Insulin Receptor Substrate Gene Knockdown Accelerates Behavioural Maturation and Shortens Lifespan in Honeybee Workers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. dsRNA Preparation
2.2. Bees
2.3. Knockdown Verification
2.4. Age of First Foraging
2.5. Lifespan and Foraging Lifespan
2.6. Statistics
3. Results
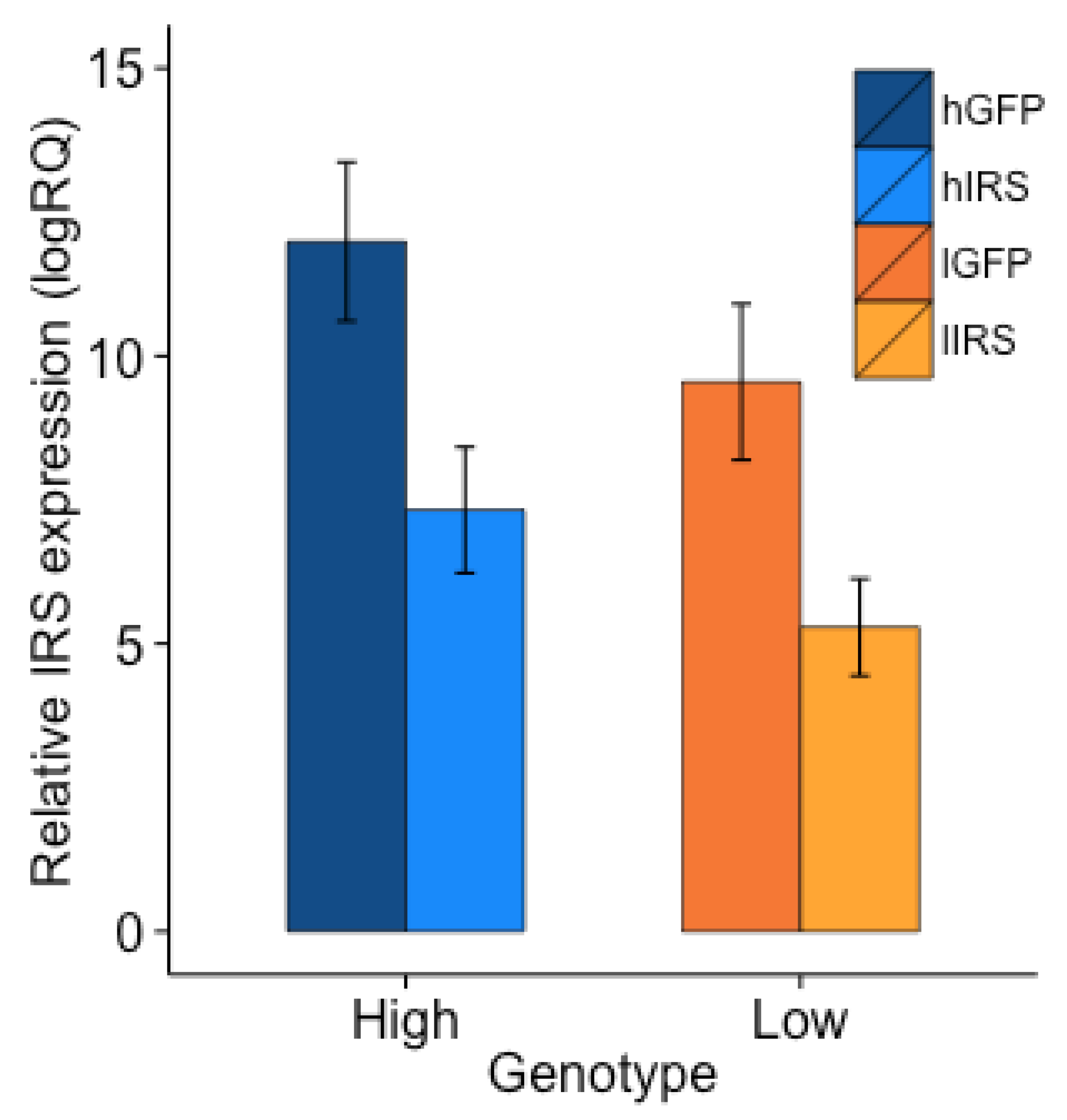
3.1. Knockdown Verification
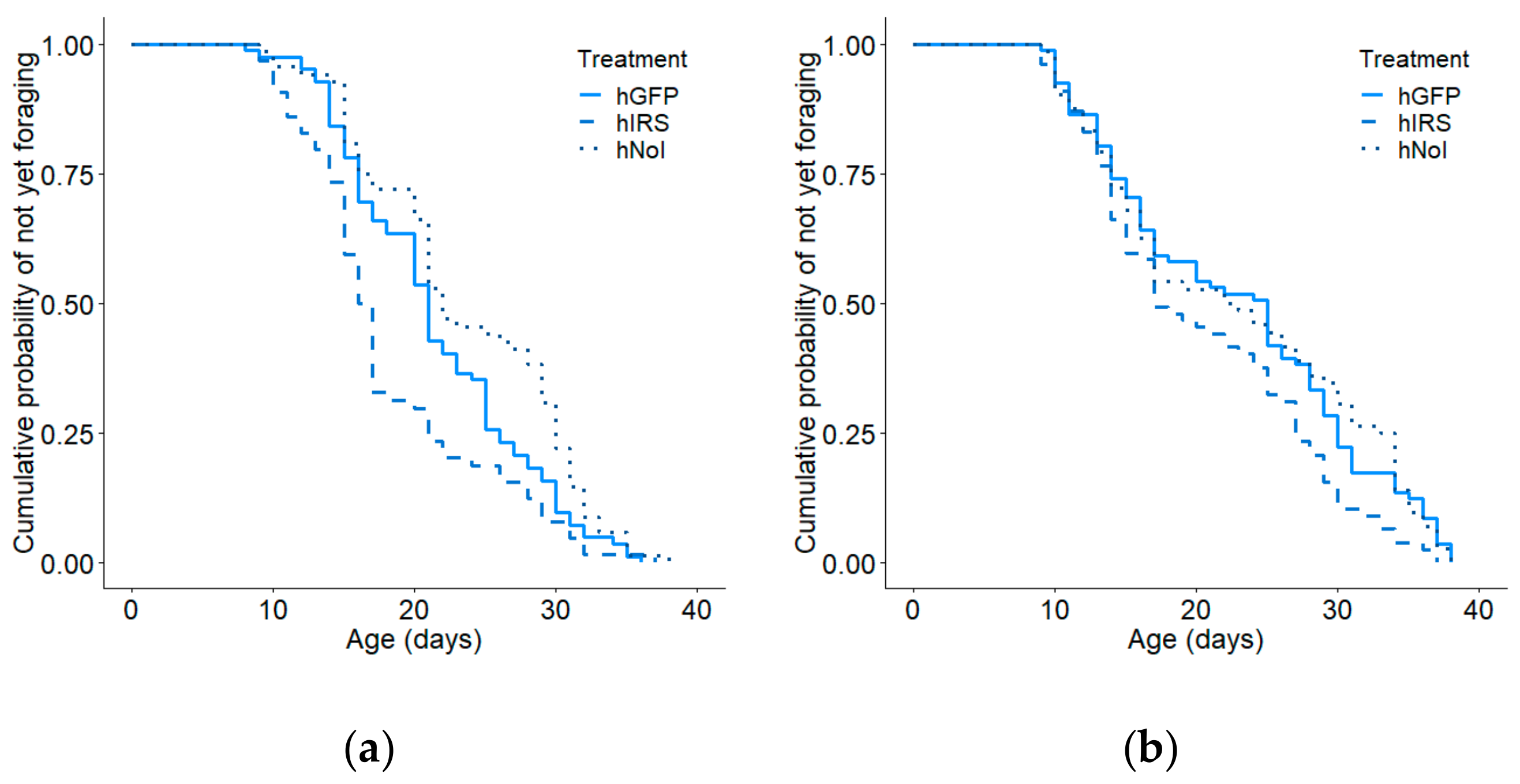
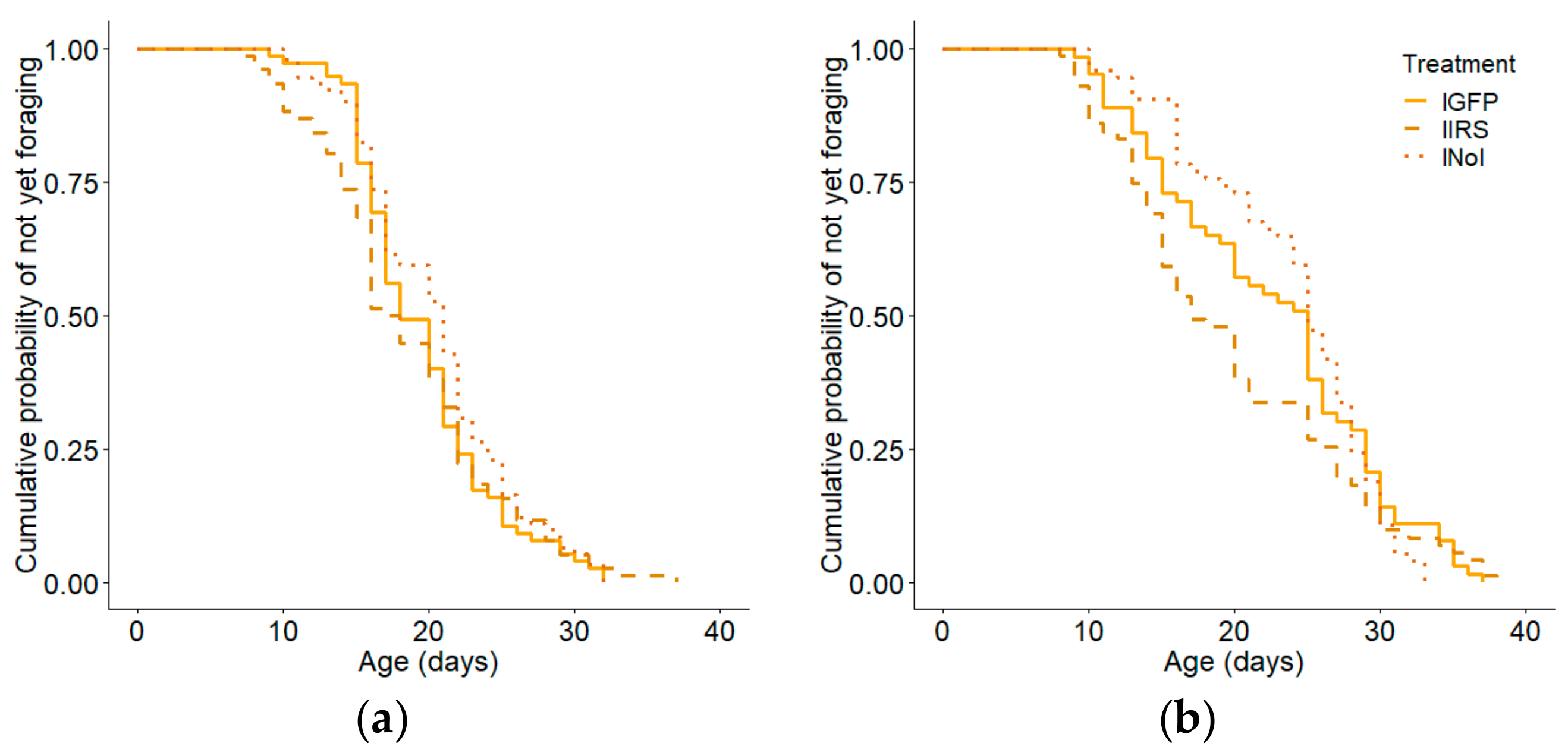
3.2. Age of First Foraging
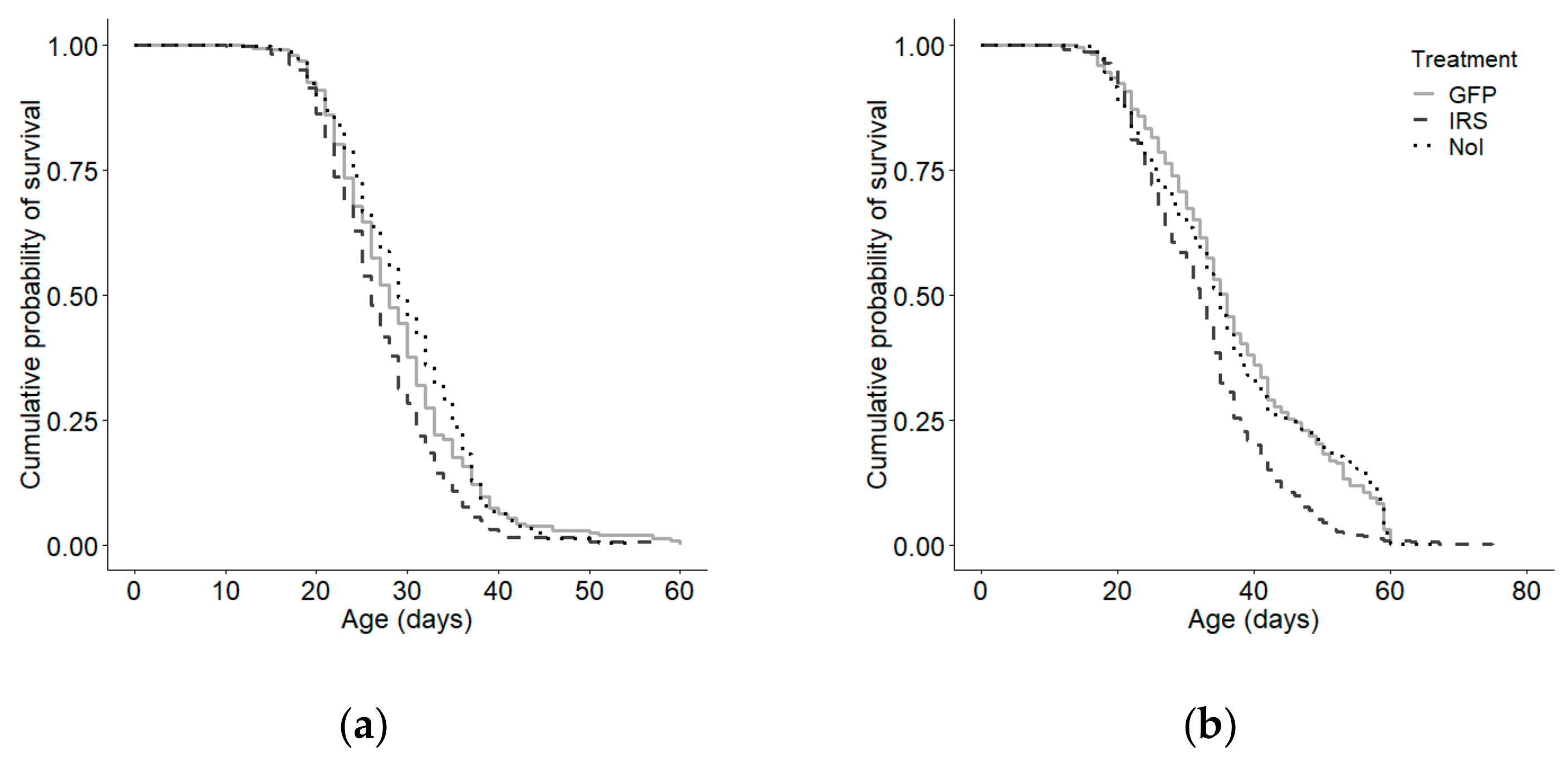
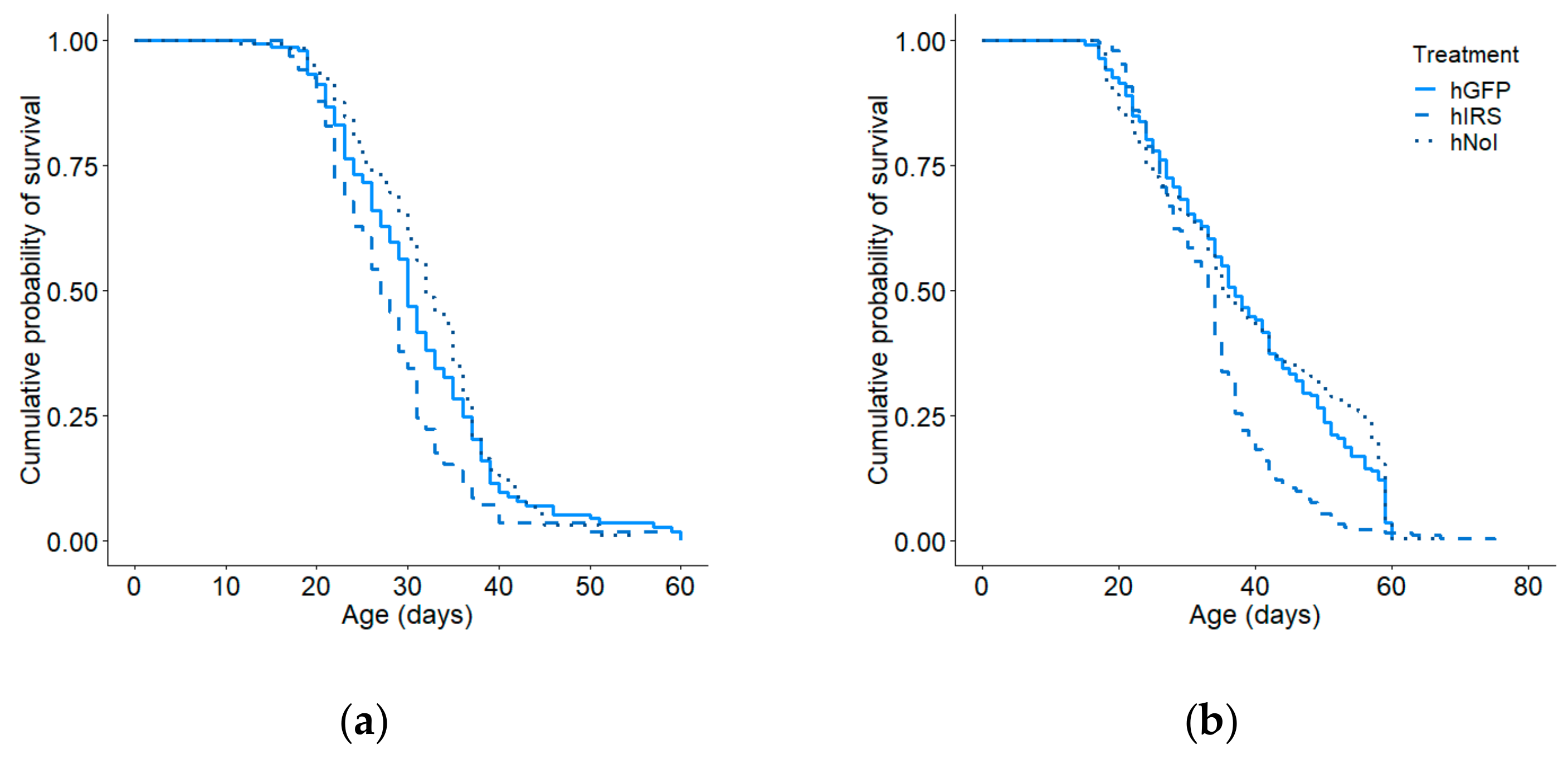
3.3. Total Lifespan and Foraging Lifespan
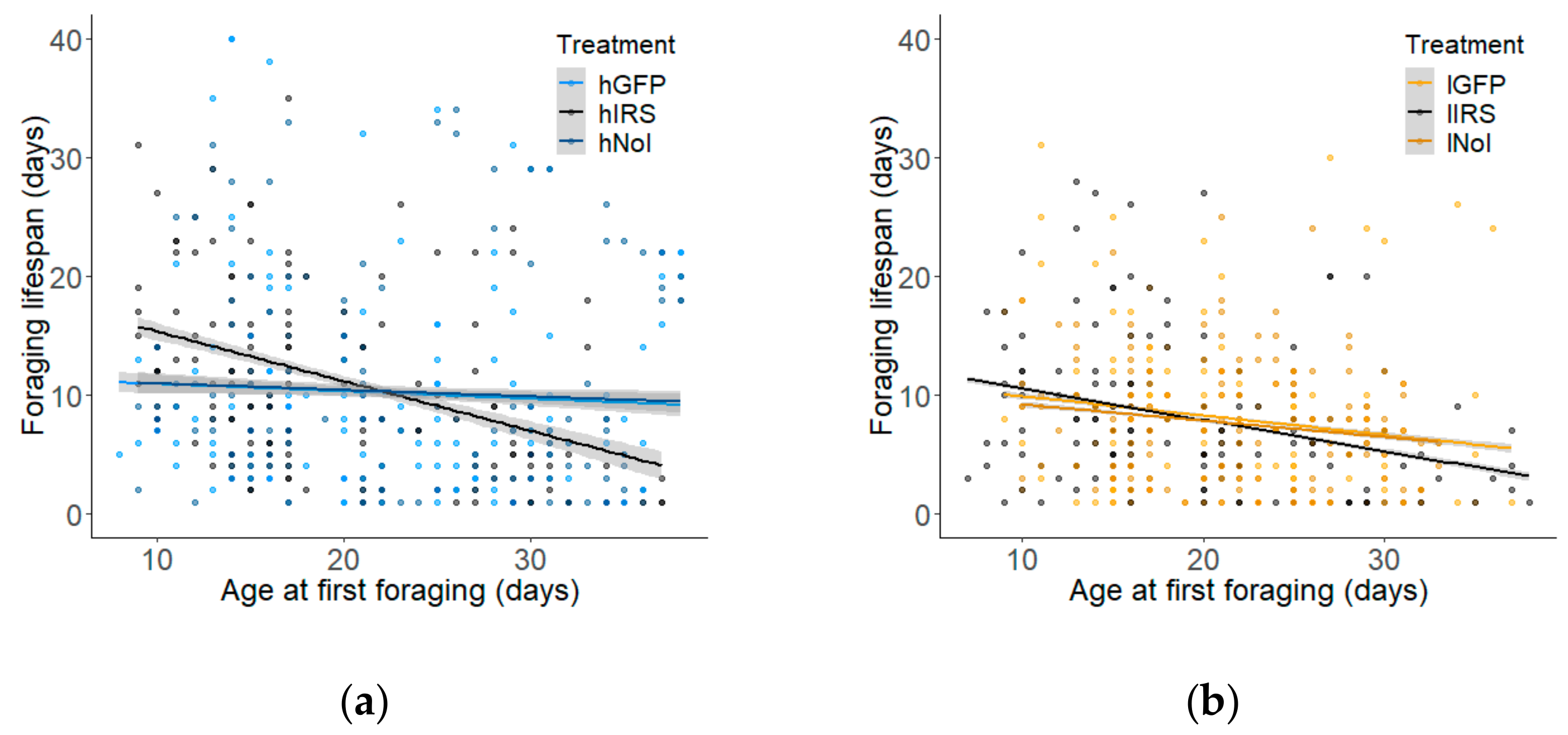
3.4. Relationship between Age of First Foraging and Lifespan Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weindruch, R.; Walford, R.L. The Retardation of Aging and Disease by Dietary Restriction; CC Thomas: Springfield, IL, USA, 1988. [Google Scholar]
- Piper, M.D.W.; Bartke, A. Diet and aging. Cell Metab. 2008, 8, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Soultoukis, G.A.; Partridge, L. Dietary Protein, Metabolism, and Aging. Annu. Rev. Biochem. 2016, 85, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Le Couteur, D.G.; Solon-Biet, S.; Cogger, V.C.; Mitchell, S.J.; Senior, A.; De Cabo, R.; Raubenheimer, D.; Simpson, S.J. The impact of low-protein high-carbohydrate diets on aging and lifespan. Cell. Mol. Life Sci. 2016, 73, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef]
- Vellai, T.; Takacs-Vellai, K.; Zhang, Y.; Kovacs, A.L.; Orosz, L.; Muller, F. Genetics: Influence of TOR kinase on lifespan in C. elegans. Nature 2003, 426, 620. [Google Scholar] [CrossRef]
- Britton, J.S.; Lockwood, W.K.; Li, L.; Cohen, S.M.; Edgar, B.A. Drosophila’s insulin/P13-kinase pathway coordinates cellular metabolism with nutritional conditions. Dev. Cell 2002, 2, 239–249. [Google Scholar] [CrossRef]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Z.W.; Shen, P. Regulation of aversion to noxious food by Drosophila neuropeptide Y–and insulin-like systems. Nat. Neurosci. 2005, 8, 1350–1355. [Google Scholar] [CrossRef]
- Wang, Y.; Mutti, N.S.; Ihle, K.E.; Siegel, A.; Dolezal, A.G.; Kaftanoglu, O.; Amdam, G.V. Down-Regulation of Honey Bee IRS Gene Biases Behavior toward Food Rich in Protein. PLoS Genet. 2010, 6, e1000896. [Google Scholar] [CrossRef]
- Semaniuk, U.V.; Gospodaryov, D.V.; Feden’ko, K.M.; Yurkevych, I.S.; Vaiserman, A.M.; Storey, K.B.; Simpson, S.J.; Lushchak, O. Insulin-Like Peptides Regulate Feeding Preference and Metabolism in Drosophila. Front. Physiol. 2018, 9, 1083. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending Healthy Life Span-From Yeast to Humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Post, S.; Kweon, Y. Nutrient control of Drosophila longevity. Trends Endocrinol. Metab. 2014, 25, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Ament, S.A.; Corona, M.; Pollock, H.S.; Robinson, G.E. Insulin signaling is involved in the regulation of worker division of labor in honey bee colonies. Proc. Natl. Acad. Sci. USA 2008, 105, 4226–4231. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.M.; Ament, S.A.; Rodriguez-Zas, S.L.; Southey, B.; Robinson, G.E. Diet and endocrine effects on behavioral maturation-related gene expression in the pars intercerebralis of the honey bee brain. J. Exp. Biol. 2015, 218, 4005–4014. [Google Scholar] [CrossRef]
- Nilsen, K.A.; Ihle, K.E.; Frederick, K.; Fondrk, M.K.; Smedal, B.; Hartfelder, K.; Amdam, G.V. Insulin-like peptide genes in honey bee fat body respond differently to manipulation of social behavioral physiology. J. Exp. Biol. 2011, 214, 1488–1497. [Google Scholar] [CrossRef]
- Page, R.E.; Rueppell, O.; Amdam, G.V. Genetics of Reproduction and Regulation of Honeybee (Apis mellifera L.) Social Behavior. Annu. Rev. Genet. 2012, 46, 97–119. [Google Scholar] [CrossRef]
- Chandra, V.; Fetter-Pruneda, I.; Oxley, P.R.; Ritger, A.L.; McKenzie, S.K.; Libbrecht, R.; Kronauer, D.J.C. Social regulation of insulin signaling and the evolution of eusociality in ants. Science 2018, 361, 398–402. [Google Scholar] [CrossRef]
- Seeley, T.D. The Wisdom of the Hive: The Social Physiology of Honey Bee Colonies; Harvard University Press: Boston, MA, USA, 1985. [Google Scholar]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef]
- Ament, S.A.; Chan, Q.W.; Wheeler, M.M.; Nixon, S.E.; Johnson, S.P.; Rodriguez-Zas, S.L.; Foster, L.J.; Robinson, G.E. Mechanisms of stable lipid loss in a social insect. J. Exp. Biol. 2011, 214, 3808–3821. [Google Scholar] [CrossRef]
- Ihle, K.E.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. Genotype effect on lifespan following vitellogenin knockdown. Exp. Gerontol. 2015, 61, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Neukirch, A. Dependence of the life-span of the honeybee (Apis mellifica) upon flight performance and energy-consumption. J. Comp. Physiol. 1982, 146, 35–40. [Google Scholar] [CrossRef]
- Amdam, G.V.; Aase, A.; Seehuus, S.C.; Fondrk, M.K.; Norberg, K.; Hartfelder, K. Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 2005, 40, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Behrends, A.; Scheiner, R.; Baker, N.; Amdam, G. Cognitive aging is linked to social role in honey bees (Apis mellifera). Exp. Gerontol. 2007, 42, 1146–1153. [Google Scholar] [CrossRef]
- Rueppell, O.; Bachelier, C.; Fondrk, M.K.; Page, R.E. Regulation of life history determines lifespan of worker honey bees (Apis mellifera L.). Exp. Gerontol. 2007, 42, 1020–1032. [Google Scholar] [CrossRef]
- Maurizio, A. The influence of pollen feeding and brood rearing on the length of life and physiological condition of the honeybee. Bee World 1950, 31, 9–12. [Google Scholar] [CrossRef]
- Altaye, S.Z.; Pirk, C.W.W.; Crewe, R.M.; Nicolson, S.W. Convergence of carbohydrate-biased intake targets in caged worker honeybees fed different protein sources. J. Exp. Biol. 2010, 213, 3311–3318. [Google Scholar] [CrossRef]
- Pirk, C.W.W.; Boodhoo, C.; Human, H.; Nicolson, S. The importance of protein type and protein to carbohydrate ratio for survival and ovarian activation of caged honeybees (Apis mellifera scutellata). Apidologie 2010, 41, 62–72. [Google Scholar] [CrossRef]
- Ihle, K.E.; Baker, N.A.; Amdam, G.V. Insulin-like peptide response to nutritional input in honey bee workers. J. Insect Physiol. 2014, 69, 49–55. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Carrillo-Tripp, J.; Miller, W.A.; Bonning, B.C.; Toth, A.L. Pollen contaminated with field-relevantlevels of Cyhalothrin affects honey bee burvival, nutritional physiology, and pollen consumption behavior. J. Econ. Entomol. 2016, 109, 41–48. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey hee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.S.; Schmidt, J.O.; Rao, H.; Wang, W.; Ligen, X. Feeding preference and survival of young worker honey bees (Hymenoptera: Apidae) fed rape, sesame, and sunflower pollen. J. Econ. Entomol. 1995, 88, 1591–1595. [Google Scholar] [CrossRef]
- Schulz, D.J.; Huang, Z.Y.; Robinson, G.E. Effects of colony food shortage on behavioral development in honey bees. Behav. Ecol. Sociobiol. 1998, 42, 295–303. [Google Scholar] [CrossRef]
- Toth, A.L.; Kantarovich, S.; Meisel, A.F.; Robinson, G.E. Nutritional status influences socially regulated foraging ontogeny in honey bees. J. Exp. Biol. 2005, 208, 4641–4649. [Google Scholar] [CrossRef]
- Toth, A.L.; Robinson, G.E. Worker nutrition and division of labour in honeybees. Anim. Behav. 2005, 69, 427–435. [Google Scholar] [CrossRef]
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Fluri, P.; Luscher, M.; Wille, H.; Gerig, L. Changes in Weight of the Pharyngeal Gland and Hemolymph Titers of Juvenile-Hormone, Protein and Vitellogenin in Worker Bees. J. Insect Physiol. 1982, 28, 61–68. [Google Scholar] [CrossRef]
- Crailsheim, K.; Schneider, L.H.W.; Hrassnigg, N.; Buhlmann, G.; Brosch, U.; Gmeinbauer, R.; Schoffmann, B. Pollen consumption and utilization in worker honeybees (Apis mellifera carnica):Dependence on individual age and function. J. Insect Physiol. 1992, 38, 409–419. [Google Scholar] [CrossRef]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, 673–677. [Google Scholar] [CrossRef]
- Crailsheim, K. Dependence of protein-metabolism on age and season in the honeybee (Apis mellifica carnica Pollm). J. Insect Physiol. 1986, 32, 629–634. [Google Scholar] [CrossRef]
- Marco Antonio, D.S.; Guidugli-Lazzarini, K.R.; Do Nascimento, A.M.; Simoes, Z.L.P.; Hartfelder, K. RNAi-mediated silencing of vitellogenin gene function turns honeybee (Apis mellifera) workers into extremely precocious foragers. Naturewissenschaften 2008, 95, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Ihle, K.E.; Page, R.E.; Frederick, K.; Fondrk, M.K.; Amdam, G.V. Genotype effect on regulation of behaviour by vitellogenin supports reproductive origin of honeybee foraging bias. Anim. Behav. 2010, 79, 1001–1006. [Google Scholar] [CrossRef] [PubMed]
- Paes de Oliveira, V.T.; Cruz-Landim, C. Morphology and function of insect fat body cells: A review. Biociências 2003, 11, 195–205. Available online: http://revistaseletronicas.pucrs.br/ojs/index.php/fabio/article/view/148 (accessed on 27 May 2015).
- Page, R.E.; Fondrk, M.K. The Effects of Colony Level Selection of the Social-Organization of Honey-Bee (Apis melifera L) Colonies: Colony Level Components of Pollen Hoarding. Behav. Ecol. Sociobiol. 1995, 36, 135–144. [Google Scholar] [CrossRef]
- Page, R.E. The Spirit of the Hive: The Mechanisms of Social Evolution; Harvard University Press: London, UK, 2013. [Google Scholar]
- Amdam, G.V.; Simoes, Z.L.; Guidugli, K.R.; Norberg, K.; Omholt, S.W. Disruption of vitellogenin gene function in adult honeybees by intra-abdominal injection of double-stranded RNA. BMC Biotechnol. 2003, 3, 1. [Google Scholar] [CrossRef]
- Amdam, G.V.; Nilsen, K.A.; Norberg, K.; Fondrk, M.K.; Hartfelder, K. Variation in endocrine signaling underlies variation in social life history. Am. Nat. 2007, 170, 37–46. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Lourenco, A.P.; Mackert, A.; Cristino, A.D.; Simoes, Z.L.P. Validation of reference genes for gene expression studies in the honey bee, Apis mellifera, by quantitative real-time RT-PCR. Apidologie 2008, 39, 372–385. [Google Scholar] [CrossRef]
- Scharlaken, B.; De Graaf, D.C.; Goossens, K.; Brunain, M.; Peelman, L.J.; Jacobs, F.J. Reference gene selection for insect expression studies using quantitative real-time PCR: The head of the honeybee, Apis mellifera, after a bacterial challenge. J. Insect Sci. 2008, 8, 33. [Google Scholar] [CrossRef]
- Wolschin, F.; Mutti, N.S.; Amdam, G.V. Insulin receptor substrate influences female caste development in honeybees. Biol. Lett. 2011, 7, 112–115. [Google Scholar] [CrossRef]
- Herb, B.R.; Wolschin, F.; Hansen, K.D.; Aryee, M.J.; Langmead, B.; Irizarry, R.; Amdam, G.V.; Feinberg, A.P. Reversible switching between epigenetic states in honeybee behavioral subcastes. Nat. Neurosci. 2012, 15, 1371–1373. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Powers, S.J. Real-Time Quantitative RT-PCR: Design, Calculations, and Statistics. Plant Cell 2009, 21, 1031–1033. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 467, 622. [Google Scholar] [CrossRef]
- Huang, Z.-Y.; Robinson, G.E. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996, 39, 147–158. [Google Scholar] [CrossRef]
- Nunes, M.F.F.; Aleixo, C.A.; Barchuk, R.A.; Bomtorin, D.A.; Grozinger, M.C.; Simões, L.P.Z. Non-Target Effects of Green Fluorescent Protein (GFP)-Derived Double-Stranded RNA (dsRNA-GFP) Used in Honey Bee RNA Interference (RNAi) Assays. Insects 2013, 4, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Rueppell, O. Characterization of Quantitative Trait Loci for the Age of First Foraging in Honey Bee Workers. Behav. Genet. 2009, 39, 541–553. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Flatt, T.; Tu, M.P.; Tatar, M. Hormonal pleiotropy and the juvenile hormone regulation of Drosophila development and life history. BioEssays 2005, 27, 999–1010. [Google Scholar] [CrossRef]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, Fat Metabolism, and Life Span: What Is the Connection? Cell Metab. 2013, 17, 10–19. [Google Scholar] [CrossRef]
- Guidugli, K.R.; Nascimento, A.M.; Amdam, G.V.; Barchuk, A.R.; Omholt, S.; Simoes, Z.L.P.; Hartfelder, K. Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Lett. 2005, 579, 4961–4965. [Google Scholar] [CrossRef]
- Pinto, L.Z.; Bitondi, M.M.G.; Simoes, Z.L.P. Inhibition of vitellogenin synthesis in Apis mellifera workers by a juvenile hormone analogue, pyriproxyfen. J. Insect Physiol. 2000, 46, 153–160. [Google Scholar] [CrossRef]
- Seehuus, S.C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Ihle, K.E.; Rueppell, O.; Huang, Z.Y.; Wang, Y.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. Genetic Architecture of a Hormonal Response to Gene Knockdown in Honey Bees. J. Hered. 2015, 106, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Novoa, E.; Page, R.E.; Gary, N.E. Behavioral and life-history components of division-of-labor in honey-bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 1994, 34, 409–417. [Google Scholar] [CrossRef]
- Visscher, P.K.; Dukas, R. Survivorship of foraging honey bees. Insectes Soc. 1997, 44, 1–5. [Google Scholar] [CrossRef]
- Page, R.E.; Fondrk, M.K.; Rueppell, O. Complex pleiotropy characterizes the pollen hoarding syndrome in honey bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2012, 66, 1459–1466. [Google Scholar] [CrossRef] [PubMed][Green Version]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ihle, K.E.; Mutti, N.S.; Kaftanoglu, O.; Amdam, G.V. Insulin Receptor Substrate Gene Knockdown Accelerates Behavioural Maturation and Shortens Lifespan in Honeybee Workers. Insects 2019, 10, 390. https://doi.org/10.3390/insects10110390
Ihle KE, Mutti NS, Kaftanoglu O, Amdam GV. Insulin Receptor Substrate Gene Knockdown Accelerates Behavioural Maturation and Shortens Lifespan in Honeybee Workers. Insects. 2019; 10(11):390. https://doi.org/10.3390/insects10110390
Chicago/Turabian StyleIhle, Kate E., Navdeep S. Mutti, Osman Kaftanoglu, and Gro V. Amdam. 2019. "Insulin Receptor Substrate Gene Knockdown Accelerates Behavioural Maturation and Shortens Lifespan in Honeybee Workers" Insects 10, no. 11: 390. https://doi.org/10.3390/insects10110390
APA StyleIhle, K. E., Mutti, N. S., Kaftanoglu, O., & Amdam, G. V. (2019). Insulin Receptor Substrate Gene Knockdown Accelerates Behavioural Maturation and Shortens Lifespan in Honeybee Workers. Insects, 10(11), 390. https://doi.org/10.3390/insects10110390