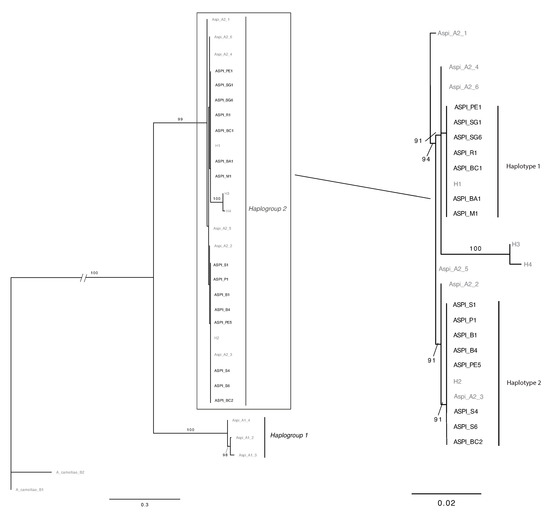
Figure 1.
Maximum likelihood trees based on COI sequences of A. spiniferus. Bootstrap values >70% are shown above the branches. Complete tree (left) and zoom on phylogenetical relationships in Haplogroup 2 (right). Sequences from [9] and [16] are in grey; sequences obtained in this work are in black.
3.3. Natural Enemies
One single predator was reared from A. spiniferus colonies and a preliminary morphological identification identified the collected ladybeetles as D. catalinae. The sequenced region of the mitochondrial COI gene of the Italian sample and those provided by biofactories (Table 3) were identical to each other. The BLAST analysis of this COI sequence showed a 100% similarity to the D. catalinae sequence presents in GenBank (Accession number MF152800).
4. Discussion
Aleurocanthus spiniferus was found on several host plant species in the new areas of colonization confirming previous surveys [15]; in addition, 11 new hosts were found. These findings highlight the already known polyphagy of OSW, which could accelerate its spread in territories where the main hosts are absent, permitting its quick spread in Italy and Albania. This is the first record of OSW in Albania.
In the present study, only two haplotypes (H1 and H2) were found, different from what was recently found in Greece, where four different haplotypes were recorded [16]. Therefore, some conclusions are possible (Figure 2):
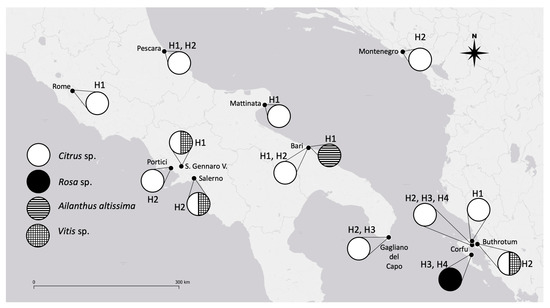
Figure 2.
A. spiniferus distribution, mitochondrial haplotypes, and respective host plants in EPPO zone. EPPO, European and Mediterranean Plant Protection Organization areas.
- (1)
- The H4 haplotype seems to have a reduced diffusion (present only in Greece) [16] because it was not found in any other of the collection areas (Italy and Albania).
- (2)
- The H3 haplotype was previously found in Apulia [16] in the area of the first interception of OSW but it was not found in the present study. This could be due to two possible causes: a poor diffusion of this haplotype (20% of the survey in Italy) [16] and a predominance of the other haplotypes.
- (3)
- The H1 haplotype was recovered in Apulia (both in Bari and in Mattinata) during our sampling but it was not found in the study performed by Kapantaidaki [16]. The different compositions of haplotypes in the samples collected by different authors could be linked both to different sampling methodologies and/or to a patch-like distribution of the different haplotypes.
The reduced genetic variability of OSW specimens collected in the EPPO area (four haplotypes), in the present and previous work [16], compared to those found in the native country (12 haplotypes) [9] may be due to the founder effect that affects invasive species. This is a common pattern for invasive species whose population is established by a few specimens [4,51,52,53]. Interestingly, three out of four haplotypes (H1, H3, and H4) are not found in the country of origin [9,16]. Among them, H1 is the most widespread in European invaded countries. This scenario is similar to the spread of the eucalyptus gall wasp, Leptocybe invasa Fisher and La Salle, in which the main globally spreading haplogroup was never found in the native territories [54]. H4 was found only in Greece, where several different haplotypes have also been found. This finding suggests that the population that invaded Greece may act as a bridgehead for the subsequent introductions to the other countries; however, based on the invasive history of OSW in Europe, this scenario is not temporally plausible [16]. Therefore, the most well-founded hypothesis is that multiple introductions of this species have occurred in the EPPO area. However, for a definitive confirmation of this hypothesis, a wider sampling is necessary.
Our results definitively exclude the possibility that a specificity of a haplotype exists for a host plant because both haplotypes were collected on several different host plants (Table 1 and Figure 2).
Differently, during the growing season, A. altissima hosted only specimens with mt-H1, whereas Vitis sp. hosted solely a population with the mt-H2 haplotype. Both Vitis sp. and A. altissima are deciduous plants and, therefore, during the winter, they cannot host OSW populations, so these species are re-colonized only in the next spring. However, their role (as that of other deciduous trees hosting OSW) is probably crucial in the increase in OSW populations. In spring–summer, such plants could produce a “flywheel effect” in increasing the adult population overwintering on evergreen plants, increasing the chances of survival of winter, thus enabling severe infestations during the following spring.
The finding of OSW on A. altissima requires more detailed studies. The only whitefly reported on the tree of heaven is Dialeurodes citri (Ashmead), whereas congeneric species A. excelsa (Roxb.) seems to be a well-known host for at least four different genera of whiteflies [30,55,56] and OSW is not included.
Studies of genetic distances, supported by phylogenetic analysis and statistical parsimony, indicated a high genetic distance between the two haplogroups (12.1%). These results are consistent with a previous study [9], providing strong indication that the two haplogroups should be reevaluated through an integrative approach because they could result in different species. An integrative approach, considering other molecular markers, biological characteristics, and morphometric analyses, often allows the delimitation and description of different species previously considered single species [53,57,58,59]. The genetic diversity could have important implications in the management of the pest because natural enemies could have different specificity toward distinct cryptic species [60]; for example, the different biology of two pests could affect the approach necessary for their management [61].
During our survey, only a single predator (D. catalinae) was recorded and our samplings highlighted the presence of different developmental stages of D. catalinae feeding on A. spiniferus populations.
Delphastus catalinae is a polyphagous species that has been previously recorded on several prey species: A. floccosus, Pealius kelloggi (Bemis), Dialeurodes citri, D. citrifolii (Morgan), Bemisia tabaci (Genn.), Aleurodicus dispersus Russel, Trialeurodes vaporariorum (Westwood), and A. woglumi [62,63].
The genus Delphastus Casey, belonging to the tribe Serangiini, is native to the Nearctic region and does not include any species native to Europe [62,63]. All the members of the tribe are obligate whitefly predators [62,64,65] and, due to their use as biological control agents, are mass-produced in the USA [66]. Gordon [62] defined D. catalinae distribution as “an artificial distribution that includes South, Central, and North America, as well as the Canary Islands and Hawaii […] probably results from commercial trade”. The sequences of all the examined specimens, even if of different origins, were all identical, which could be evidence that the population found in Italy was derived from field releases and therefore from a biofactory. Booth and Polaszek [63] based their comments about the species on additional laboratory material from Israel and the Netherlands (cultures from Israel) and the U.K. (cultures from Canada). In addition, a similar species, D. pusillus, was released in several augmentative biological program attempts [67,68]. However, because our results demonstrated that the species considered D. pusillus reared in commercial insectaries was instead D. catalinae, only the latter was probably to date used in biological control programs [63,69]. Correct identification of species reared and employed in the biological control program is crucial [70]. However, the identification (especially of small and live specimens) can lead to rearing and introducing incorrectly identified species if recently revised identification protocols are not used or the identification is not confirmed by a taxonomic specialist. An erroneous release could also occur when biofactories either rear congeneric species or introduce wild specimens to avoid the negative effect of prolonged inbreeding [58]. Several problems related to the small size of this species, some clearing and mounting artefacts, and to the high release of D. pusillus in several augmentative biological programs around the world, necessitate a re-description of the species.
As well as other members of the tribe Serangiini, the genus has a small body that is ovoid and strongly convex at the dorsum [70]. In particular, D. catalinae presents a one-segmented antennal club, 2.2 times longer than wide (Figure 3a); maxillary palp with apical segment conspicuous, two times longer than wide, ovoid, with the inner face truncated in oblique where sensilla are placed (Figure 3b); prosternum shows dense setose punctures (n = 10) consisting of seta, each encircled by not less than five loculi (Figure 3c,d). Female length is 1.38 ± 0.17 mm (n = 10); males are 1.22 ± 0.1 mm (n = 10) in length. Male genitalia are asymmetrical, phallobase with unpaired apodeme, and sipho arcuate with spathiform siphonal proximal capsule (Figure 3e,f); the parameres are very short but recognizable and with long setae, which reach the apex of the median lobe (Figure 3g); legs with expanded femurs, middle and hind tibiae are arcuate with bristles on outer margin, and tarsus three-segmented (Figure 3h).
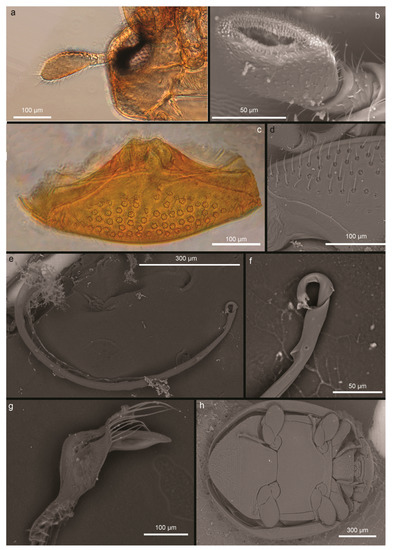
Figure 3.
Delphastus catalinae morphological characters. (a) antenna, (b) apical segment of maxillary palp, (c) prosternum, (d) particular of the setae encircled by loculi, (e) sipho, (f) tip of the sipho, (g) median lobe of parameres, and (h) body (ventral side).
Some preliminary tests conducted on leaves of C. limon infested with OSW in the laboratory confirmed D. catalinae preys on OSW. Studies are underway to examine the performance of the predator on crops in the field, its phenology, and the preferences of the hosts and plants in the new colonized environments. The activity of natural enemies will probably be exploited by employing proper conservation and augmentation techniques.
5. Conclusions
Aleurocanthus spiniferus is still spreading in the Mediterranean Basin, invading new areas and infesting new host plants. This invasive species has the potential to strongly affect the production and development of some species common in the Mediterranean orchards and gardens.
After the first record in 2008 in Southeast Italy, the alien invasive and quarantine pest A. spiniferus (Quaintance) (Hemiptera: Aleyrodidae) has gradually spread throughout Europe, infesting several new host plants in addition to the known hosts. Molecular characterization of some Italian populations and a newly found Albanian population highlighted only two different haplotypes invading Europe belonging to one of the haplogroups previously recorded in China. Through morpho-molecular characterization, the ladybird beetle D. catalinae, a Nearctic member of the tribe Serangiini, was recorded for the first time in fields in Italy in association with OSW and other whitefly populations. D. catalinae shows potential as a biocontrol agent to manage A. spiniferus outbreaks either in Italy or in other invaded countries. The finding of D. catalinae on several host plants feeding on the OSW population indicates the possibility for an eco-compatible solution to control this threatening phytophagous insect, but evaluations on its field effectiveness are still in progress.
Author Contributions
Conceptualization, F.N., F.P. and U.B.; methodology, F.N. and S.L.; software, F.N.; validation, F.N., S.L. and U.B.; formal analysis, F.N.; investigation, F.N. and S.L.; resources, U.B. and F.P.; data curation, F.N. and S.L.; writing—original draft preparation, F.N. and S.L.; writing—review and editing, F.N., U.B., S.L., F.P., G.J. and M.G.M.J.; visualization, F.N. and G.J.; supervision, U.B.; project administration, U.B. and F.P.; funding acquisition, U.B., S.L. and F.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was equally funded by the Campania Region-funded URCOFI project (Unità Regionale Coordinamento Fitosanitario) and by the Apulia Region-funded CoBiAS project (Controllo Biologico per Incremento contro Aleourocanthus spiniferus).
Acknowledgments
The authors wish to thank Anna Giulia Nappo for her technical help; Elia Russo, Flavia de Benedetta, Fortuna Miele, and Anna Bortoli for their help with the sampling and collecting activities; Giovanni Bubici for reporting of the first infestation on A. altissima; Andrea Sala and Marco Mosti for ladybeetles samples.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- EASIN—European Alien Species Information Network European Commission—Joint Research Centre 2019. Available online: https://easin.jrc.ec.europa.eu/ (accessed on 21 October 2019).
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Essl, F. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Roques, A.; Auger-Rozenberg, A.M.; Blackburn, T.M.; Garnas, J.; Pyšek, P.; Rabitsch, W.; Richardson, D.M.; Wingfield, M.J.; Liebhold, A.M.; Duncan, R.P. Temporal and interspecific variation in rates of spread for insect species invading Europe during the last 200 years. Biol. Invasions 2016, 18, 907–920. [Google Scholar] [CrossRef]
- Bernardo, U.; van Nieukerken, E.J.; Sasso, R.; Gebiola, M.; Gualtieri, L.; Viggiani, G. Characterization, distribution, biology and impact on Italian walnut orchards of the invasive North-American leafminer Coptodisca lucifluella (Lepidoptera: Heliozelidae). Bull. Entomol. Res. 2015, 105, 210–224. [Google Scholar] [CrossRef]
- Garonna, A.P.; Scarpato, S.; Vicinanza, F.; Espinosa, B. First report of Toumeyella parvicornis (Cockerell) in Europe (Hemiptera: Coccidae). Zootaxa 2015, 3949, 142–146. [Google Scholar] [CrossRef]
- Lupi, D.; Bernardo, U.; Bonsignore, C.P.; Colombo, M.; Dindo, M.L.; Faccoli, M.; Ferracini, C.; Gualtieri, L.; Marullo, R.; Mazzon, L.; et al. Insects and globalization: Sustainable control of exotic species in Italian agro-forestry ecosystems. IOBC-WPRS Bull. 2014, 100, 87–90. [Google Scholar]
- Montecchio, L.; Faccoli, M. First record of thousand cankers disease Geosmithia morbida and walnut twig beetle Pityophthorus juglandis on Juglans nigra in Europe. Plant Dis. 2014, 98, 696. [Google Scholar] [CrossRef]
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First record of an invasive fruit fly belonging to Bactrocera dorsalis complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, R.; Sato, Y.; Han, B.Y.; Huang, Z.D.; Yara, K.; Furuhashi, K. Molecular evidence for multiple phylogenetic groups within two species of invasive spiny whiteflies and their parasitoid wasp. Bull. Entomol. Res. 2016, 10, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Quaintance, A.L. New oriental Aleurodidae. Can. Entomol. 1903, 35, 61–64. [Google Scholar] [CrossRef]
- Van den Berg, M.A.; Greenland, J. Classical biological control of the spiny black fly, Aleurocanthus spiniferus (Hem.: Aleyrodidae), on citrus in Southern Africa. Entomophaga 1997, 42, 459–465. [Google Scholar] [CrossRef]
- Nguyen, R.; Sailer, R.I.; Hamon, A.B. Catalog of Aleyrodidae on Citrus and Their Natural Enemies (Homoptera—Aleyrodidae); Florida Department of Agriculture & Consumer Services, Division of Plant Industry, Bureau of Entomology: St. Gainesville, FL, USA; Centre for Agriculture and Bioscience International: Wallingford, UK, 1993; Volume 730.
- Muniappan, R.; Purea, M.; Sengebau, F.; Reddy, G.V.P. Orange spiny whitefly, Aleurocanthus spiniferus (Quaintance) (Homoptera: Aleyrodidae), and its parasitoids in the Republic of Palau. Proc. Hawaii. Entomol. Soc. 2006, 38, 21–25. [Google Scholar]
- Gillespie, P.S. A review of the whitefly genus Aleurocanthus Quaintance & Baker (Hemiptera: Aleyrodidae) in Australia. Zootaxa 2012, 3252, 1–42. [Google Scholar] [CrossRef]
- Cioffi, M.; Cornara, D.; Corrado, I.; Jansen, M.G.M.; Porcelli, F. The status of Aleurocanthus spiniferus from its unwanted introduction in Italy to date. Bull. Insectol. 2013, 66, 273–281. [Google Scholar]
- Kapantaidaki, D.E.; Antonatos, S.; Kontodimas, D.; Milonas, P.; Papachristos, D.P. Presence of the invasive whitefly Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Greece. OEPP/EPPO Bull. 2019, 49, 127–131. [Google Scholar] [CrossRef]
- Porcelli, F. First record of Aleurocanthus spiniferus (Homoptera: Aleyrodidae) in Apulia, Southern Italy. OEPP/EPPO Bull. 2008, 38, 516–518. [Google Scholar] [CrossRef]
- Nutricato, S.; D’onghia, A.M.; Fedele, A.; Filì, V.; Garonna, A.P.; Guario, A.; Guastamacchia, F.; Mazzeo, M.; Parisi, V.; Percoco, A.; et al. Nuovo aleirode da quarantena segnalato in Puglia. Inf. Agrar. 2009, 65, 57–59. [Google Scholar]
- El Kenawy, A.; Cornara, D.; Corrado, I.; El-Heneidy, A.; Rapisarda, C.; Porcelli, F. Aleurocanthus spiniferus (Quaintance) (Hemiptera Aleyrodidae) is spreading throughout the Italian region Apulia. In Proceedings of the 5th International Scientific Agricultural Symposium ‘Agrosym 2014’, Jahorina, Bosnia and Herzegovina, 23–26 October 2014. [Google Scholar] [CrossRef]
- EFSA—European Food Safety Authority. Scientific opinion on Pest categorisation of Aleurocanthus spp. EFSA J. 2018, 16, 5436. [Google Scholar]
- Šimala, M.; Masten, M.T. First record of the orange spiny whitefly, Aleurocanthus spiniferus Quaintance, 1903 (Hemiptera: Aleyrodidae), in Croatia. In Proceedings of the Conference ‘Zbornik Predavanj in Referatov, 11, Slovenskega Posvetovanja o Varstvu Rastlin Z Mednarodno Udelezbo, Bled, Slovenia, 5–6 March 2013; Available online: http://dvrs.bf.uni-lj.si/spvr/2013/60Simala.pdf (accessed on 3 August 2017).
- Radonjić, S.; Hrnčić, S.; Malumphy, C. First record of Aleurocanthus spiniferus (Quaintance) (Hemiptera Aleyrodidae) in Montenegro. Redia 2014, 77, 141–145. [Google Scholar]
- Jeffers, L. New Pest Advisory Group Report. Aleurocanthus Spiniferus Quaintance: Orange Spiny Whitefly, Hemiptera/Aleyrodidae. Available online: http://www.aphis.usda.gov/plant_health/cphst/npag/downloads/SampleReports/AleurocanthusspiniferusReport.pdf (accessed on 15 February 2018).
- Gyeltshen, J.; Hodges, A.; Hodges, G.S. Orange Spiny Whitefly, Aleurocanthus spiniferus (Quaintance) (Insecta: Hemiptera: Aleyrodidae). Available online: https://edis.ifas.ufl.edu/pdffiles/IN/IN61800.pdf (accessed on 12 August 2019).
- Zhang, Q.B. The reasons for rampage of citrus spiny white fly and its control. South China Fruits 2006, 2, 20–21. [Google Scholar]
- Clausen, C.P.; Bartlett, B.R.; Debach, P.; Goeden, R.D.; Legner, E.F.; Mcmurtry, J.A.; Oatman, E.R.; Bay, E.C.; Rosen, D. Introduced parasites and predators of arthropod pests and weeds: A world review. USDA Agric. Handb. 1978, 480, 1–545. [Google Scholar]
- Quezada, J.R. Biological control of Aleurocanthus woglumi (Homoptera: Aleyrodidae) in El Salvador. Entomophaga 1974, 19, 243–254. [Google Scholar] [CrossRef]
- Smith, H.D. Citrus blackfly on the west coast of Mexico and the importation and colonization of Eretmocerus serius Silv. for its control. Fitofolio 1945, 4, 67–103. [Google Scholar]
- Niu, J.Z.; Hull-Sanders, H.; Zhang, Y.X.; Lin, J.Z.; Dou, W.; Wang, J.J. Biological control of arthropod pests in citrus orchards in China. Biol. Control 2014, 68, 15–22. [Google Scholar] [CrossRef]
- Evans, G.A. Host Plant List of the Whiteflies (Aleyrodidae) of the World. 2007. Available online: http://entomofaune.qc.ca/entomofaune/aleurodes/references/Evans_2007_Hosts_whiteflies.pdf (accessed on 12 August 2019).
- Silvestri, F. Contribuzione alla conoscenza degli Aleurodidae (lnsecta: Hemiptera) viventi su Citrus in Estremo Oriente e dei loro parassiti. Bol. Lab. Zool. Gen. E Agric. R. Scuola Super. Agric. Portici 1928, 21, 1–60. [Google Scholar]
- Hodek, I.; Honěk, A. Ecology of Coccinellidae, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1966; p. 464. [Google Scholar]
- Mehrnejad, M.R.; Jalali, M.A. Life history parameters of the coccinellids beetle Oenopia conglobata contaminata, an important predator of the common pistachio psylla, Agonoscena pistaciae (Hemiptera: Psylloidea). Biocontrol Sci. Technol. 2004, 14, 701–711. [Google Scholar] [CrossRef]
- Han, B.Y.; Cui, L. The infection and prevalent of Aschersonia aleyrodis and Aegerita webberi against Aleurocanthus spiniferus. China Tea 2004, 2, 19. [Google Scholar]
- Zhang, Q.B.; Lei, H.D.; Li, H.J.; Liu, H.Q.; Yao, T.S.; Tian, W.H.; Qian, K.M. The damage and control of Aleurocanthus spiniferus. South China Fruits 2004, 33, 15. [Google Scholar]
- Guo, L.; Qiu, B.L.; Ren, S.X. Summary on the classify of natural enemy germplasm recourses of Aleurocanthus spiniferus (Homoptera: Aleyrodidae). Guangdong Agric. Sci. 2006, 2, 9–10. [Google Scholar]
- Campanian Executive Decree (Regione Campania) n. 2 June 22nd 2017—Misure Fitosanitarie Regionali Per il Controllo del Fitofago Aleurocanthus spiniferus. 2017. Available online: http://burc.regione.campania.it/eBurcWeb/publicContent/archivio/archivio.iface (accessed on 20 March 2019).
- Gebiola, M.; Bernardo, U.; Monti, M.M.; Navone, P.; Viggiani, G. Pnigalio agraules (Walker) and Pnigalio mediterraneus Ferrière and Delucchi (Hymenoptera: Eulophidae): Two closely related valid species. J. Nat. Hist. 2009, 43, 2465–2480. [Google Scholar] [CrossRef]
- Bink-Moenen, R.M. Revision of the African whiteflies (Aleyrodidae). Monogr. Ned. Entomol. Ver. 1983, 10, 1–211. [Google Scholar]
- Martin, J.H. An identification guide to the common whitefly pest species of the world (Homoptera, Aleyrodidae). Trop. Pest Manag. 1987, 33, 298–322. [Google Scholar] [CrossRef]
- Martin, J.H. The whitefly fauna of Australia (Sternorrhyncha: Aleyrodidae), a taxonomic account and identification guide. CSIRO Entomol. Tech. Pap. 1999, 38, 1–197. [Google Scholar]
- Kanmiya, K.; Ueda, S.; Kasai, A.; Yamashita, K.; Sato, Y.; Yoshiyasu, Y. Proposal of new specific status for tea-infesting populations of the nominal citrus spiny whitefly Aleurocanthus spiniferus (Homoptera: Aleyrodidae). Zootaxa 2011, 2797, 25–44. [Google Scholar] [CrossRef]
- Jansen, M.; Porcelli, F. Aleurocanthus camelliae (Hemiptera: Aleyrodidae), a species possibly new for the European fauna of a genus in great need of revision. Tijdschr. Entomol. 2018, 161, 63–78. [Google Scholar] [CrossRef]
- Uesugi, R.; Sato, Y. Differentiation of the tea-infesting population of citrus spiny whitefly Aleurocanthus spiniferus (Homoptera: Aleyrodidae) from the citrus-infesting population in Japan on the basis of differences in the mitochondrial cytochrome c oxidase subunit I gene. Jpn. J. Appl. Entomol. Zool. 2011, 55, 155–161. [Google Scholar] [CrossRef][Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.T.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2001, 9, 1657–1659. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Posada, D. Jmodeltest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Dlugosch, K.M.; Parker, M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, R.G.; Roderick, G.K. Evolution: Geology and climate drive diversification. Nature 2014, 509, 297–298. [Google Scholar] [CrossRef] [PubMed]
- Nugnes, F.; Gebiola, M.; Monti, M.M.; Gualtieri, L.; Giorgini, M.; Wang, J.; Bernardo, U. Genetic diversity of the invasive gall wasp Leptocybe invasa (Hymenoptera: Eulophidae) and of its Rickettsia endosymbiont, and associated sex-ratio differences. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Dittrich-Schröder, G.; Hoareau, T.B.; Hurley, B.P.; Wingfield, M.J.; Lawson, S.; Nahrung, H.F.; Slippers, B. Population genetic analyses of complex global insect invasions in managed landscapes: A Leptocybe invasa (Hymenoptera) case study. Biol. Invasions 2018, 20, 2395–2420. [Google Scholar] [CrossRef]
- Mound, L.A.; Halsey, S.H. Whitefly of the World. A Systematic Catalogue of the Aleyrodidae (Homoptera) with Host Plant and Natural Enemy Data, 1st ed.; British Museum (Natural History): London, UK, 1978; p. 340. [Google Scholar]
- Evans, G. The Whiteflies (Hemiptera: Aleyrodidae) of the World and Their Host Plants and Natural Enemies. 2007. Available online: http://keys.lucidcentral.org/keys/v3/whitefly/PDF_PwP%20ETC/world-whitefly-catalog-Evans.pdf (accessed on 12 August 2019).
- Gebiola, M.; Gómez-Zurita, J.; Monti, M.M.; Navone, P.; Bernardo, U. Integration of molecular, ecological, morphological and endosymbiont data for species delimitation within the Pnigalio soemius complex (Hymenoptera: Eulophidae). Mol. Ecol. 2012, 21, 1190–1208. [Google Scholar] [CrossRef]
- Nugnes, F.; Bernardo, U.; Viggiani, G. An integrative approach to species discrimination in the Anagrus atomus group sensu stricto (Hymenoptera: Mymaridae), with a description of a new species. Syst. Biodivers. 2017, 15, 582–599. [Google Scholar] [CrossRef]
- Bernardo, U.; Nugnes, F.; Gualtieri, L.; Nicoletti, R.; Varricchio, P.; Sasso, R.; Viggiani, G. A new gall midge species of Asphondylia (Diptera: Cecidomyiidae) inducing flower galls on Clinopodium nepeta (Lamiaceae) from Europe, its phenology, and associated fungi. Environ. Entomol. 2018, 47, 609–622. [Google Scholar] [CrossRef]
- Lawson, S.; Griffiths, M.; Nahrung, H.; Noack, A.; Wingfield, M.; Wilcken, C.; Slippers, B.; Lo, N.; Thu, P.Q.; Lee, S.; et al. Biological Control of Eucalypt Pests Overseas and in Australia; Report Australian Centre for International Agricultural Research (ACIAR); ACIAR: Canberra, Australia, 2012; pp. 1–40. Available online: http://era.daf.qld.gov.au/id/eprint/2743/1/ACIAR_Final%20Report2012_26_biological_control_of_eucalypt_pests_Lawson.pdf (accessed on 12 November 2018).
- Muirhead, J.R.; Gray, D.K.; Kelly, D.W.; Ellis, S.M.; Heath, D.D.; Macisaac, H.J. Identifying the source of species invasions: Sampling intensity vs genetic diversity. Mol. Ecol. 2008, 17, 1020–1035. [Google Scholar] [CrossRef]
- Gordon, R.D. South American Coccinellidae (Coleoptera) part III: Taxonomic revision of the Western Hemisphere genus Delphastus Casey. Frustula Entomol. 1994, 17, 71–133. [Google Scholar]
- Booth, R.G.; Polaszek, A. The identities of ladybird beetle predators used for whitefly control, with notes on some whitefly parasitoids, in Europe. In Proceedings of the Brighton Crop Protection Conference—Pests and Diseases, Brighton, UK, 18–21 November 1996; pp. 69–74. [Google Scholar]
- Hoelmer, K.A.; Osborne, L.S.; Yokomi, R.K. Reproduction and feeding behavior of Delphastus catalinae (Coleoptera: Coccinellidae), a predator of Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 1993, 86, 322–329. [Google Scholar] [CrossRef]
- Hoelmer, K.A.; Osborne, L.S.; Yokomi, R.K. Interactions of the whitefly predator Delphastus catalinae (Coleoptera: Coccinellidae) with parasitized sweetpotato whitefly (Homoptera: Aleyrodidae). Environ. Entomol. 1994, 23, 136–139. [Google Scholar] [CrossRef]
- Pickett, C.; Casanave, K.; Schoenig, S.; Heinz, K. Rearing Delphastus catalinae (Coleoptera: Coccinellidae): Practical experience and a modelling analysis. Can. Entomol. 1999, 131, 115–129. [Google Scholar] [CrossRef]
- Heinz, K.M.; Parrella, M.P. Biological control of Bemisia argentifolii (Gennadius) (Homoptera: Aleyrodidae) infesting Euphorbia pulcherrima: Evaluations of releases of Encarsia luteola (Hymenoptera: Aphelinidae) and Delphastus pusillus (Coleoptera: Coccinellidae). Environ. Entomol. 1994, 23, 1346–1353. [Google Scholar] [CrossRef][Green Version]
- Heinz, K.M.; Brazzle, J.R.; Pickett, C.; Natwick, E.T.; Nelson, J.M.; Parrella, M.P. Predatory beetle may suppress silverleaf whitefly. Calif. Agric. 1994, 48, 35–40. [Google Scholar]
- Hoelmer, K.A.; Pickett, C.H. Geographic Origin and Taxonomic History of Delphastus spp. (Coleoptera: Coccinellidae) in Commercial Culture. Biocontrol Sci. Technol. 2003, 13, 529–535. [Google Scholar] [CrossRef]
- De Almeida, R.A.; Stouthamer, R. Molecular identification of Trichogramma cacoeciae Marchal (Hymenoptera: Trichogrammatidae): A new record for Peru. Neotrop. Entomol. 2003, 32, 269–272. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).