Gamma Radiation Sterilization Dose of Adult Males in Asian Tiger Mosquito Pupae

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Source: Rearing and Preparation
2.2. Irradiation Procedure
2.3. Statistical Analysis
3. Results
3.1. Radiation Effects in the F1 Generation from Irradiated Male Pupae
3.2. Gamma Radiation Effects on Longevity in the F1 Generation from Male Pupae
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adhami, B.J.; Murati, N. Présence du moustique Aedes albopictus en Albanie. Bul. Univ. Shtet. Tiranes Ser. Shk. Mjek. 1987, 1, 13–16. [Google Scholar]
- Sprenger, D.; Wuithiranyagool, T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J. Am. Mosq. Control 1986, 2, 217–218. [Google Scholar]
- Rai, K.S. Aedes albopictus in the Americas. Ann. Rev. Entomol. 1991, 36, 459–484. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by insect vectors of human disease. Ann. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Eritja, R.; Escosa, R.; Lucientes, J.; Marquès, E.; Roiz, D.; Ruiz, S. Worldwide invasion of vector mosquitoes: Present European distribution and challenges for Spain. Biol. Invasions 2005, 7, 87–97. [Google Scholar] [CrossRef]
- Romi, R. Aedes albopictus in Italy: An underestimated health problem. Annali dell’Istituto Superiore di Sanità 2001, 37, 241–247. [Google Scholar]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Futami, K.; Valderrama, A.; Baldi, M.; Minakawa, N.; Rodríguez, R.M.; Chaves, L.F. New and common haplotypes shape genetic diversity in Asian tigermosquito populations from Costa Rica and Panamá. J. Econ. Entomol. 2015, 108, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Consoli, R.A.G.B.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1994; 228p. [Google Scholar]
- Carvalho, R.; Lourenço-de-Oliveira, R.; Braga, I. Updating the geographical distribution and frequency of Aedes albopictus in Brazil with remarks regarding its range in the Americas. Mem. Inst. Oswaldo Cruz 2014, 109, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P. Identificação de Aedes (Stegomyia) albopictus (Skuse) no Brasil. Rev. Saude Publica 1986, 20, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenço-de-Oliveira, R. High vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor of the spread of Chikungunya. J. Virol. 2014, 886, 294–6306. [Google Scholar] [CrossRef]
- Castro, M.G.; Nogueira, R.M.R.; Schatzmayr, H.G.; Miagostovich, M.P.; Lourenço-de-Oliveira, R. Dengue virus detection by using reverse transcription-polymerase chain reaction in saliva and progeny of experimentally infected Aedes albopictus from Brazil. Mem. Inst. Oswaldo Cruz 2004, 99, 809–814. [Google Scholar] [CrossRef]
- Hawley, W.A.; Reiter, P.; Copeland, R.S.; Pumpuni, C.B.; Craig, G.B., Jr. Aedes albopictus in North America: Probable introduction in used tires from northern Asia. Science 1987, 236, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xu, X.F.; Gao, N.; Fan, D.Y.; An, J.; Wang, J. Preliminary evaluation of DNA vaccine candidates encoding dengue-2 prM/E and NS1: Their immunity and protective efficacy in mice. Mol. Immunol. 2013, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Eckels, K.; Putnak, J.; Lyons, A.; Thomas, S.; Vaughn, D. Experimental dengue virus challenge of human subjects previously vaccinated with live attenuated tetravalent dengue vaccines. J. Infect. Dis. Immun. 2013, 207, 700–708. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases: Second WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Vontas, J.; Kioulos, E.; Pavlidi, N.; della Torre Morou, E.; Ranson, A.H. Insecticide resistance in the major dengue vectors Aedes albopictus and Aedes aegypti. Pestic. Biochem. Physiol. 2012, 104, 126–131. [Google Scholar] [CrossRef]
- Baldacchino, F.; Caputo, B.; Chandre, F.; Della Torre, A.; Drago Montarsi, F. Control methods against invasive Aedes mosquitoes in Europe: A review. Pest. Manag. Sci. 2015, 71, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F.; Kawashima, E.; Futami, K.; Minakawa, N.; Marín Rodríguez, R. Lack of kdr mutations in a population of Asian tiger mosquitoes from Costa Rica. Bull. Insectol. 2015, 68, 61–63. [Google Scholar]
- Valderrama, A.; Chaves, L.; Futami, K. Evaluación de mutaciones kdr en Aedes albopictus (Skuse) en Panamá (Datos preliminares). Revista Médica de Panamá 2016, 36, 30–32. [Google Scholar]
- Knipling, E.F. The Basic Principles of Insect Population Suppression and Management; U.S. Department of Agriculture: Washington, DC, USA, 1979.
- Weidhaas, D.E.; Schmidt, C.H.; Seabrook, E.L. Field studies on the release of sterile males for the control of Anopheles quadrimaculatus. Mosq. News 1962, 22, 283–291. [Google Scholar]
- Patterson, R.S.; Sharma, V.P.; Singh, K.R.P.; LaBrecque, G.C.; Seetharam, P.L.; Grover, K.K. Use of radiosterilized males to control indigenous populations of Culex pipiens quinquefasciatus Say: Laboratory and field studies. Mosq. News 1975, 35, 1–7. [Google Scholar]
- Patterson, R.S.; Lowe, R.E.; Smittle, B.J.; Dame, D.A.; Boston, M.D.; Cameron, A.L. Release of radiosterilized males to control Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 1977, 14, 299–304. [Google Scholar] [CrossRef]
- Pal, R.; LaChance, L.E. The operational feasibility of genetic methods for control of insects of medical and veterinary importance. Annu. Rev. Entomol. 1974, 19, 269–291. [Google Scholar] [CrossRef]
- Lorimer, N.; Lounibos, L.P.; Petersen, J.L. Field trials with a translocation homozygote in Aedes aegypti for population replacement. J. Econ. Entomol. 1976, 69, 405–409. [Google Scholar] [CrossRef]
- Harris, A.F.; Nimmo, D.; McKemey, A.R.; Kelly, N.; Scaife, S.; Donnelly, C.A.; Beech, C.; Petrie, W.D.; Alphey, L. Field performance of engineered male mosquitoes. Nat. Biotechnol. 2011, 29, 1034–1037. [Google Scholar] [CrossRef]
- Pan American Health Organization (PAHO). Technical Note on Transgenic Mosquitoes Engineered for Aedes Aegypti Control; PAHO: Washington, DC, USA, 2014. [Google Scholar]
- Bellini, R.; Calvitti, M.; Medici, A.; Carrieri, M.; Celli, G.; Maini, S. Use of the sterile insect technique against Aedes albopictus in Italy: First results of a pilot trial. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 505–515. [Google Scholar]
- Knipling, E.F.; Laven, H.; Craig, G.B.; Pal, R.; Kitzmiller, J.B.; Smith, C.N.; Brown, A.W. Genetic control of insects of public health importance. Bull. World Health Organ. 1968, 38, 421–438. [Google Scholar]
- World Mosquito Program. ‘Eliminate Dengue’ Program in Brazil. Available online: www.eliminatedengue.com.br (accessed on 17 March 2019).
- Bourtzis, K.; Dobson, S.L.; Xi, Z.; Rasgon, J.L.; Calvitti, M.; Moreira, L.A.; Bossin, H.C.; Moretti, R.; Baton, L.A.; Hughes, G.L.; et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. 2013, 132, S150–S163. [Google Scholar] [CrossRef]
- De Barro, P.J.; Murphy, B.; Jansen, C.C.; Murray, J. The proposed release of the yellow fever mosquito, Aedes aegypti containing a naturally occurring strain of Wolbachia pipientis, a question of regulatory responsibility. J. Verbraucherschutz Leb. 2011, 6, 33–40. [Google Scholar] [CrossRef]
- Dutra, H.L.C.; dos Santos, L.M.B.; Caragata, E.P.; Silva, J.B.L.; Villela, D.A.M.; Maciel-de-Freitas, R.; Moreira, L.A. From lab to field: The influence of urban landscapes on the invasive potential of Wolbachia in Brazilian Aedes aegypti Mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003689. [Google Scholar] [CrossRef]
- Lima, K.F.A. Avaliação Biológica de Machos Esterilizados por Radiação Gama, Para Utilização em Programas de Controle Populacional de Aedes Aegypti (Diptera: Culicidae). Ph.D. Thesis, Fundação Oswaldo Cruz, Recife, Brazil, 2015. [Google Scholar]
- Lemos, S.G. Potencial do uso de Machos Esterilizados por Radiação Gama (60Co) Para o Controle Populacional de Aedes Aegypti (Diptera-culicidae): Um Caminho Biotecnológico do Laboratório Para o Campo. Ph.D. Thesis, Universidade Federal de Pernambuco, Recife, Brasil, 2017. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Arthur, V.; Gava, M.A. Controle do Aedes; Pesquisa Gazeta de Piracicaba: Piracicaba, Brasil, 2012. [Google Scholar]
- Balestrino, F.; Medici, A.; Candini, G.; Carrieri, M.; Maccagnani, B.; Calvitti, M.; Maini, S.; Bellini, R. Gamma ray dosimetry and mating capacity studies in the laboratory on Aedes albopictus males. J. Med. Entomol. 2010, 47, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.; Jacquet, M.; Gilles, J.; Lemperiere, G.; Maquart, P.; Quilici, S.; Schooneman, F.; Vreysen, M.J.; Boyer, S. The sterile insect technique for controlling populations of Aedes albopictus (Diptera: Culicidae) on Reunion Island: Mating vigor of sterilized males. PLoS ONE 2012, 7, e49414. [Google Scholar] [CrossRef] [PubMed]
- Bellini, R.; Balestrino, F.; Medici, A.; Gentile, G.; Veronesi, R.; Carrieri, M. Mating competitiveness of Aedes albopictus radio-sterilized males in large enclosures exposed to natural conditions. J. Med. Entomol. 2013, 50, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J. Development of eggs, larvae and pupae of Aedes albopictus (Skuse) (Diptera: Culicidae). Chin. J. Entomol. Zhanghua Kunchong 1994, 14, 13–32. [Google Scholar]
- Weidhaas, C.H.; Schmidt, D.E. Mating ability of male mosquitoes, Aedes aegypti (L.) sterilized chemically or by gamma radiation. Mosq. News 1963, 23, 32–34. [Google Scholar]
- Hallinan, E.; Rai, K.S. Radiation sterilization of Aedes aegypti in nitrogen and implications for sterile male technique. Nature 1973, 244, 9–368. [Google Scholar] [CrossRef]
- Economopoulos, A.P. Sexual competitiveness of gamma-ray sterilized males of Dacus oleae. Mating frequency of artificially reared and wild females. Environ. Entomol. 1972, 1, 490–497. [Google Scholar] [CrossRef]
- Helinski, M.E.H.; Knols, B.G.J. Mating competitiveness of male Anopheles arabiensis mosquitoes irradiated with a semi- or fully-sterilizing dose in small and large laboratory cages. J. Med. Entomol. 2008, 45, 698–705. [Google Scholar] [CrossRef]
- Helinski, M.; Parker, A.; Knols, B. Radiation induced sterility for pupal and adult stages of the malaria mosquito Anopheles arabiensis. Malar. J. 2006, 15, 5–41. [Google Scholar]
- Abdel-Malek, A.A.; Tantawy, A.O.; Wakid, A.M. Studies on the eradication of Anopheles pharoensis Theobald by the sterile male technique using cobalt-60. I. Biological effect of gamma radiation on different developmental stages. J. Econ. Entomol. 1966, 59, 672–678. [Google Scholar] [CrossRef]
- Proverbs, M.D. Induced sterilization and control of insects. Annu. Rev. Entomol. 1969, 14, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Vreysen, M.J.; Gilles, J.R.; Munhenga, G.; Damiens, D.D. The effects of genetic manipulation, dieldrin treatment and irradiation on the mating competitiveness of male Anopheles arabiensis in field cages. Malar. J. 2014, 13, 318. [Google Scholar] [CrossRef] [PubMed]
- Feinendegen, L.E. Evidence for beneficial low level radiation effects and radiation hormesis. Br. J. Radiol. 2005, 78, 3–7. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
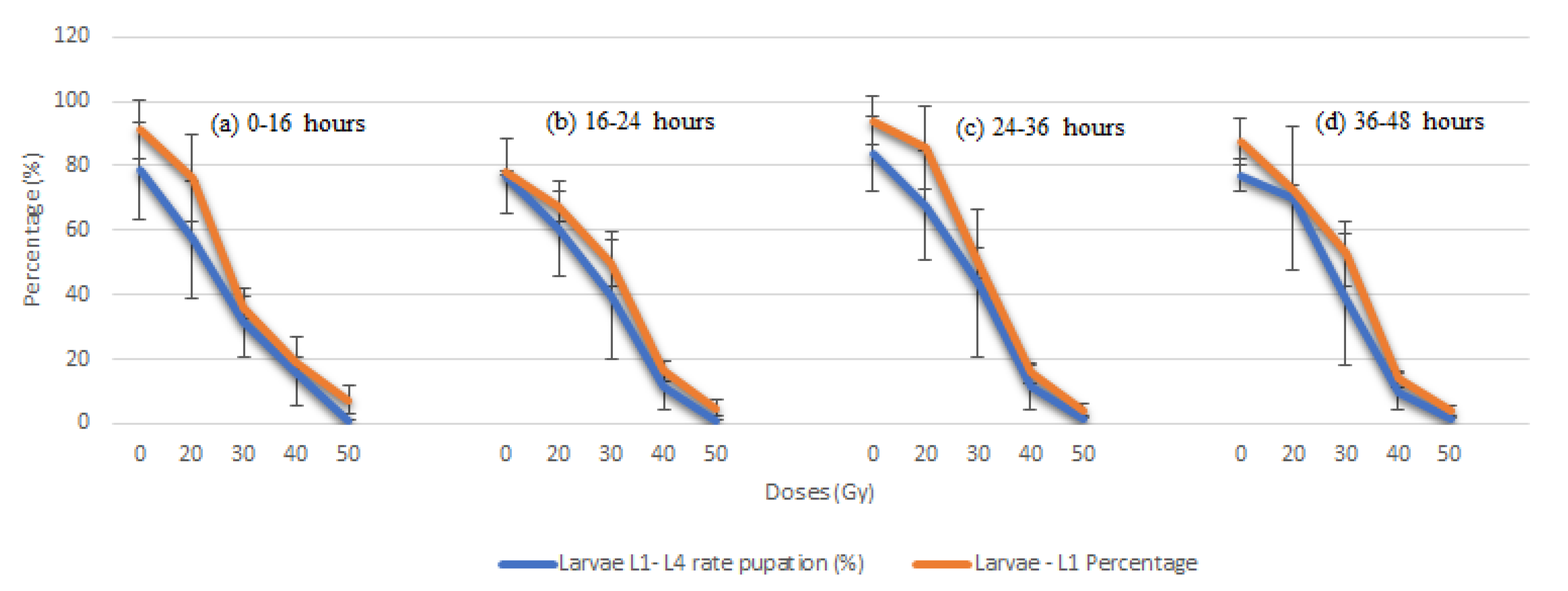
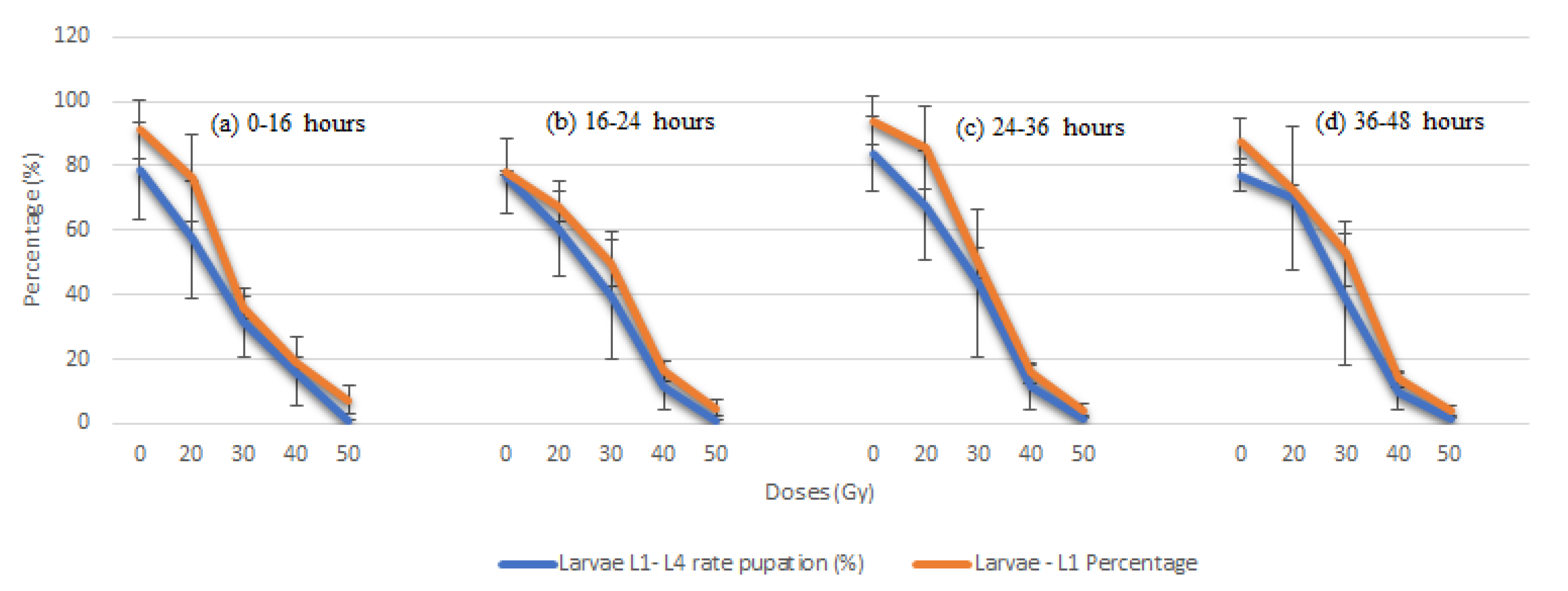
| a Age (h) | Dose Level (Gy) | Total No. Eggs | Duration of Egg Stage b (days) | No. Total Larvae L1 | Duration Larvae L1–L4 Stage | No. Total Pupae | Duration of Pupa Stage (days) | Larvae L1–L4 Rate (%) Transformed in Pupae |
|---|---|---|---|---|---|---|---|---|
| 0–16 | 0 | 2975 | 3.1 ± 1.5 | 2712 | 5.8 ± 2.1 | 2493 | 3.1 ± 2.3 | 78.4 ± 6.7 |
| 20 | 2750 | 2.9 ± 1.6 | 2099 | 5.6 ± 2.3 | 1575 | 2.8 ± 2.0 | 57.2 ± 6.0 * | |
| 30 | 2.403 | 2.7± 1.5 | 2403 | 5.6 ± 2.1 | 872 | 312 | 2.9 ± 2.1 | |
| 40 | 1.599 | 2.9 ± 1.4 | 1599 | 5.8 ± 2.3 | 302 | 49 | 3.2 ± 1.9 | |
| 50 | 1.106 | 3.3 ± 1.3 | 1106 | 5.9 ± 2.1 | 182 | 14 | 3.2 ± 1.5 | |
| 60 | 1.000 | 0.0 ± 0.0 | 1000 | 0.0 ± 0.0 | 0 | 0 | 0.0 ± 0.0 | |
| 16–24 | 0 | 3988 | 3.4 ± 2.3 | 3108 | 7.7 ± 3.4 | 2803 | 3.0 ± 2.8 | 77.0 ± 6.6 |
| 20 | 3857 | 3.2 ± 2.2 | 2594 | 6.5 ± 3.5 | 1101 | 2.7 ± 2.3 | 60.4 ± 6.1 | |
| 30 | 3203 | 2.9 ± 2.4 | 1603 | 6.2 ± 3.2 | 203 | 2.7 ± 2.0 | 39.8 ± 4.1 * | |
| 40 | 2498 | 3.6 ± 2.8 | 409 | 8.3 ± 3.3 | 34 | 3.1 ± 1.9 | 11.8 ± 2.3 * | |
| 50 | 2099 | 3.5 ± 2.6 | 100 | 8.6 ± 3.2 | 19 | 3.3 ± 2.0 | 1.1 ± 0.1 * | |
| 60 | 1902 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0.0 ± 0.0 | |
| 24–36 | 0 | 4497 | 3.3 ± 3.1 | 4229 | 7.6 ± 3.8 | 3697 | 2.9 ± 3.4 | 83.7 + 7.3 |
| 20 | 4518 | 3.3 ± 3.3 | 3876 | 6.3 ± 3.6 | 1103 | 2.6 ± 3.2 | 67.5 + 6.3 | |
| 30 | 4398 | 3.2 ± 3.4 | 2203 | 6.0 ± 3.4 | 129 | 2.8 ± 3.0 | 43.5 + 4.6 * | |
| 40 | 2998 | 3.3 ± 3.1 | 469 | 8.7 ± 3.3 | 27 | 3.2 ± 2.5 | 11.4 ± 2.9 * | |
| 50 | 2303 | 3.5 ± 3.0 | 98 | 9.0 ± 3.4 | 28 | 3.3 ± 2.4 | 1.5 ± 0.1 * | |
| 60 | 2101 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0.0 ± 0.0 | |
| 36–48 | 0 | 4336 | 3.3 ± 3.2 | 3789 | 7.7 ± 3.8 | 3687 | 2.9 ± 3.3 | 77.1 ± 6.4 |
| 20 | 5125 | 3.2 ± 3.2 | 3699 | 6.5 ± 4.1 | 1201 | 2.8 ± 3.0 | 69.9 ± 6.0 | |
| 30 | 3011 | 3.1 ± 3.0 | 1590 | 6.4 ± 3.9 | 72 | 2.8 ± 2.3 | 38.7 ± 3.9 * | |
| 40 | 2901 | 3.4 ± 3.2 | 399 | 9.1 ± 3.4 | 15 | 3.3 ± 1.2 | 9.9 ± 2.5 * | |
| 50 | 2108 | 3.6 ± 2.9 | 82 | 9.3 ± 2.9 | 16 | 3.4 ± 1.6 | 1.6 ± 0.1 * | |
| 60 | 1998 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
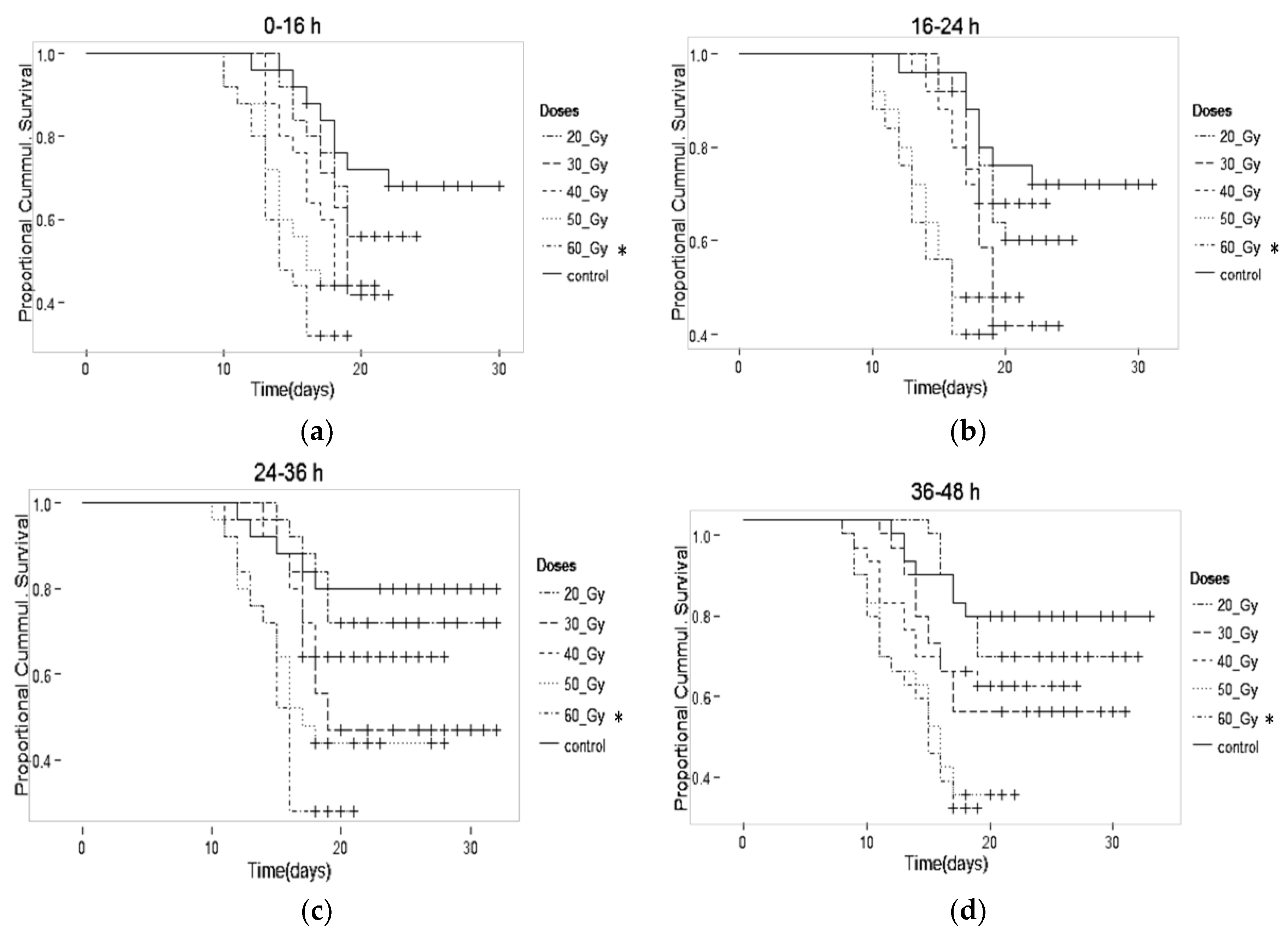
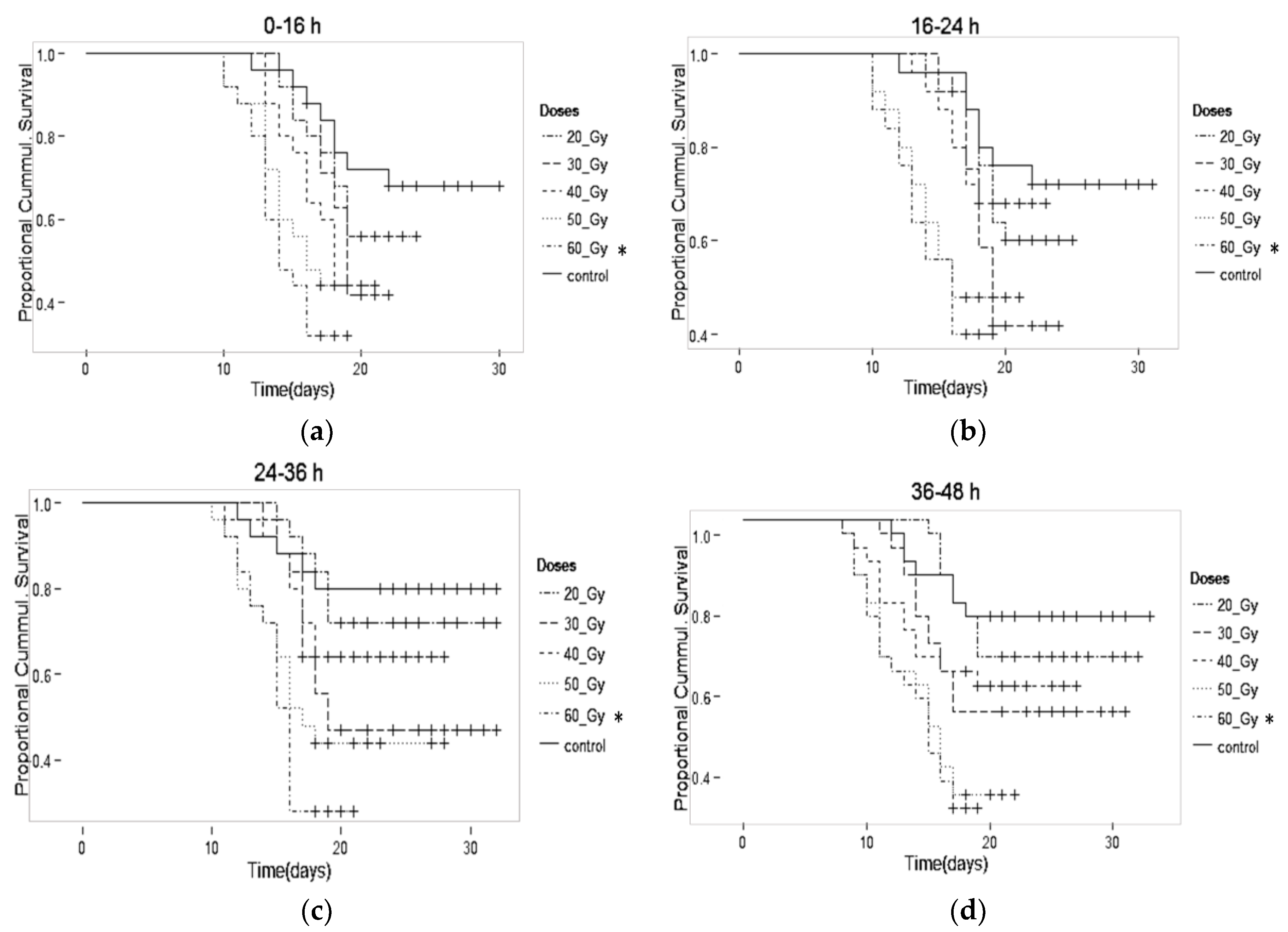
| 0–16 h | Adult Male (%) | Male Mean Survival (days) |
| Control | 75.9 ± 3.5 | 22.4 ± 2.9 |
| 20 Gy | 48.1 ± 2.9 * | 19.0 ± 2.1 |
| 30 Gy | 12.2 ± 1.8 * | 18.9 ± 2.2 |
| 40 Gy | 2.0 ± 1.0 * | 17.4 ± 2.0 * |
| 50 Gy | 0.3 ± 0.3 * | 15.6 ± 1.9 * |
| a 60 Gy | 0.0 ± 0.0 * | 14.7 ± 1.7 * |
| 16–24 h | Adult Male (%) | Male Mean Survival (days) |
| Control | 76.0 ± 3.7 | 23.2 ± 3.2 |
| 20 Gy | 50.8 ± 3.0 * | 20.2 ± 2.4 |
| 30 Gy | 22.0 ± 2.3 * | 19.2 ± 2.3 |
| 40 Gy | 0.9 ± 1.1 * | 18.8 ± 2.3 * |
| 50 Gy | 0.3 ± 0.2 * | 15.9 ± 2.1 * |
| a 60 Gy | 0.0 ± 0.0 * | 15.0 ± 2.0 * |
| 24–36 h | Adult Male (%) | Male Mean Survival (days) |
| Control | 75.8 ± 3.2 | 24.5 ± 3.3 |
| 20 Gy | 48.1 ± 2.7 * | 23.7 ± 3.2 |
| 30 Gy | 17.9 ± 2.0 * | 21.3 ± 3.1 |
| 40 Gy | 0.5 ± 0.4 * | 20.3 ± 3.1 * |
| 50 Gy | 0.2 ± 0.1 * | 17.4 ± 2.4 * |
| a 60 Gy | 0.0 ± 0.0 * | 15.6 ± 2.1 * |
| 36–48 h | Adult Male (%) | Male Mean Survival (days) |
| Control | 74.8 ± 3.0 | 22.84 ± 3.1 |
| 20 Gy | 41.0 ± 2.3 * | 22.68 ± 3.1 |
| 30 Gy | 9.9 ± 1.6 * | 19.5 ± 3.0 |
| 40 Gy | 0.3 ± 0.2 * | 17.8 ± 2.7 * |
| 50 Gy | 0.2 ± 0.1 * | 14.1 ± 2.1 * |
| a 60 Gy | 0.0 ± 0.0 * | 13.4 ± 1.8 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricardo Machi, A.; Rodrigues Mayne, R.; Adriani Gava, M.; Bergamin Arthur, P.; Arthur, V. Gamma Radiation Sterilization Dose of Adult Males in Asian Tiger Mosquito Pupae. Insects 2019, 10, 101. https://doi.org/10.3390/insects10040101
Ricardo Machi A, Rodrigues Mayne R, Adriani Gava M, Bergamin Arthur P, Arthur V. Gamma Radiation Sterilization Dose of Adult Males in Asian Tiger Mosquito Pupae. Insects. 2019; 10(4):101. https://doi.org/10.3390/insects10040101
Chicago/Turabian StyleRicardo Machi, Andre, Raquel Rodrigues Mayne, Márcio Adriani Gava, Paula Bergamin Arthur, and Valter Arthur. 2019. "Gamma Radiation Sterilization Dose of Adult Males in Asian Tiger Mosquito Pupae" Insects 10, no. 4: 101. https://doi.org/10.3390/insects10040101
APA StyleRicardo Machi, A., Rodrigues Mayne, R., Adriani Gava, M., Bergamin Arthur, P., & Arthur, V. (2019). Gamma Radiation Sterilization Dose of Adult Males in Asian Tiger Mosquito Pupae. Insects, 10(4), 101. https://doi.org/10.3390/insects10040101