
In Vitro Models for Improved Therapeutic Interventions in Atrial Fibrillation

 , , and
, , and
Abstract
1. Introduction
1.1. Current Therapeutic Options for Atrial Fibrillation
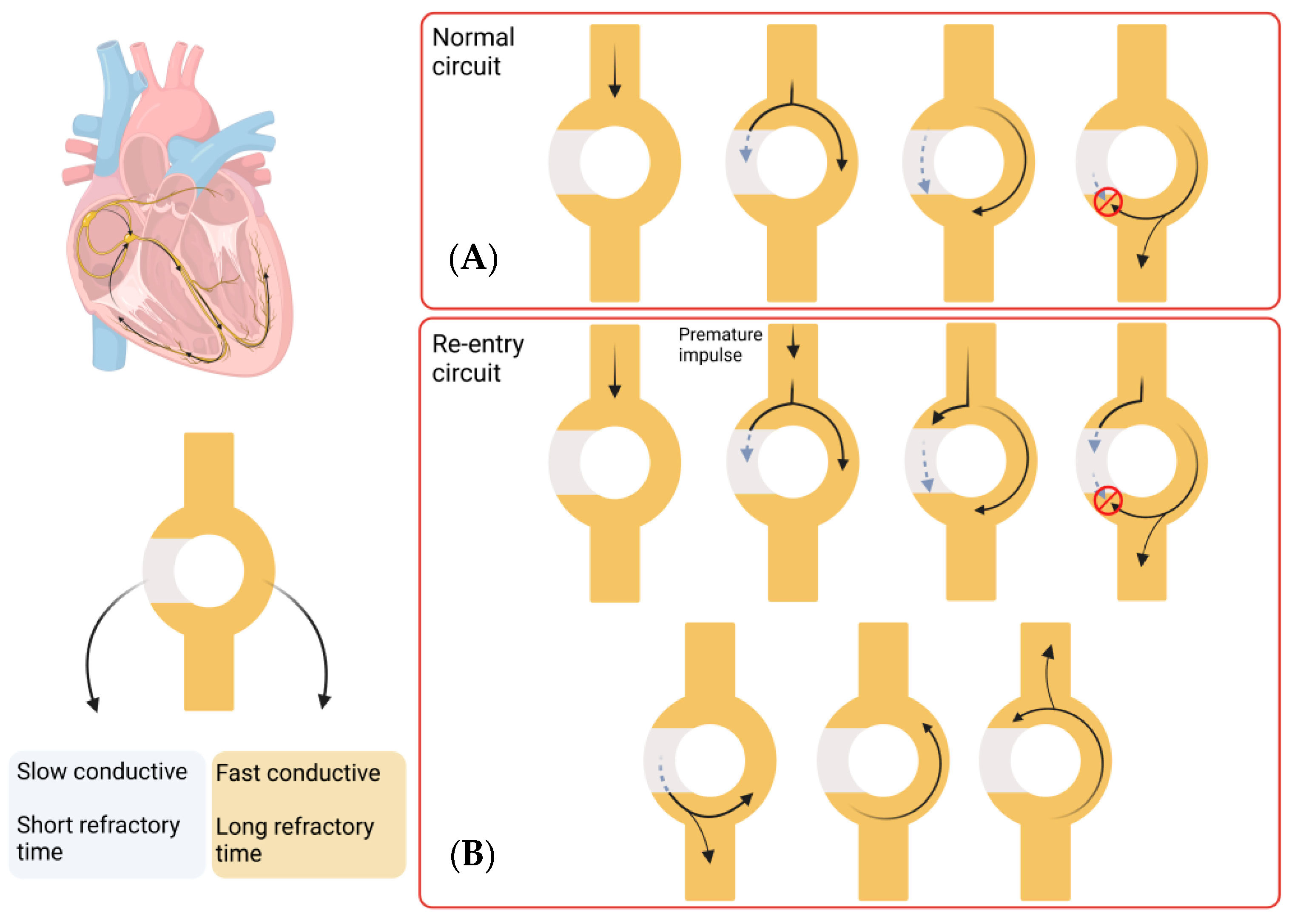
1.2. Catheter Ablation
1.3. Limitations of Current Treatment Approaches
2. Current Experimental Models of Atrial Fibrillation
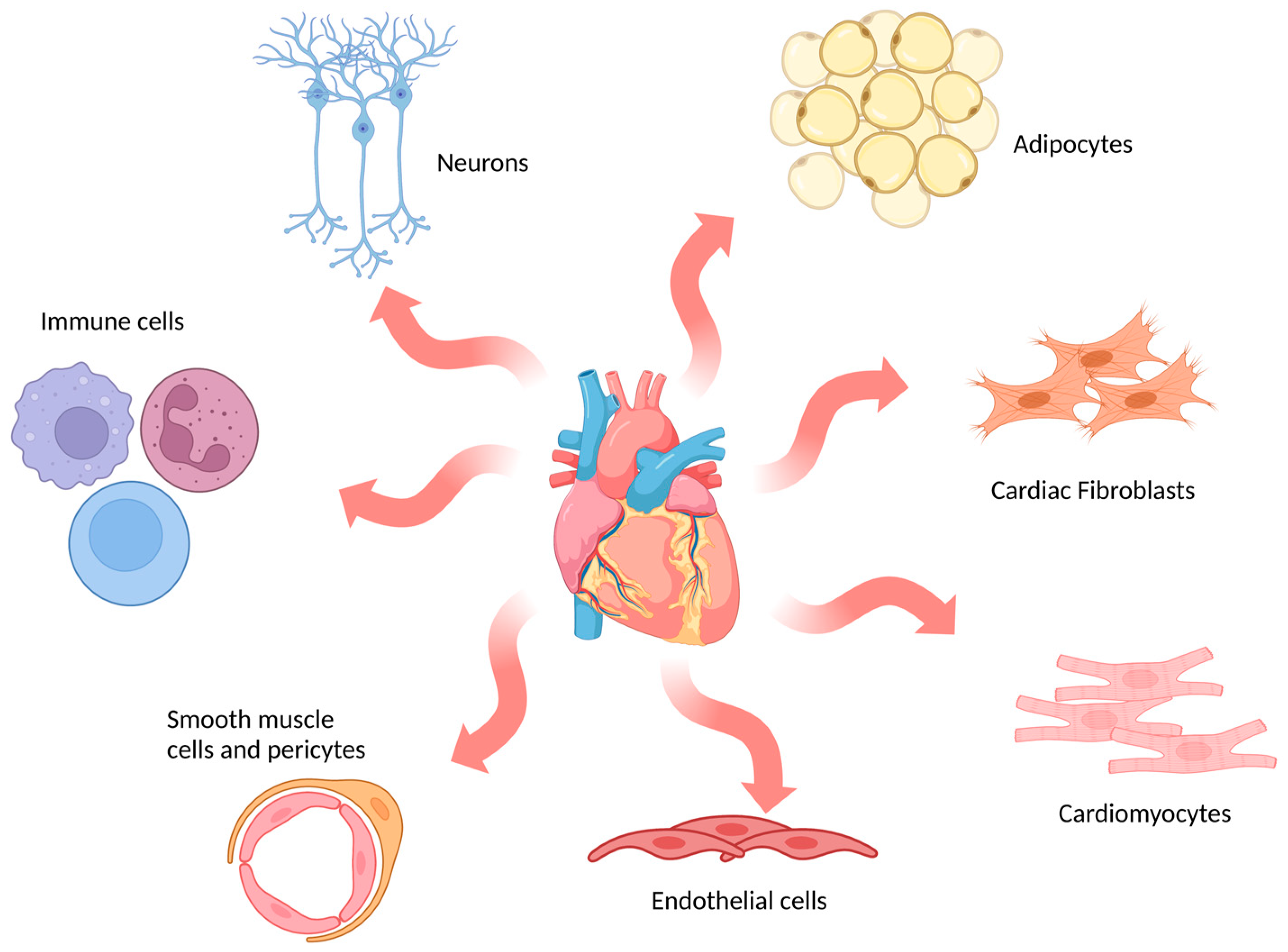
2.1. Ex Vivo Models
2.2. In Vitro Models
2.2.1. Adherent Cell (2D) Models
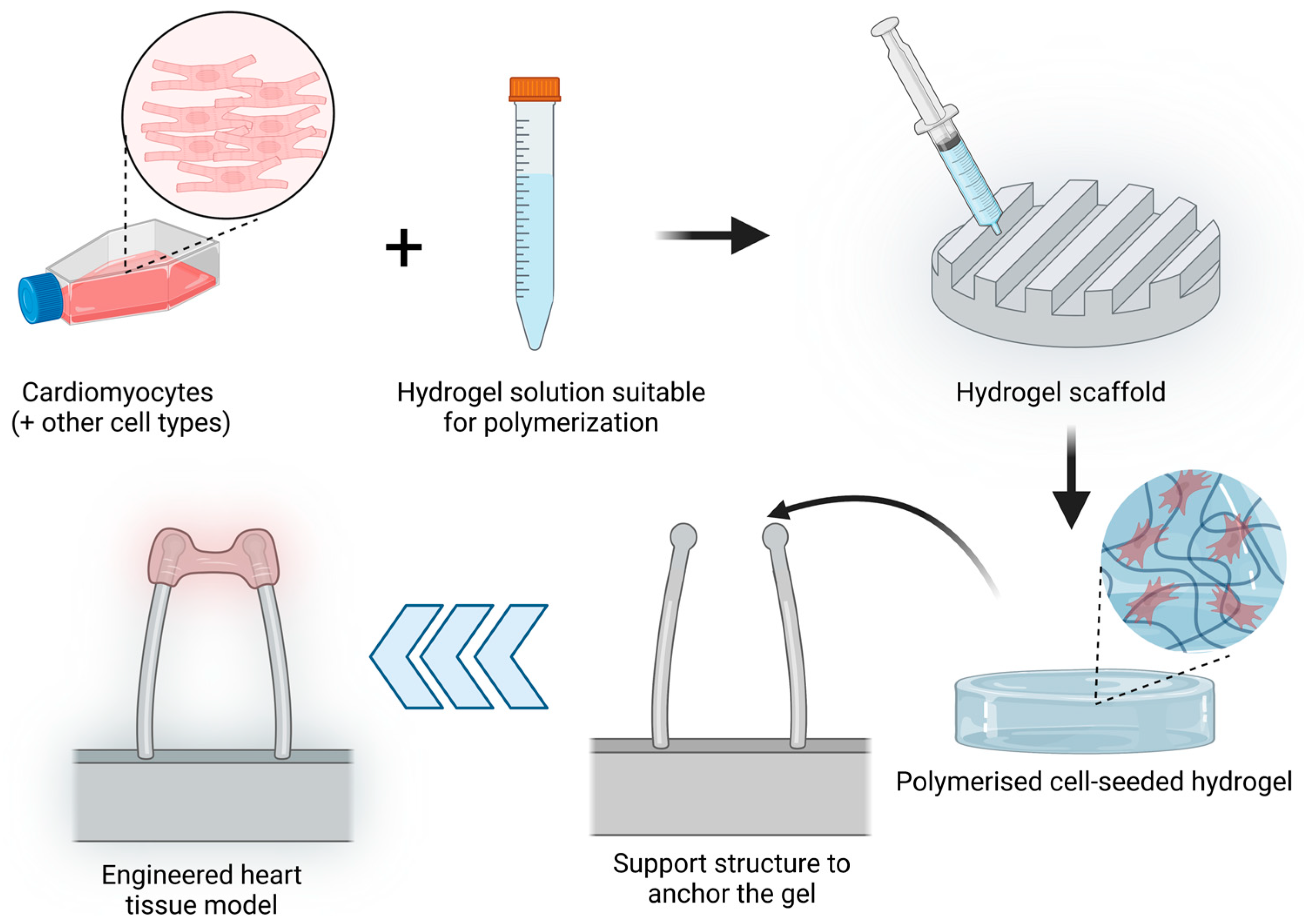
2.2.2. 3D Cell Culture Models
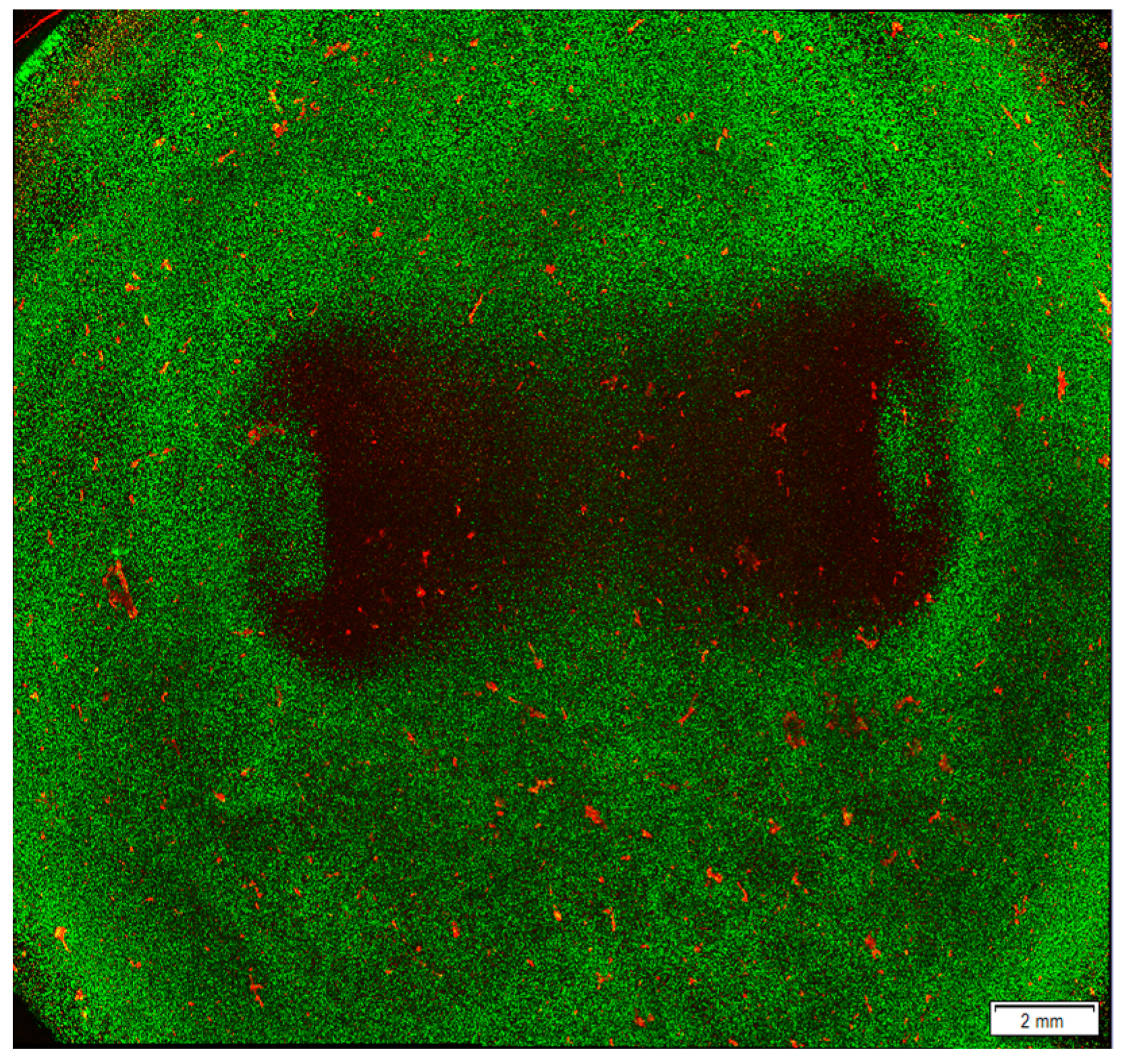
3D Hydrogels
3D Organoids
3. Conclusions and Future Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Staerk, L.; Sherer, J.A.; Ko, D.; Benjamin, E.J.; Helm, R.H. Atrial fibrillation: Epidemiology, pathophysiology, and clinical outcomes. Circ. Res. 2017, 120, 1501–1517. [Google Scholar] [CrossRef]
- Krijthe, B.P.; Kunst, A.; Benjamin, E.J.; Lip, G.Y.H.; Franco, O.H.; Hofman, A.; Witteman, J.C.M.; Stricker, B.H.; Heeringa, J. Projections on the number of individuals with atrial fibrillation in the European Union, from 2000 to 2060. Eur. Heart J. 2013, 34, 2746–2751. [Google Scholar] [CrossRef] [PubMed]
- Morin, D.P.; Bernard, M.L.; Madias, C.; Rogers, P.A.; Thihalolipavan, S.; Estes, N.A.M. The state of the art: Atrial fibrillation epidemiology, prevention, and treatment. Mayo Clin. Proc. 2016, 91, 1778–1810. [Google Scholar] [CrossRef] [PubMed]
- Wańkowicz, P.; Staszewski, J.; Dębiec, A.; Nowakowska-Kotas, M.; Szylińska, A.; Rotter, I. Ischemic Stroke Risk Factors in Patients with Atrial Fibrillation Treated with New Oral Anticoagulants. J. Clin. Med. 2021, 10, 1223. [Google Scholar] [CrossRef] [PubMed]
- Gawałko, M.; Kapłon-Cieślicka, A.; Hohl, M.; Dobrev, D.; Linz, D. COVID-19 associated atrial fibrillation: Incidence, putative mechanisms and potential clinical implications. Int. J. Cardiol. Heart Vasc. 2020, 30, 100631. [Google Scholar] [CrossRef] [PubMed]
- January, C.T.; Wann, L.S.; Alpert, J.S.; Calkins, H.; Cigarroa, J.E.; Cleveland, J.C.; Conti, J.B.; Ellinor, P.T.; Ezekowitz, M.D.; Field, M.E.; et al. 2014 AHA/ACC/HRS guideline for the management of patients with atrial fibrillation: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and the Heart Rhythm Society. J. Am. Coll. Cardiol. 2014, 64, e1–e76. [Google Scholar] [CrossRef]
- Haïssaguerre, M.; Jaïs, P.; Shah, D.C.; Takahashi, A.; Hocini, M.; Quiniou, G.; Garrigue, S.; Le Mouroux, A.; Le Métayer, P.; Clémenty, J. Spontaneous initiation of atrial fibrillation by ectopic beats originating in the pulmonary veins. N. Engl. J. Med. 1998, 339, 659–666. [Google Scholar] [CrossRef]
- Wu, T.J.; Liang, K.W.; Ting, C.T. Relation between the rapid focal activation in the pulmonary vein and the maintenance of paroxysmal atrial fibrillation. Pacing Clin. Electrophysiol. 2001, 24, 902–905. [Google Scholar] [CrossRef]
- Sanders, P.; Morton, J.B.; Davidson, N.C.; Spence, S.J.; Vohra, J.K.; Sparks, P.B.; Kalman, J.M. Electrical remodeling of the atria in congestive heart failure: Electrophysiological and electroanatomic mapping in humans. Circulation 2003, 108, 1461–1468. [Google Scholar] [CrossRef]
- Mahida, S.; Sacher, F.; Derval, N.; Berte, B.; Yamashita, S.; Hooks, D.; Denis, A.; Amraoui, S.; Hocini, M.; Haissaguerre, M.; et al. Science linking pulmonary veins and atrial fibrillation. Arrhythm. Electrophysiol. Rev. 2015, 4, 40–43. [Google Scholar] [CrossRef]
- Azevedo, P.S.; Polegato, B.F.; Minicucci, M.F.; Paiva, S.A.R.; Zornoff, L.A.M. Cardiac remodeling: Concepts, clinical impact, pathophysiological mechanisms and pharmacologic treatment. Arq. Bras. Cardiol. 2016, 106, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, A.J.; Abascal, V.M.; Sheehan, M.; Oertel, L.B.; Harrigan, P.; Hughes, R.A.; Weyman, A.E. Atrial enlargement as a consequence of atrial fibrillation. A prospective echocardiographic study. Circulation 1990, 82, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Heijman, J.; Voigt, N.; Nattel, S.; Dobrev, D. Cellular and molecular electrophysiology of atrial fibrillation initiation, maintenance, and progression. Circ. Res. 2014, 114, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Veenhuyzen, G.D.; Simpson, C.S.; Hoshiar, A. Atrial fibrillation. CMAJ 2004, 171, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Lafuente-Lafuente, C.; Valembois, L.; Bergmann, J.-F.; Belmin, J. Antiarrhythmics for maintaining sinus rhythm after cardioversion of atrial fibrillation. Cochrane Database Syst. Rev. 2015, 3, CD005049. [Google Scholar] [CrossRef]
- Zimetbaum, P. Antiarrhythmic drug therapy for atrial fibrillation. Circulation 2012, 125, 381–389. [Google Scholar] [CrossRef]
- EM, V.W. A classification of antiarrhythmic actions reassessed after a decade of new drugs. J. Clin. Pharmacol. 1984, 24, 129–147. [Google Scholar]
- Roden, D.M. Risks and benefits of antiarrhythmic therapy. N. Engl. J. Med. 1994, 331, 785–791. [Google Scholar]
- Di Biase, L.; Burkhardt, J.D.; Mohanty, P.; Sanchez, J.; Mohanty, S.; Horton, R.; Gallinghouse, G.J.; Bailey, S.M.; Zagrodzky, J.D.; Santangeli, P.; et al. Left atrial appendage: An underrecognized trigger site of atrial fibrillation. Circulation 2010, 122, 109–118. [Google Scholar] [CrossRef]
- Bi, S.; Jia, F.; Lv, C.; He, Q.; Xu, X.; Xue, Z.; Su, S. Preclinical study of biphasic asymmetric pulsed field ablation. Front. Cardiovasc. Med. 2022, 9, 859480. [Google Scholar] [CrossRef]
- Calkins, H.; Kuck, K.H.; Cappato, R.; Brugada, J.; Camm, A.J.; Chen, S.A.; Crijns, H.J.; Damiano, R.J., Jr.; Davies, D.W.; DiMarco, J.; et al. Heart Rhythm Society Task Force on Catheter and Surgical Ablation of Atrial Fibrillation. Heart Rhythm 2012, 9, 632–696.e21. [Google Scholar] [PubMed]
- Calkins, H.; Reynolds, M.R.; Spector, P.; Sondhi, M.; Xu, Y.; Martin, A.; Williams, C.J.; Sledge, I. Treatment of atrial fibrillation with antiarrhythmic drugs or radiofrequency ablation: Two systematic literature reviews and meta-analyses. Circ. Arrhythm. Electrophysiol. 2009, 2, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Haegeli, L.M.; Calkins, H. Catheter ablation of atrial fibrillation: An update. Eur. Heart J. 2014, 35, 2454–2459. [Google Scholar] [CrossRef] [PubMed]
- He, D.S.; Zimmer, J.E.; Hynynen, K.; Marcus, F.I.; Caruso, A.C.; Lampe, L.F.; Aguirre, M.L. Application of ultrasound energy for intracardiac ablation of arrhythmias. Eur. Heart J. 1995, 16, 961–966. [Google Scholar] [CrossRef]
- Engel, D.J.; Muratore, R.; Hirata, K.; Otsuka, R.; Fujikura, K.; Sugioka, K.; Marboe, C.; Lizzi, F.L.; Homma, S. Myocardial lesion formation using high-intensity focused ultrasound. J. Am. Soc. Echocardiogr. 2006, 19, 932–937. [Google Scholar] [CrossRef]
- Schmidt, B.; Antz, M.; Ernst, S.; Ouyang, F.; Falk, P.; Chun, J.K.R.; Kuck, K.-H. Pulmonary vein isolation by high-intensity focused ultrasound: First-in-man study with a steerable balloon catheter. Heart Rhythm 2007, 4, 575–584. [Google Scholar] [CrossRef]
- Jin, E.S.; Wang, P.J. Cryoballoon ablation for atrial fibrillation: A comprehensive review and practice guide. Korean Circ. J. 2018, 48, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, D.K. Mechanisms of tissue destruction following cryosurgery. Ann. R. Coll. Surg. Engl. 1984, 66, 313–318. [Google Scholar] [PubMed]
- Kaszala, K.; Ellenbogen, K.A. Biophysics of the second-generation cryoballoon: Cryobiology of the big freeze. Circ. Arrhythm. Electrophysiol. 2015, 8, 15–17. [Google Scholar] [CrossRef]
- Nair, K.K.M.; Danon, A.; Valaparambil, A.; Koruth, J.S.; Singh, S.M. Atrioesophageal fistula: A review. J. Atr. Fibrillation 2015, 8, 1331. [Google Scholar]
- Nuccitelli, R. Application of pulsed electric fields to cancer therapy. Bioelectricity 2018, 1, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Wittkampf, F.H.M.; van Es, R.; Neven, K. Electroporation and its relevance for cardiac catheter ablation. JACC Clin. Electrophysiol. 2018, 4, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Heijman, J.; Guichard, J.-B.; Dobrev, D.; Nattel, S. Translational challenges in atrial fibrillation. Circ. Res. 2018, 122, 752–773. [Google Scholar] [CrossRef] [PubMed]
- Heijman, J.; Algalarrondo, V.; Voigt, N.; Melka, J.; Wehrens, X.H.T.; Dobrev, D.; Nattel, S. The value of basic research insights into atrial fibrillation mechanisms as a guide to therapeutic innovation: A critical analysis. Cardiovasc. Res. 2016, 109, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Halațiu, V.B.; Perian, M.; Balan, A.I.; Scridon, A. Transesophageal atrial burst pacing for atrial fibrillation induction in rats. J. Vis. Exp. 2022, 180, e63567. [Google Scholar] [CrossRef]
- Scridon, A.; Perian, M.; Grigoraş, T.; Halaţiu, V.; Vântu, A.; Balan, A.; Cosma, I.; Barmou, A.; Finascu, B.; Moldovan, D.; et al. Spontaneous atrial fibrillation after long-term transesophageal atrial burst pacing in rats. Technical and procedural approach to a new in vivo atrial fibrillation model. Rev. Romana Med. Lab. 2018, 26, 105–111. [Google Scholar] [CrossRef]
- Tayebjee, M.H.; Hunter, R.J.; Baker, V.; Creta, A.; Duncan, E.; Sporton, S.; Earley, M.J.; Schilling, R.J. Pulmonary vein isolation with radiofrequency ablation followed by cryotherapy: A novel strategy to improve clinical outcomes following catheter ablation of paroxysmal. Europace 2011, 13, 1250–1255. [Google Scholar] [CrossRef]
- Schüttler, D.; Bapat, A.; Kääb, S.; Lee, K.; Tomsits, P.; Clauss, S.; Hucker, W.J. Animal models of atrial fibrillation. Circ. Res. 2020, 127, 91–110. [Google Scholar] [CrossRef]
- Bussek, A.; Schmidt, M.; Bauriedl, J.; Ravens, U.; Wettwer, E.; Lohmann, H. Cardiac tissue slices with prolonged survival for in vitro drug safety screening. J. Pharmacol. Toxicol. Methods 2012, 66, 145–151. [Google Scholar] [CrossRef]
- Rosen, M. Isolated tissue models and proarrhythmia. Eur. Heart J. Suppl. 2001, 3, K64–K69. [Google Scholar] [CrossRef]
- Boukens, B.J.; Sulkin, M.S.; Gloschat, C.R.; Ng, F.S.; Vigmond, E.J.; Efimov, I.R. Transmural APD gradient synchronizes repolarization in the human left ventricular wall. Cardiovasc. Res. 2015, 108, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Jost, N.; Virág, L.; Comtois, P.; Ordög, B.; Szuts, V.; Seprényi, G.; Bitay, M.; Kohajda, Z.; Koncz, I.; Nagy, N.; et al. Ionic mechanisms limiting cardiac repolarization reserve in humans compared to dogs. J. Physiol. 2013, 591, 4189–4206. [Google Scholar] [CrossRef] [PubMed]
- Gramley, F.; Lorenzen, J.; Jedamzik, B.; Gatter, K.; Koellensperger, E.; Munzel, T.; Pezzella, F. Atrial fibrillation is associated with cardiac hypoxia. Cardiovasc. Pathol. 2010, 19, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yi, X.; Li, X.; Jiang, X. Fibroblast growth factor-21 is positively associated with atrial fibrosis in atrial fibrillation patients with rheumatic heart disease. Int. J. Clin. Exp. Pathol. 2015, 8, 14901–14908. [Google Scholar]
- Csepe, T.A.; Hansen, B.J.; Fedorov, V.V. Atrial fibrillation driver mechanisms: Insight from the isolated human heart. Trends Cardiovasc. Med. 2017, 27, 1–11. [Google Scholar] [CrossRef][Green Version]
- Calzolari, V.; De Mattia, L.; Basso, F.; Crosato, M.; Scalon, A.; Squasi, P.A.M.; Del Favero, S.; Cernetti, C. Ablation catheter orientation: In vitro effects on lesion size and in vivo analysis during PVI for atrial fibrillation. Pacing Clin. Electrophysiol. 2020, 43, 1554–1563. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.D.; Ravi, V.; Rhodes, P.; Du-Fay-de-Lavallaz, J.M.; Winterfield, J.; Allen-Proctor, M.; Wasserlauf, J.; Krishnan, K.; Trohman, R.; Sharma, P.S.; et al. Use of infrared thermography to delineate temperature gradients and critical isotherms during catheter ablation with normal and half normal saline: Implications for safety and efficacy. J. Cardiovasc. Electrophysiol. 2021, 32, 2035–2044. [Google Scholar] [CrossRef]
- Masnok, K.; Watanabe, N. Catheter contact area strongly correlates with lesion area in radiofrequency cardiac ablation: An ex vivo porcine heart study. J. Interv. Card. Electrophysiol. 2022, 63, 561–572. [Google Scholar] [CrossRef]
- Huang, S.-T.; Dong, J.-Z.; Du, X.; Wu, J.-H.; Yu, R.-H.; Long, D.-Y.; Ning, M.; Sang, C.-H.; Jiang, C.-X.; Bai, R.; et al. Relationship Between Ablation Lesion Size Estimated by Ablation Index and Different Ablation Settings-an Ex Vivo Porcine Heart Study. J. Cardiovasc. Transl. Res. 2020, 13, 965–969. [Google Scholar] [CrossRef]
- Park, S.; Hwang, J.; Park, J.E.; Ahn, Y.C. Application of ultrasound thermal imaging for monitoring laser ablation in ex vivo cardiac tissue. Lasers Surg. Med. 2020, 52, 218–227. [Google Scholar] [CrossRef]
- Langendorff, O. Untersuchungen am überlebenden Säugethierherzen. Pflügers Arch. 1895, 61, 291–332. [Google Scholar] [CrossRef]
- Ninio, D.M.; Murphy, K.J.; Howe, P.R.; Saint, D.A. Dietary fish oil protects against stretch-induced vulnerability to atrial fibrillation in a rabbit model. J. Cardiovasc. Electrophysiol. 2005, 16, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Sarmast, F. Cholinergic atrial fibrillation: IK,ACh gradients determine unequal left/right atrial frequencies and rotor dynamics. Cardiovasc. Res. 2003, 59, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, J.E.; Diness, J.G.; Sheykhzade, M.; Grunnet, M.; Jespersen, T. Synergistic antiarrhythmic effect of combining inhibition of Ca2+-activated K+ (SK) channels and voltage-gated Na+ channels in an isolated heart model of atrial fibrillation. Heart Rhythm 2015, 12, 409–418. [Google Scholar] [CrossRef]
- Liao, C.; Akazawa, H.; Tamagawa, M.; Ito, K.; Yasuda, N.; Kudo, Y.; Yamamoto, R.; Ozasa, Y.; Fujimoto, M.; Wang, P.; et al. Cardiac mast cells cause atrial fibrillation through PDGF-A-mediated fibrosis in pressure-overloaded mouse hearts. J. Clin. Investig. 2010, 120, 242–253. [Google Scholar] [CrossRef]
- Chen, J.; Mandapati, R.; Berenfeld, O.; Skanes, A.C.; Gray, R.A.; Jalife, J. Dynamics of wavelets and their role in atrial fibrillation in the isolated sheep heart. Cardiovasc. Res. 2000, 48, 220–232. [Google Scholar] [CrossRef]
- Pak, H.-N.; Oh, Y.-S.; Liu, Y.-B.; Wu, T.-J.; Karagueuzian, H.S.; Lin, S.-F.; Chen, P.-S. Catheter ablation of ventricular fibrillation in rabbit ventricles treated with beta-blockers. Circulation 2003, 108, 3149–3156. [Google Scholar] [CrossRef]
- Deng, C.X.; Qu, F.; Nikolski, V.P.; Zhou, Y.; Efimov, I.R. Fluorescence imaging for real-time monitoring of high-intensity focused ultrasound cardiac ablation. Ann. Biomed. Eng. 2005, 33, 1352–1359. [Google Scholar] [CrossRef]
- Skrzypiec-Spring, M.; Grotthus, B.; Szelag, A.; Schulz, R. Isolated heart perfusion according to Langendorff—Still viable in the new millennium. J. Pharmacol. Toxicol. Methods 2007, 55, 113–126. [Google Scholar] [CrossRef]
- Doring, H.J.; Dehnert, H. The Isolated Perfused Heart according to Langendorff; BVM-Biomesstechnik Verlag: Berlin/Heidelberg, Germany, 1987. [Google Scholar]
- Litviňuková, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Worth, C.L.; Lindberg, E.L.; Kanda, M.; Polanski, K.; Heinig, M.; Lee, M.; et al. Cells of the adult human heart. Nature 2020, 588, 466–472. [Google Scholar] [CrossRef]
- Woodcock, E.A.; Matkovich, S.J. Cardiomyocytes structure, function and associated pathologies. Int. J. Biochem. Cell Biol. 2005, 37, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Doppler, S.A.; Carvalho, C.; Lahm, H.; Deutsch, M.-A.; Dreßen, M.; Puluca, N.; Lange, R.; Krane, M. Cardiac fibroblasts: More than mechanical support. J. Thorac. Dis. 2017, 9, S36–S51. [Google Scholar] [CrossRef] [PubMed]
- Simmons, M.A.; Creazzo, T.; Hartzell, H.C. A time-dependent and voltage-sensitive K+ current in single cells from frog atrium. J. Gen. Physiol. 1986, 88, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Jahnel, U.; Nawrath, H.; Carmeliet, E.; Vereecke, J. Depolarization-induced influx of sodium in response to phenylephrine in rat atrial heart muscle. J. Physiol. 1991, 432, 621–637. [Google Scholar] [CrossRef]
- Kuo, P.-L.; Lee, H.; Bray, M.-A.; Geisse, N.A.; Huang, Y.-T.; Adams, W.J.; Sheehy, S.P.; Parker, K.K. Myocyte shape regulates lateral registry of sarcomeres and contractility. Am. J. Pathol. 2012, 181, 2030–2037. [Google Scholar] [CrossRef]
- Vites, A.M.; Pappano, A.J. Distinct modes of inhibition by ruthenium red and ryanodine of calcium-induced calcium release in avian atrium. J. Pharmacol. Exp. Ther. 1994, 268, 1476–1484. [Google Scholar]
- Cheung, J.Y.; Tillotson, D.L.; Yelamarty, R.V.; Scaduto, R.C. Cytosolic free calcium concentration in individual cardiac myocytes in primary culture. Am. J. Physiol. 1989, 256, C1120–C1130. [Google Scholar] [CrossRef]
- Sowell, B.; Fast, V.G. Ionic mechanism of shock-induced arrhythmias: Role of intracellular calcium. Heart Rhythm 2012, 9, 96–104. [Google Scholar] [CrossRef][Green Version]
- Maqsood, M.I.; Matin, M.M.; Bahrami, A.R.; Ghasroldasht, M.M. Immortality of cell lines: Challenges and advantages of establishment. Cell Biol. Int. 2013, 37, 1038–1045. [Google Scholar] [CrossRef]
- Claycomb, W.C.; Lanson, N.A.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J. HL-1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef]
- Ke, L.; Qi, X.Y.; Dijkhuis, A.-J.; Chartier, D.; Nattel, S.; Henning, R.H.; Kampinga, H.H.; Brundel, B.J.J.M. Calpain mediates cardiac troponin degradation and contractile dysfunction in atrial fibrillation. J. Mol. Cell. Cardiol. 2008, 45, 685–693. [Google Scholar] [CrossRef]
- Wiersma, M.; Meijering, R.A.M.; Qi, X.-Y.; Zhang, D.; Liu, T.; Hoogstra-Berends, F.; Sibon, O.C.M.; Henning, R.H.; Nattel, S.; Brundel, B.J.J.M. Endoplasmic reticulum stress is associated with autophagy and cardiomyocyte remodeling in experimental and human atrial fibrillation. J. Am. Heart Assoc. 2017, 6, e006458. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.; Deng, C.-Y.; Wu, S.-L.; Xiao, D.-Z.; Yu, X.-Y.; Kuang, S.-J.; Lin, Q.-X.; Shan, Z.-X. Involvement of Src in L-type Ca2+ channel depression induced by macrophage migration inhibitory factor in atrial myocytes. J. Mol. Cell. Cardiol. 2009, 47, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Kimes, B.W.; Brandt, B.L. Properties of a clonal muscle cell line from rat heart. Exp. Cell Res. 1976, 98, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Chueh, P.J.; Tseng, C.-T.; Liu, K.-Y.; Tsai, H.-Y.; Kuo, W.-W.; Chou, M.-Y.; Yang, J.-J. ZAK re-programs atrial natriuretic factor expression and induces hypertrophic growth in H9c2 cardiomyoblast cells. Biochem. Biophys. Res. Commun. 2004, 324, 973–980. [Google Scholar] [CrossRef]
- Vindis, C.; D’Angelo, R.; Mucher, E.; Nègre-Salvayre, A.; Parini, A.; Mialet-Perez, J. Essential role of TRPC1 channels in cardiomyoblasts hypertrophy mediated by 5-HT2A serotonin receptors. Biochem. Biophys. Res. Commun. 2010, 391, 979–983. [Google Scholar] [CrossRef]
- Davidson, M.M.; Nesti, C.; Palenzuela, L.; Walker, W.F.; Hernandez, E.; Protas, L.; Hirano, M.; Isaac, N.D. Novel cell lines derived from adult human ventricular cardiomyocytes. J. Mol. Cell. Cardiol. 2005, 39, 133–147. [Google Scholar] [CrossRef]
- Dutta, D.; Xu, J.; Kim, J.-S.; Dunn, W.A.; Leeuwenburgh, C. Upregulated autophagy protects cardiomyocytes from oxidative stress-induced toxicity. Autophagy 2013, 9, 328–344. [Google Scholar] [CrossRef]
- Cui, L.; Guo, J.; Zhang, Q.; Yin, J.; Li, J.; Zhou, W.; Zhang, T.; Yuan, H.; Zhao, J.; Zhang, L.; et al. Erythropoietin activates SIRT1 to protect human cardiomyocytes against doxorubicin-induced mitochondrial dysfunction and toxicity. Toxicol. Lett. 2017, 275, 28–38. [Google Scholar] [CrossRef]
- Wen, J.; Shen, J.; Zhou, Y.; Zhao, X.; Dai, Z.; Jin, Y. Pyrroloquinoline quinone attenuates isoproterenol hydrochloride-induced cardiac hypertrophy in AC16 cells by inhibiting the NF-κB signaling pathway. Int. J. Mol. Med. 2020, 45, 873–885. [Google Scholar] [CrossRef]
- Baena-Montes, J.M.; O’Halloran, T.; Clarke, C.; Donaghey, K.; Dunne, E.; O’Halloran, M.; Quinlan, L.R. Electroporation parameters for human cardiomyocyte ablation in vitro. J. Cardiovasc. Dev. Dis. 2022, 9, 240. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Banerjee, I. Recent advances in the applications of iPSC technology. Curr. Opin. Biotechnol. 2019, 60, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Vogel, G. Stem cells. Diseases in a dish take off. Science 2010, 330, 1172–1173. [Google Scholar] [CrossRef]
- Burridge, P.W.; Keller, G.; Gold, J.D.; Wu, J.C. Production of de novo cardiomyocytes: Human pluripotent stem cell differentiation and direct reprogramming. Cell Stem Cell 2012, 10, 16–28. [Google Scholar]
- Funakoshi, S.; Yoshida, Y. Recent progress of iPSC technology in cardiac diseases. Arch. Toxicol. 2021, 95, 3633–3650. [Google Scholar]
- Knollmann, B.C. Induced pluripotent stem cell-derived cardiomyocytes: Boutique science or valuable arrhythmia model? Circ. Res. 2013, 112, 969–976; discussion 976. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Ono, K.; Hata, S.; Suzuki, T.; Kumamoto, H.; Sasano, H. The advantages of co-culture over mono cell culture in simulating in vivo environment. J. Steroid Biochem. Mol. Biol. 2012, 131, 68–75. [Google Scholar] [CrossRef]
- Cartledge, J.E.; Kane, C.; Dias, P.; Tesfom, M.; Clarke, L.; Mckee, B.; Al Ayoubi, S.; Chester, A.; Yacoub, M.H.; Camelliti, P.; et al. Functional crosstalk between cardiac fibroblasts and adult cardiomyocytes by soluble mediators. Cardiovasc. Res. 2015, 105, 260–270. [Google Scholar] [CrossRef]
- Thompson, S.A.; Copeland, C.R.; Reich, D.H.; Tung, L. Mechanical coupling between myofibroblasts and cardiomyocytes slows electric conduction in fibrotic cell monolayers. Circulation 2011, 123, 2083–2093. [Google Scholar] [CrossRef]
- Rahmutula, D.; Marcus, G.M.; Wilson, E.E.; Ding, C.-H.; Xiao, Y.; Paquet, A.C.; Barbeau, R.; Barczak, A.J.; Erle, D.J.; Olgin, J.E. Molecular basis of selective atrial fibrosis due to overexpression of transforming growth factor-β1. Cardiovasc. Res. 2013, 99, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Rohr, S. Arrhythmogenic implications of fibroblast-myocyte interactions. Circ. Arrhythm. Electrophysiol. 2012, 5, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Tibbitt, M.W.; Anseth, K.S. Hydrogels as extracellular matrix mimics for 3D cell culture. Biotechnol. Bioeng. 2009, 103, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Sirish, P.; Li, N.; Timofeyev, V.; Zhang, X.-D.; Wang, L.; Yang, J.; Lee, K.S.S.; Bettaieb, A.; Ma, S.M.; Lee, J.H.; et al. Molecular mechanisms and new treatment paradigm for atrial fibrillation. Circ. Arrhythm. Electrophysiol. 2016, 9, e003721. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Liu, X.; Wang, Q.-X.; Guo, M.; Liu, F.; Song, Z.-P.; Zhang, D.-D. Beneficial effects of pioglitazone on atrial structural and electrical remodeling in vitro cellular models. J. Mol. Cell. Cardiol. 2013, 65, 1–8. [Google Scholar] [CrossRef]
- Casciola, M.; Feaster, T.K.; Caiola, M.J.; Keck, D.; Blinova, K. Human in vitro assay for irreversible electroporation cardiac ablation. Front. Physiol. 2022, 13, 1064168. [Google Scholar] [CrossRef]
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. J. Cell Sci. 2010, 123, 4201–4213. [Google Scholar] [CrossRef]
- Whitesides, G.M.; Ostuni, E.; Takayama, S.; Jiang, X.; Ingber, D.E. Soft lithography in biology and biochemistry. Annu. Rev. Biomed. Eng. 2001, 3, 335–373. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Whitesides, G.M. Soft Lithography. Annu. Rev. Mater. Sci. 1998, 28, 153–184. [Google Scholar] [CrossRef]
- Bélisle, J.M.; Kunik, D.; Costantino, S. Rapid multicomponent optical protein patterning. Lab Chip 2009, 9, 3580–3585. [Google Scholar] [CrossRef]
- Badie, N.; Bursac, N. Novel micropatterned cardiac cell cultures with realistic ventricular microstructure. Biophys. J. 2009, 96, 3873–3885. [Google Scholar] [CrossRef]
- McDevitt, T.C.; Angello, J.C.; Whitney, M.L.; Reinecke, H.; Hauschka, S.D.; Murry, C.E.; Stayton, P.S. In Vitro generation of differentiated cardiac myofibers on micropatterned laminin surfaces. J. Biomed. Mater. Res. 2002, 60, 472–479. [Google Scholar] [CrossRef]
- Thomas, S.P.; Bircher-Lehmann, L.; Thomas, S.A.; Zhuang, J.; Saffitz, J.E.; Kléber, A.G. Synthetic strands of neonatal mouse cardiac myocytes: Structural and electrophysiological properties. Circ. Res. 2000, 87, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Biendarra-Tiegs, S.M.; Yechikov, S.; Shergill, B.; Brumback, B.; Takahashi, K.; Shirure, V.S.; Gonzalez, R.E.; Houshmand, L.; Zhong, D.; Weng, K.-C.; et al. An iPS-derived in vitro model of human atrial conduction. Physiol. Rep. 2022, 10, e15407. [Google Scholar] [CrossRef] [PubMed]
- Laksman, Z.; Wauchop, M.; Lin, E.; Protze, S.; Lee, J.; Yang, W.; Izaddoustdar, F.; Shafaattalab, S.; Gepstein, L.; Tibbits, G.F.; et al. Modeling Atrial Fibrillation using Human Embryonic Stem Cell-Derived Atrial Tissue. Sci. Rep. 2017, 7, 5268. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Yasuda, S.; Katoh, M.; Yamada, K.P.; Yamashita, H.; Saeki, Y.; Sunagawa, K.; Nagai, R.; Hisada, T.; Sugiura, S. Single cell mechanics of rat cardiomyocytes under isometric, unloaded, and physiologically loaded conditions. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H196–H202. [Google Scholar] [CrossRef] [PubMed]
- Häneke, T.; Sahara, M. Progress in bioengineering strategies for heart regenerative medicine. Int. J. Mol. Sci. 2022, 23, 3482. [Google Scholar] [CrossRef]
- Mishra, B.; Upadhyay, M.; Adena, S.R. Hydrogels: An introduction to a controlled drug delivery device, synthesis and application in drug delivery and tissue engineering. Austin. J. Biomed. Eng. 2017, 4, 1037. [Google Scholar]
- Eschenhagen, T.; Fink, C.; Remmers, U.; Scholz, H.; Wattchow, J.; Weil, J.; Zimmermann, W.; Dohmen, H.H.; Schäfer, H.; Bishopric, N.; et al. Three-dimensional reconstitution of embryonic cardiomyocytes in a collagen matrix: A new heart muscle model system. FASEB J. 1997, 11, 683–694. [Google Scholar] [CrossRef]
- Hansen, A.; Eder, A.; Bönstrup, M.; Flato, M.; Mewe, M.; Schaaf, S.; Aksehirlioglu, B.; Schwoerer, A.P.; Uebeler, J.; Eschenhagen, T. Development of a drug screening platform based on engineered heart tissue. Circ. Res. 2010, 107, 35–44. [Google Scholar] [CrossRef]
- Zimmermann, W.H.; Schneiderbanger, K.; Schubert, P.; Didié, M.; Münzel, F.; Heubach, J.F.; Kostin, S.; Neuhuber, W.L.; Eschenhagen, T. Tissue engineering of a differentiated cardiac muscle construct. Circ. Res. 2002, 90, 223–230. [Google Scholar] [CrossRef]
- Fink, C.; Ergün, S.; Kralisch, D.; Remmers, U.; Weil, J.; Eschenhagen, T. Chronic stretch of engineered heart tissue induces hypertrophy and functional improvement. FASEB J. 2000, 14, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Mannhardt, I.; Breckwoldt, K.; Letuffe-Brenière, D.; Schaaf, S.; Schulz, H.; Neuber, C.; Benzin, A.; Werner, T.; Eder, A.; Schulze, T.; et al. Human engineered heart tissue: Analysis of contractile force. Stem Cell Rep. 2016, 7, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Hirt, M.N.; Boeddinghaus, J.; Mitchell, A.; Schaaf, S.; Börnchen, C.; Müller, C.; Schulz, H.; Hubner, N.; Stenzig, J.; Stoehr, A.; et al. Functional improvement and maturation of rat and human engineered heart tissue by chronic electrical stimulation. J. Mol. Cell. Cardiol. 2014, 74, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Liau, B.; Jackman, C.P.; Li, Y.; Bursac, N. Developmental stage-dependent effects of cardiac fibroblasts on function of stem cell-derived engineered cardiac tissues. Sci. Rep. 2017, 7, 42290. [Google Scholar] [CrossRef]
- Tiburcy, M.; Hudson, J.E.; Balfanz, P.; Schlick, S.; Meyer, T.; Chang Liao, M.-L.; Levent, E.; Raad, F.; Zeidler, S.; Wingender, E.; et al. Defined engineered human myocardium with advanced maturation for applications in heart failure modeling and repair. Circulation 2017, 135, 1832–1847. [Google Scholar] [CrossRef]
- Goldfracht, I.; Protze, S.; Shiti, A.; Setter, N.; Gruber, A.; Shaheen, N.; Nartiss, Y.; Keller, G.; Gepstein, L. Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes. Nat. Commun. 2020, 11, 75. [Google Scholar] [CrossRef]
- Eder, A.; Vollert, I.; Hansen, A.; Eschenhagen, T. Human engineered heart tissue as a model system for drug testing. Adv. Drug Deliv. Rev. 2016, 96, 214–224. [Google Scholar] [CrossRef]
- Maruyama, H.; Yokota, Y.; Hosono, K.; Arai, F. Hydrogel Heart Model with Temperature Memory Properties for Surgical Simulation. Sensors 2019, 19, 1102. [Google Scholar] [CrossRef]
- Yokota, Y.; Maruyama, H.; Arai, F. Hydrogel Heart Model Having Temperature Indicating Function. In Proceedings of the 2018 International Symposium on Micro-NanoMechatronics and Human Science (MHS), Nagoya, Japan, 9–12 December 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 1–2. [Google Scholar]
- Arena, C.B.; Szot, C.S.; Garcia, P.A.; Rylander, M.N.; Davalos, R.V. A three-dimensional in vitro tumor platform for modeling therapeutic irreversible electroporation. Biophys. J. 2012, 103, 2033–2042. [Google Scholar] [CrossRef]
- Wasson, E.M.; Alinezhadbalalami, N.; Brock, R.M.; Allen, I.C.; Verbridge, S.S.; Davalos, R.V. Understanding the role of calcium-mediated cell death in high-frequency irreversible electroporation. Bioelectrochemistry 2020, 131, 107369. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.J.; Van Es, J.H.; Van den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Huch, M.; Gehart, H.; van Boxtel, R.; Hamer, K.; Blokzijl, F.; Verstegen, M.M.A.; Ellis, E.; van Wenum, M.; Fuchs, S.A.; de Ligt, J.; et al. Long-term culture of genome-stable bipotent stem cells from adult human liver. Cell 2015, 160, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Vela, I.; Sboner, A.; Iaquinta, P.J.; Karthaus, W.R.; Gopalan, A.; Dowling, C.; Wanjala, J.N.; Undvall, E.A.; Arora, V.K.; et al. Organoid cultures derived from patients with advanced prostate cancer. Cell 2014, 159, 176–187. [Google Scholar] [CrossRef]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef]
- Kumar, A.; Starly, B. Large scale industrialized cell expansion: Producing the critical raw material for biofabrication processes. Biofabrication 2015, 7, 044103. [Google Scholar] [CrossRef]
- Tung, Y.-C.; Hsiao, A.Y.; Allen, S.G.; Torisawa, Y.; Ho, M.; Takayama, S. High-throughput 3D spheroid culture and drug testing using a 384 hanging drop array. Analyst 2011, 136, 473–478. [Google Scholar] [CrossRef]
- Hoarau-Véchot, J.; Rafii, A.; Touboul, C.; Pasquier, J. Halfway between 2D and Animal Models: Are 3D Cultures the Ideal Tool to Study Cancer-Microenvironment Interactions? Int. J. Mol. Sci. 2018, 19, 181. [Google Scholar] [CrossRef]
- Yuhas, J.M.; Li, A.P.; Martinez, A.O.; Ladman, A.J. A simplified method for production and growth of multicellular tumor spheroids. Cancer Res. 1977, 37, 3639–3643. [Google Scholar]
- Tseng, H.; Gage, J.A.; Raphael, R.M.; Moore, R.H.; Killian, T.C.; Grande-Allen, K.J.; Souza, G.R. Assembly of a three-dimensional multitype bronchiole coculture model using magnetic levitation. Tissue Eng. Part C Methods 2013, 19, 665–675. [Google Scholar] [CrossRef]
- Rawal, P.; Tripathi, D.M.; Ramakrishna, S.; Kaur, S. Prospects for 3D bioprinting of organoids. Bio-Des. Manuf. 2021, 4, 627–640. [Google Scholar] [CrossRef]
- Cui, H.; Nowicki, M.; Fisher, J.P.; Zhang, L.G. 3D bioprinting for organ regeneration. Adv. Healthc. Mater. 2017, 6, 1601118. [Google Scholar] [CrossRef] [PubMed]
- Shinnawi, R.; Shaheen, N.; Huber, I.; Shiti, A.; Arbel, G.; Gepstein, A.; Ballan, N.; Setter, N.; Tijsen, A.J.; Borggrefe, M.; et al. Modeling Reentry in the Short QT Syndrome With Human-Induced Pluripotent Stem Cell-Derived Cardiac Cell Sheets. J. Am. Coll. Cardiol. 2019, 73, 2310–2324. [Google Scholar] [CrossRef] [PubMed]
- Pollen, A.A.; Bhaduri, A.; Andrews, M.G.; Nowakowski, T.J.; Meyerson, O.S.; Mostajo-Radji, M.A.; Di Lullo, E.; Alvarado, B.; Bedolli, M.; Dougherty, M.L.; et al. Establishing Cerebral Organoids as Models of Human-Specific Brain Evolution. Cell 2019, 176, 743–756.e17. [Google Scholar] [CrossRef] [PubMed]
- Huch, M.; Knoblich, J.A.; Lutolf, M.P.; Martinez-Arias, A. The hope and the hype of organoid research. Development 2017, 144, 938–941. [Google Scholar] [CrossRef] [PubMed]
- de Souza, N. Organoids. Nat. Methods 2018, 15, 23. [Google Scholar] [CrossRef]
- Munsie, M.; Hyun, I.; Sugarman, J. Ethical issues in human organoid and gastruloid research. Development 2017, 144, 942–945. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Implementation | Advantages | Limitations | References |
|---|---|---|---|---|
| Ex Vivo | Isolated animal whole hearts Heart slices | Controlled environment Highly reproducible experiments Perfusion studies with a complete beating heart Can have human origin Allow physical positioning and opening of a catheter for ablation studies (inside or outside of the heart) | Non-physiological conditions (i.e., blood composition) Whole hearts: complex to maintain for any extended period Heart slices: limited tissue and cell viability; limited supply of human samples | [39,41,46,47,48,49,50,51] |
| In Vitro | 2D adherent models | Rapid screening process Lower costs Wide range of cell options Established protocols Can be human-derived (hiPSC) | Overly simplistic models of the whole heart structure Lacks 3D structure Non-physiological morphology and cell responses Absence of conductivity values for ablation studies | [96,101,102,103,104,105,106,108] |
| 3D models | 3D structure for ablation Potential to measure conductivity Cell–cell and cell–matrix interactions Multiple cell types in physiological disposition (organoids) Can be human-derived (hiPSC), displaying contractile activity | A simple model of the whole heart structure Challenging to produce, manipulate and maintain for extended period Slower screening process Lack of vascular and immune systems | [110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,129,131,132,135,136,139] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baena-Montes, J.M.; Kraśny, M.J.; O’Halloran, M.; Dunne, E.; Quinlan, L.R. In Vitro Models for Improved Therapeutic Interventions in Atrial Fibrillation. J. Pers. Med. 2023, 13, 1237. https://doi.org/10.3390/jpm13081237
Baena-Montes JM, Kraśny MJ, O’Halloran M, Dunne E, Quinlan LR. In Vitro Models for Improved Therapeutic Interventions in Atrial Fibrillation. Journal of Personalized Medicine. 2023; 13(8):1237. https://doi.org/10.3390/jpm13081237
Chicago/Turabian StyleBaena-Montes, Jara M., Marcin J. Kraśny, Martin O’Halloran, Eoghan Dunne, and Leo R. Quinlan. 2023. "In Vitro Models for Improved Therapeutic Interventions in Atrial Fibrillation" Journal of Personalized Medicine 13, no. 8: 1237. https://doi.org/10.3390/jpm13081237
APA StyleBaena-Montes, J. M., Kraśny, M. J., O’Halloran, M., Dunne, E., & Quinlan, L. R. (2023). In Vitro Models for Improved Therapeutic Interventions in Atrial Fibrillation. Journal of Personalized Medicine, 13(8), 1237. https://doi.org/10.3390/jpm13081237