
1,8-Cineol Attenuates Checkpoint Molecule PDL-1 and Adhesion Molecule CX3CR1 in Circulating Monocytes in Otitis Media Patients
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Characteristics of Examined Patients
2.2. Blood Collection and FACS Analysis of Monocyte Subsets in Whole Blood
2.3. THP-1 Cells and Culture Conditions
2.4. Cytokine Analyses
2.5. Statistical Analyses
3. Results
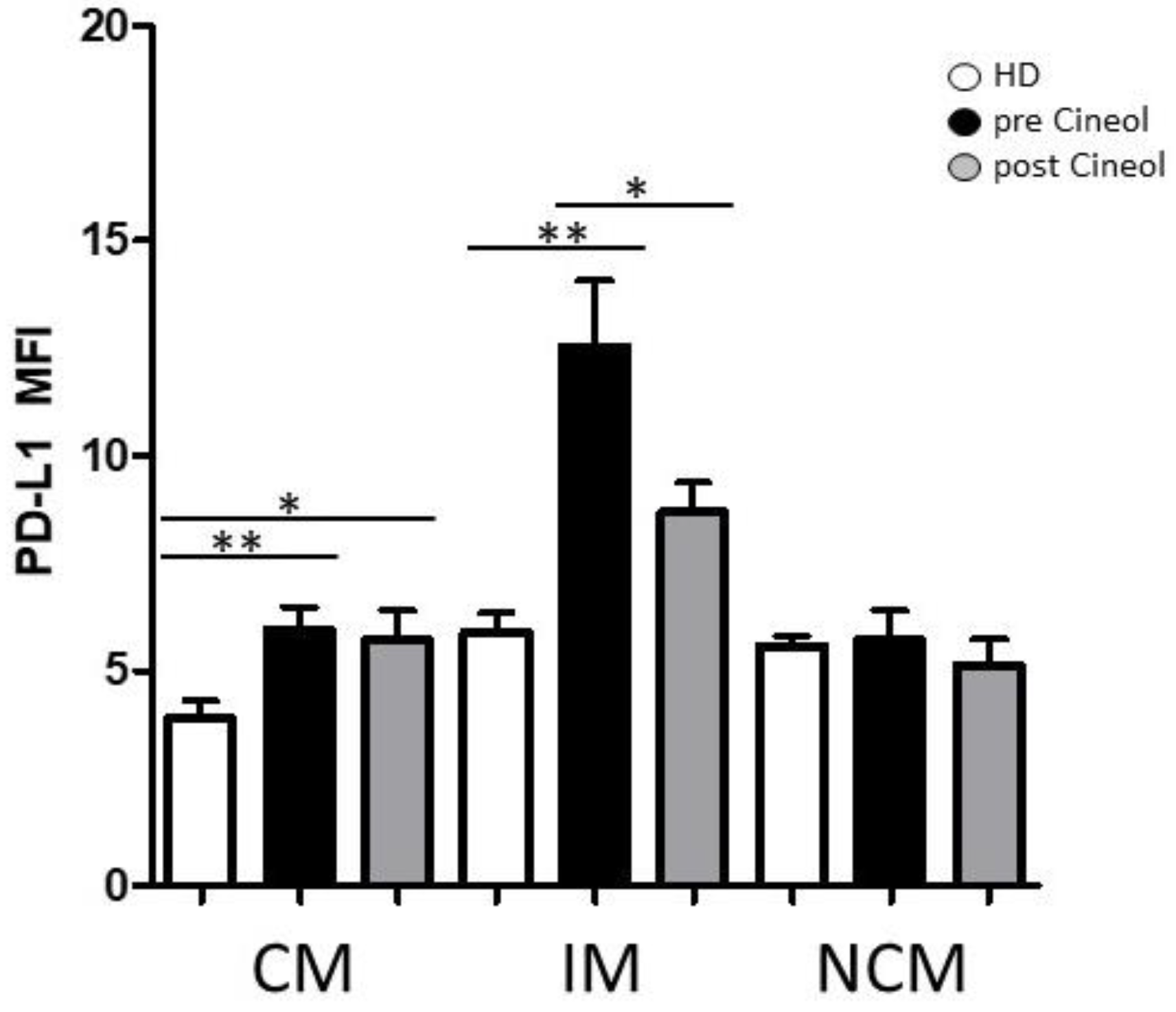
3.1. Monocyte Subset Characteristics upon 1,8-Cineol Tretament of Otitis Media Patients
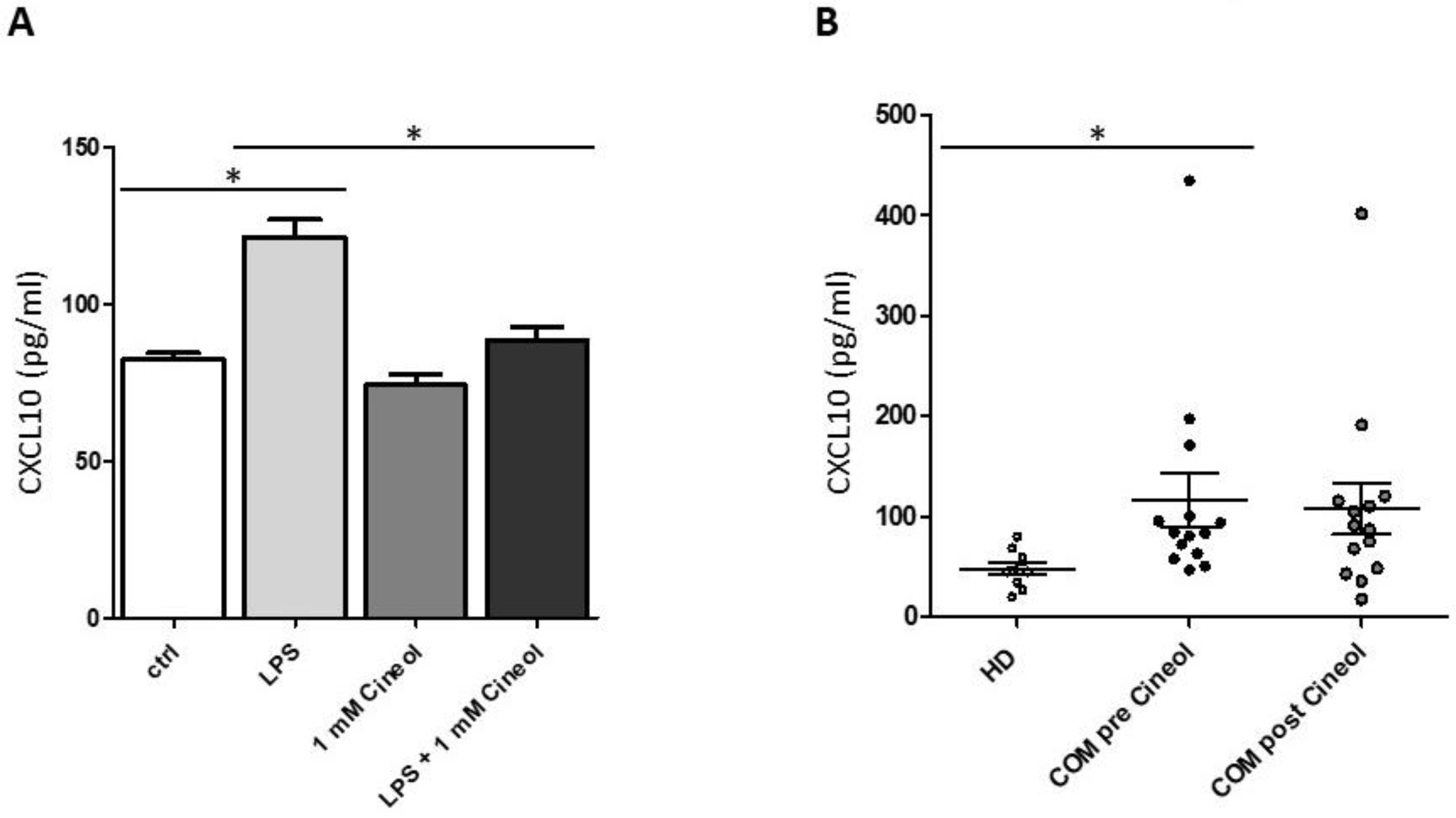
3.2. Treatment of THP-1 Monocytes with Lipopolysaccharide and 1,8-Cineol
4. Discussion
4.1. Alteration of Monocytic PD-L1 and CX3CR1
4.2. Impact of 1,8-Cineol on LPS-Induced Monocytic Cytokine Secretion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kapellos, T.S.; Bonaguro, L.; Gemund, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef] [PubMed]
- Leichtle, A.; Hernandez, M.; Ebmeyer, J.; Yamasaki, K.; Lai, Y.; Radek, K.; Choung, Y.H.; Euteneuer, S.; Pak, K.; Gallo, R.; et al. CC chemokine ligand 3 overcomes the bacteriocidal and phagocytic defect of macrophages and hastens recovery from experimental otitis media in TNF-/- mice. J. Immunol. 2010, 184, 3087–3097. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Thai, A.; Cao, Z.; Chen, X.; Chen, J.; Bacacao, B.; Bekale, L.A.; Schiel, V.; Bollyky, P.L.; Maria, P.L.S. Chronic suppurative otitis media causes macrophage-associated sensorineural hearing loss. J. Neuroinflamm. 2022, 19, 224. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sun, J. Intracranial complications of chronic otitis media. Eur. Arch. Oto-Rhino-Laryngol. Off. J. Eur. Fed. Oto-Rhino-Laryngol. Soc. 2014, 271, 2923–2926. [Google Scholar] [CrossRef] [PubMed]
- Shi, X. Pathophysiology of the cochlear intrastrial fluid-blood barrier (review). Hear. Res. 2016, 338, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Leichtle, A.; Kurabi, A.; Leffers, D.; Darr, M.; Draf, C.S.; Ryan, A.F.; Bruchhage, K.L. Immunomodulation as a Protective Strategy in Chronic Otitis Media. Front. Cell. Infect. Microbiol. 2022, 12, 826192. [Google Scholar] [CrossRef] [PubMed]
- Monasta, L.; Ronfani, L.; Marchetti, F.; Montico, M.; Vecchi Brumatti, L.; Bavcar, A.; Grasso, D.; Barbiero, C.; Tamburlini, G. Burden of disease caused by otitis media: Systematic review and global estimates. PLoS ONE 2012, 7, e36226. [Google Scholar] [CrossRef]
- Bruchhage, K.L.; Lupatsii, M.; Mollenkolk, F.; Leffers, D.; Kurabi, A.; Jurgens, T.; Graspeuntner, S.; Hollfelder, D.; Leichtle, A. Hearing rehabilitation and microbial shift after middle ear surgery with Vibrant Soundbridge in patients with chronic otitis media. Eur. Arch. Oto-Rhino-Laryngol. Off. J. Eur. Fed. Oto-Rhino-Laryngol. Soc. 2023, 280, 3107–3118. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.; Wallis, S.; Coatesworth, A.P. Acute otitis media. Postgrad. Med. 2015, 127, 386–390. [Google Scholar] [CrossRef]
- Bhutta, M.F.; Lambie, J.; Hobson, L.; Williams, D.; Tyrer, H.E.; Nicholson, G.; Brown, S.D.M.; Brown, H.; Piccinelli, C.; Devailly, G.; et al. Transcript Analysis Reveals a Hypoxic Inflammatory Environment in Human Chronic Otitis Media With Effusion. Front. Genet. 2019, 10, 1327. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Lisi, C.V.; Gerring, R.; Mittal, J.; Mathee, K.; Narasimhan, G.; Azad, R.K.; Yao, Q.; Grati, M.; Yan, D.; et al. Current concepts in the pathogenesis and treatment of chronic suppurative otitis media. J. Med. Microbiol. 2015, 64, 1103–1116. [Google Scholar] [CrossRef]
- Guo, H.; Li, M.; Xu, L.J. Apigetrin treatment attenuates LPS-induced acute otitis media though suppressing inflammation and oxidative stress. Biomed. Pharmacother. 2019, 109, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Heitbrock, L. Blood Monocytes and Their Subsets: Established Features and Open Questions. Front. Immunol. 2015, 6, 423. [Google Scholar] [CrossRef]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef]
- Medrano-Bosch, M.; Simon-Codina, B.; Jimenez, W.; Edelman, E.R.; Melgar-Lesmes, P. Monocyte-endothelial cell interactions in vascular and tissue remodeling. Front. Immunol. 2023, 14, 1196033. [Google Scholar] [CrossRef]
- Pries, R.; Jeschke, S.; Leichtle, A.; Bruchhage, K.L. Modes of Action of 1,8-Cineol in Infections and Inflammation. Metabolites 2023, 13, 751. [Google Scholar] [CrossRef]
- Juergens, L.J.; Worth, H.; Juergens, U.R. New Perspectives for Mucolytic, Anti-inflammatory and Adjunctive Therapy with 1,8-Cineole in COPD and Asthma: Review on the New Therapeutic Approach. Adv. Ther. 2020, 37, 1737–1753. [Google Scholar] [CrossRef] [PubMed]
- Simsek, M.; Duman, R. Investigation of Effect of 1,8-cineole on Antimicrobial Activity of Chlorhexidine Gluconate. Pharmacogn. Res. 2017, 9, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Djenane, D.; Yanguela, J.; Amrouche, T.; Boubrit, S.; Boussad, N.; Roncales, P. Chemical composition and antimicrobial effects of essential oils of Eucalyptus globulus, Myrtus communis and Satureja hortensis against Escherichia coli O157:H7 and Staphylococcus aureus in minced beef. Food Sci. Technol. Int. Cienc. Y Tecnol. Los. Aliment. Int. 2011, 17, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Ocana, A.; Reglero, G. Effects of Thyme Extract Oils (from Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on Cytokine Production and Gene Expression of oxLDL-Stimulated THP-1-Macrophages. J. Obes. 2012, 2012, 104706. [Google Scholar] [CrossRef]
- Bruchhage, K.L.; Koennecke, M.; Drenckhan, M.; Plotze-Martin, K.; Pries, R.; Wollenberg, B. 1,8-cineol inhibits the Wnt/beta-catenin signaling pathway through GSK-3 dephosphorylation in nasal polyps of chronic rhinosinusitis patients. Eur. J. Pharmacol. 2018, 835, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Greiner, J.F.; Muller, J.; Zeuner, M.T.; Hauser, S.; Seidel, T.; Klenke, C.; Grunwald, L.M.; Schomann, T.; Widera, D.; Sudhoff, H.; et al. 1,8-Cineol inhibits nuclear translocation of NF-kappaB p65 and NF-kappaB-dependent transcriptional activity. Biochim. Biophys. Acta 2013, 1833, 2866–2878. [Google Scholar] [CrossRef] [PubMed]
- Kehrl, W.; Sonnemann, U.; Dethlefsen, U. Therapy for acute nonpurulent rhinosinusitis with cineole: Results of a double-blind, randomized, placebo-controlled trial. Laryngoscope 2004, 114, 738–742. [Google Scholar] [CrossRef]
- Worth, H.; Schacher, C.; Dethlefsen, U. Concomitant therapy with Cineole (Eucalyptole) reduces exacerbations in COPD: A placebo-controlled double-blind trial. Respir. Res. 2009, 10, 69. [Google Scholar] [CrossRef]
- Worth, H.; Dethlefsen, U. Patients with asthma benefit from concomitant therapy with cineole: A placebo-controlled, double-blind trial. J. Asthma Off. J. Assoc. Care Asthma 2012, 49, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Polasky, C.; Steffen, A.; Loyal, K.; Lange, C.; Bruchhage, K.; Pries, R. Redistribution of Monocyte Subsets in Obstructive Sleep Apnea Syndrome Patients Leads to an Imbalanced PD-1/PD-L1 Cross-Talk with CD4/CD8 T Cells. J. Immunol. 2021, 206, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Schittenhelm, L.; Hilkens, C.M.; Morrison, V.L. beta(2) Integrins As Regulators of Dendritic Cell, Monocyte, and Macrophage Function. Front. Immunol. 2017, 8, 1866. [Google Scholar] [CrossRef] [PubMed]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007, 317, 666–670. [Google Scholar] [CrossRef]
- Wu, S.P.; Liao, R.Q.; Tu, H.Y.; Wang, W.J.; Dong, Z.Y.; Huang, S.M.; Guo, W.B.; Gou, L.Y.; Sun, H.W.; Zhang, Q.; et al. Stromal PD-L1-Positive Regulatory T cells and PD-1-Positive CD8-Positive T cells Define the Response of Different Subsets of Non-Small Cell Lung Cancer to PD-1/PD-L1 Blockade Immunotherapy. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2018, 13, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Carlsson, R.; Comabella, M.; Wang, J.; Kosicki, M.; Carrion, B.; Hasan, M.; Wu, X.; Montalban, X.; Dziegiel, M.H.; et al. FoxA1 directs the lineage and immunosuppressive properties of a novel regulatory T cell population in EAE and MS. Nat. Med. 2014, 20, 272–282. [Google Scholar] [CrossRef] [PubMed]
- McKendry, R.T.; Spalluto, C.M.; Burke, H.; Nicholas, B.; Cellura, D.; Al-Shamkhani, A.; Staples, K.J.; Wilkinson, T.M. Dysregulation of Antiviral Function of CD8(+) T Cells in the Chronic Obstructive Pulmonary Disease Lung. Role of the PD-1-PD-L1 Axis. Am. J. Respir. Crit. Care Med. 2016, 193, 642–651. [Google Scholar] [CrossRef]
- Rutigliano, J.A.; Sharma, S.; Morris, M.Y.; Oguin, T.H.; McClaren, J.L.; Doherty, P.C.; Thomas, P.G. Highly Pathological Influenza A Virus Infection Is Associated with Augmented Expression of PD-1 by Functionally Compromised Virus-Specific CD8(+) T Cells. J. Virol. 2014, 88, 1636–1651. [Google Scholar] [CrossRef]
- Goto, S.; Konnai, S.; Okagawa, T.; Nishimori, A.; Maekawa, N.; Gondaira, S.; Higuchi, H.; Koiwa, M.; Tajima, M.; Kohara, J.; et al. Increase of cells expressing PD-1 and PD-L1 and enhancement of IFN-gamma production via PD-1/PD-L1 blockade in bovine mycoplasmosis. Immun. Inflamm. Dis. 2017, 5, 355–363. [Google Scholar] [CrossRef]
- Wallis, S.; Atkinson, H.; Coatesworth, A.P. Chronic otitis media. Postgrad. Med. 2015, 127, 391–395. [Google Scholar] [CrossRef]
- Leffers, D.; Penxova, Z.; Kempin, T.; Darr, M.; Fleckner, J.; Hollfelder, D.; Ryan, A.F.; Bruchhage, K.L.; Kurabi, A.; Leichtle, A. Immunomodulatory Response of the Middle Ear Epithelial Cells in Otitis Media. Otol. Neurotol. Off. Public Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2024, 5, e248–e255. [Google Scholar] [CrossRef] [PubMed]
- Skovbjerg, S.; Roos, K.; Andersson, M.; Rabe, H.; Nilsson, S.; Lindh, M.; Wold, A.E. Inflammatory Mediator Profiles in Secretory Otitis Media in Relationship to Viable Bacterial Pathogens and Bacterial and Viral Nucleic Acids. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2020, 40, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Filatov, V.A.; Kulyak, O.Y.; Kalenikova, E.I. Chemical Composition and Antimicrobial Potential of a Plant-Based Substance for the Treatment of Seborrheic Dermatitis. Pharmaceuticals 2023, 16, 328. [Google Scholar] [CrossRef]
- Tahiri, N.E.H.; Saghrouchni, H.; Hamamouch, N.; Khomsi, M.E.; Alzahrani, A.; Salamatullah, A.M.; Badiaa, L.; Lrhorfi, L.A. Treatment with Glyphosate Induces Tolerance of Citrus Pathogens to Glyphosate and Fungicides but Not to 1,8-Cineole. Molecules 2022, 27, 8300. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Wang, X.F.; Tang, F.D.; Zhou, J.Y.; Lu, G.H.; Wang, Y.; Bian, R.L. Inhibitory effect of 1,8-cineol (eucalyptol) on Egr-1 expression in lipopolysaccharide-stimulated THP-1 cells. Acta Pharmacol. Sin. 2007, 28, 908–912. [Google Scholar] [CrossRef]
- Yadav, N.; Chandra, H. Suppression of inflammatory and infection responses in lung macrophages by eucalyptus oil and its constituent 1,8-cineole: Role of pattern recognition receptors TREM-1 and NLRP3, the MAP kinase regulator MKP-1, and NFkappaB. PLoS ONE 2017, 12, e0188232. [Google Scholar] [CrossRef] [PubMed]
- Asensio, V.C.; Maier, J.; Milner, R.; Boztug, K.; Kincaid, C.; Moulard, M.; Phillipson, C.; Lindsley, K.; Krucker, T.; Fox, H.S.; et al. Interferon-independent, human immunodeficiency virus type 1 gp120-mediated induction of CXCL10/IP-10 gene expression by astrocytes in vivo and in vitro. J. Virol. 2001, 75, 7067–7077. [Google Scholar] [CrossRef] [PubMed]
- Petrovic-Djergovic, D.; Popovic, M.; Chittiprol, S.; Cortado, H.; Ransom, R.F.; Partida-Sanchez, S. CXCL10 induces the recruitment of monocyte-derived macrophages into kidney, which aggravate puromycin aminonucleoside nephrosis. Clin. Exp. Immunol. 2015, 180, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.F.; Kim, D.H.; Yoon, Y.S.; Jin, D.; Huang, X.Z.; Li, J.H.; Deung, Y.K.; Lee, K.J. Essential involvement of cross-talk between IFN-gamma and TNF-alpha in CXCL10 production in human THP-1 monocytes. J. Cell. Physiol. 2009, 220, 690–697. [Google Scholar] [CrossRef]
- Zhao, Q.; Kim, T.; Pang, J.; Sun, W.; Yang, X.; Wang, J.; Song, Y.; Zhang, H.; Sun, H.; Rangan, V.; et al. A novel function of CXCL10 in mediating monocyte production of proinflammatory cytokines. J. Leukoc. Biol. 2017, 102, 1271–1280. [Google Scholar] [CrossRef]
- Ioannidis, L.J.; Eriksson, E.; Hansen, D.S. CD14(+) monocytes are the main leucocytic sources of CXCL10 in response to Plasmodium falciparum. Parasitology 2020, 147, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Inchaustegui, D.A.; Hogg, A.E.; Tulliano, G.; Llanos-Cuentas, A.; Arevalo, J.; Endsley, J.J.; Soong, L. CXCL10 production by human monocytes in response to Leishmania braziliensis infection. Infect. Immun. 2010, 78, 301–308. [Google Scholar] [CrossRef]
- DeMaria, T.F.; McGhee, R.B., Jr.; Lim, D.J. Rheumatoid factor in otitis media with effusion. Arch. Otolaryngol. 1984, 110, 279–280. [Google Scholar] [CrossRef] [PubMed]
- van Hooij, A.; Boeters, D.M.; Tjon Kon Fat, E.M.; van den Eeden, S.J.F.; Corstjens, P.; van der Helm-van Mil, A.H.M.; Geluk, A. Longitudinal IP-10 Serum Levels Are Associated with the Course of Disease Activity and Remission in Patients with Rheumatoid Arthritis. Clin. Vaccine Immunol. CVI 2017, 24, e00060-17. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.S.; Silva, M.F.S.; Nunes, X.P.; Colombo, A.V.; Oliveira, H.P.; Goto, P.L.; Blanzat, M.; Piva, H.L.; Tedesco, A.C.; Siqueira-Moura, M.P. Cineole-containing nanoemulsion: Development, stability, and antibacterial activity. Chem. Phys. Lipids 2021, 239, 105113. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.P.; Ennen, I.; Greiner, J.F.W.; Dietsch, R.; Patel, A.; Kaltschmidt, B.; Kaltschmidt, C.; Hutten, A. Preparation of Terpenoid-Invasomes with Selective Activity against S. aureus and Characterization by Cryo Transmission Electron Microscopy. Biomedicines 2020, 8, 105. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leichtle, A.; Jeschke, S.; Plötze-Martin, K.; Idel, C.; Bruchhage, K.-L.; Pries, R. 1,8-Cineol Attenuates Checkpoint Molecule PDL-1 and Adhesion Molecule CX3CR1 in Circulating Monocytes in Otitis Media Patients. J. Pers. Med. 2024, 14, 279. https://doi.org/10.3390/jpm14030279
Leichtle A, Jeschke S, Plötze-Martin K, Idel C, Bruchhage K-L, Pries R. 1,8-Cineol Attenuates Checkpoint Molecule PDL-1 and Adhesion Molecule CX3CR1 in Circulating Monocytes in Otitis Media Patients. Journal of Personalized Medicine. 2024; 14(3):279. https://doi.org/10.3390/jpm14030279
Chicago/Turabian StyleLeichtle, Anke, Stephanie Jeschke, Kirstin Plötze-Martin, Christian Idel, Karl-Ludwig Bruchhage, and Ralph Pries. 2024. "1,8-Cineol Attenuates Checkpoint Molecule PDL-1 and Adhesion Molecule CX3CR1 in Circulating Monocytes in Otitis Media Patients" Journal of Personalized Medicine 14, no. 3: 279. https://doi.org/10.3390/jpm14030279
APA StyleLeichtle, A., Jeschke, S., Plötze-Martin, K., Idel, C., Bruchhage, K.-L., & Pries, R. (2024). 1,8-Cineol Attenuates Checkpoint Molecule PDL-1 and Adhesion Molecule CX3CR1 in Circulating Monocytes in Otitis Media Patients. Journal of Personalized Medicine, 14(3), 279. https://doi.org/10.3390/jpm14030279