
Mechanisms of Allergen Immunotherapy and Potential Biomarkers for Clinical Evaluation
 ,
,  ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
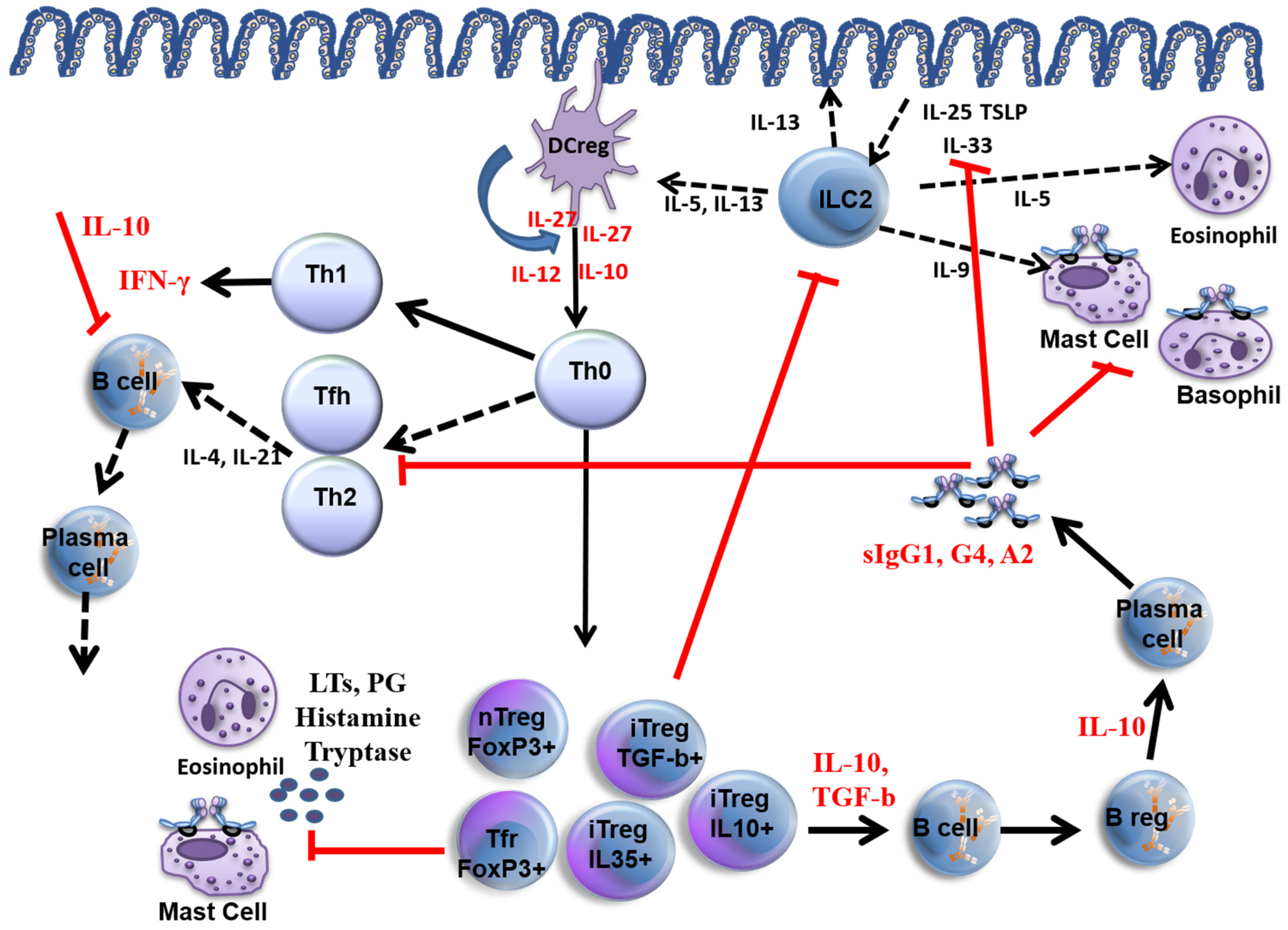
2. Mechanisms of AIT
2.1. Antibody Responses
2.2. Cellular Responses
2.3. Innate Lymphoid Cells and AIT
2.4. Histamine and Histamine Receptors
3. Biomarkers for AIT

{kind=link}
| Possible Biomarkers and Examples | Advantage | Limitation | References |
|---|---|---|---|
Specific IgE sIgE/tIgE | -Easy test to run, serum-based laboratory test -sIgE/tIgE seems to be reliable and promising marker to show clinical response | -sIgE rising at the beginning of AIT and not associated with the clinical response -sIgE/tIgE not validated yet. -Need for equivalence studies between tIgE units and sIgE units | [12,21,45,46,48,49,92,93,94,95,96] |
sIgG4 sIgE/sIgG4 | -Easy test to run, serum-based laboratory test -sIgG4 informative for allergen exposure -Currently, there is a commercial kit for sIgG4 | -It is not clear whether the sIgG4 levels and symptom and medication scores associate well. Need for further studies -Insufficient data on other IgG subsets -Limited data on local antibody levels -Low sIgG4 levels is a difficulty for measurement | [12,13,18,19,20,21,47,48,60,68,97,98,99,100] |
IgE-BF ELIFAB | -Serum-based laboratory test -High reproducibility is an advantage for IgE-FAB -IgE-FAB and IgE-BF associated with the clinical responses and reported in previous studies | -IgE-BF is not commercially available -It is not clear if IgE-FAB discriminates good responders -Despite the association between clinic response and IgE-FAB the number of studies is limited | [11,12,13,16,21,55,56,57,59,60,61,101,102,103,104] |
CD203c CD107 Diamine Oxidase | -Shows basophil activation with FcεRI mediated in-vivo response -Small amount of blood needed | -Test variability between different centers -Difficult technique -Lack of dose-response curves -Unresponsive basophils in some people | [11,60,63,64,105,106,107,108,109,110,111,112] |
IL-2R/IL-2 IL-4, IL-5, IL-9, IL-13, IL-17 IFN-γ, IL-12 TGF-β, IL-10 CCR3 TARC Tryptase | -Useful to understand the mechanisms of AIT -Local cytokine production may relate more to clinical manifestations | -There is no cytokine, chemokine or molecules identified to predict clinical response until now -Results may differ between centers and studies. -Specific T cell originated cytokine levels may be very low to be determined | [71,72,73,74,76,113] |
B regs Dendritic cells ILCs | -Tregs are very important in tolerance development -Bregs take very important roles in the mechanism of AIT -Treg and Breg cells may be more useful for drug development | -Technical difficulties -No data to show the association between Tregs and clinical response -Tregs appear very early in AIT so difficult to be a biomarker -Very low frequency of Tregs and Bregs | [7,78,79] |
Intradermal tests Conjunctival provocation tests Nasal Provocation tests Bronchial provocation tests Environmental Challenge Chambers (ECC) Food challenges Sting Challenges Allergen Challenges | -Standardised environmental factors -Seasonal pollen variations may be avoided -Surrogate markers of clinical response to AIT -ECC decrease variability in clinical studies, and they allow dose-response studies. -Food challenges help to find threshold levels for safe consumption -Early sting challenges are fairly reliable | -Mimics natural exposure but not the same -Lack of standardization for some allergen challenges -Cost of ECC is high, some reproducibility problems exist -Food challenges sometimes may be time-consuming, resource-intensive, and carries the risk of serious allergic reactions, including anaphylaxis -As time passes after treatment cessation sting challenges are poor to demonstrate relapse | [4,79,81,82,83,85,86,87,88,89,90] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pfaar, O.; Bousquet, J.; Durham, S.R.; Kleine-Tebbe, J.; Larche, M.; Roberts, G.; Shamji, M.H.; Gerth van Wijk, R. One hundred and ten years of Allergen Immunotherapy: A journey from empiric observation to evidence. Allergy 2022, 77, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Hise, K.; Rabin, R.L. Oral Immunotherapy for Food Allergy-a US Regulatory Perspective. Curr. Allergy Asthma Rep. 2020, 20, 77. [Google Scholar] [CrossRef]
- Alvaro-Lozano, M.; Akdis, C.A.; Akdis, M.; Alviani, C.; Angier, E.; Arasi, S.; Arzt-Gradwohl, L.; Barber, D.; Bazire, R.; Cavkaytar, O.; et al. Allergen Immunotherapy User’s Guide. Pediatr. Allergy Immunol. 2020, 31 (Suppl. 25), 1–101. [Google Scholar] [CrossRef] [PubMed]
- Sturm, G.J.; Varga, E.M.; Roberts, G.; Mosbech, H.; Bilo, M.B.; Akdis, C.A.; Antolín-Amérigo, D.; Cichocka-Jarosz, E.; Gawlik, R.; Jakob, T.; et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy 2018, 73, 744–764. [Google Scholar] [CrossRef] [PubMed]
- Dhami, S.; Nurmatov, U.; Arasi, S.; Khan, T.; Asaria, M.; Zaman, H.; Agarwal, A.; Netuveli, G.; Roberts, G.; Pfaar, O.; et al. Allergen immunotherapy for allergic rhinoconjunctivitis: A systematic review and meta-analysis. Allergy 2017, 72, 1597–1631. [Google Scholar] [CrossRef] [PubMed]
- Incorvaia, C.; Ridolo, E.; Mauro, M.; Pucciarini, F.; Heffler, E.; Canonica, G.W. Venom Immunotherapy and Aeroallergen Immunotherapy: How Do Their Outcomes Differ? Front. Allergy 2022, 3, 854080. [Google Scholar] [CrossRef]
- Shamji, M.H.; Kappen, J.H.; Akdis, M.; Jensen-Jarolim, E.; Knol, E.F.; Kleine-Tebbe, J.; Bohle, B.; Chaker, A.M.; Till, S.J.; Valenta, R.; et al. Biomarkers for monitoring clinical efficacy of allergen immunotherapy for allergic rhinoconjunctivitis and allergic asthma: An EAACI Position Paper. Allergy 2017, 72, 1156–1173. [Google Scholar] [CrossRef]
- Shamji, M.H.; Durham, S.R. Mechanisms of allergen immunotherapy for inhaled allergens and predictive biomarkers. J. Allergy Clin. Immunol. 2017, 140, 1485–1498. [Google Scholar] [CrossRef]
- Burks, A.W.; Calderon, M.A.; Casale, T.; Cox, L.; Demoly, P.; Jutel, M.; Nelson, H.; Akdis, C.A. Update on allergy immunotherapy: American Academy of Allergy, Asthma & Immunology/European Academy of Allergy and Clinical Immunology/PRACTALL consensus report. J. Allergy Clin. Immunol. 2013, 131, 1288–1296.e3. [Google Scholar]
- Cox, L.; Nelson, H.; Lockey, R.; Calabria, C.; Chacko, T.; Finegold, I.; Nelson, M.; Weber, R.; Bernstein, D.I.; Blessing-Moore, J.; et al. Allergen immunotherapy: A practice parameter third update. J. Allergy Clin. Immunol. 2011, 127, S1–S55. [Google Scholar] [CrossRef]
- Shamji, M.H.; Layhadi, J.A.; Scadding, G.W.; Cheung, D.K.; Calderon, M.A.; Turka, L.A.; Phippard, D.; Durham, S.R. Basophil expression of diamine oxidase: A novel biomarker of allergen immunotherapy response. J. Allergy Clin. Immunol. 2015, 135, 913–921.e9. [Google Scholar] [CrossRef] [PubMed]
- Didier, A.; Malling, H.J.; Worm, M.; Horak, F.; Jager, S.; Montagut, A.; Andre, C.; de Beaumont, O.; Melac, M. Optimal dose, efficacy, and safety of once-daily sublingual immunotherapy with a 5-grass pollen tablet for seasonal allergic rhinitis. J. Allergy Clin. Immunol. 2007, 120, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Dahl, R.; Kapp, A.; Colombo, G.; de Monchy, J.G.; Rak, S.; Emminger, W.; Riis, B.; Grønager, P.M.; Durham, S.R. Sublingual grass allergen tablet immunotherapy provides sustained clinical benefit with progressive immunologic changes over 2 years. J. Allergy Clin. Immunol. 2008, 121, 512–518.e2. [Google Scholar] [CrossRef] [PubMed]
- Huoman, J.; Papapavlou, G.; Pap, A.; Alm, J.; Nilsson, L.J.; Jenmalm, M.C. Sublingual immunotherapy alters salivary IgA and systemic immune mediators in timothy allergic children. Pediatr. Allergy Immunol. 2019, 30, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.H.; Larson, D.; Eifan, A.; Scadding, G.W.; Qin, T.; Lawson, K.; Sever, M.L.; Macfarlane, E.; Layhadi, J.A.; Würtzen, P.A.; et al. Differential induction of allergen-specific IgA responses following timothy grass subcutaneous and sublingual immunotherapy. J. Allergy Clin. Immunol. 2021, 148, 1061–1071.e11. [Google Scholar] [CrossRef]
- Platts-Mills, T.A.; von Maur, R.K.; Ishizaka, K.; Norman, P.S.; Lichtenstein, L.M. IgA and IgG anti-ragweed antibodies in nasal secretions. Quantitative measurements of antibodies and correlation with inhibition of histamine release. J. Clin. Investig. 1976, 57, 1041–1050. [Google Scholar] [CrossRef]
- Reisinger, J.; Horak, F.; Pauli, G.; van Hage, M.; Cromwell, O.; Konig, F.; Valenta, R.; Niederberger, V. Allergen-specific nasal IgG antibodies induced by vaccination with genetically modified allergens are associated with reduced nasal allergen sensitivity. J. Allergy Clin. Immunol. 2005, 116, 347–354. [Google Scholar] [CrossRef]
- La Rosa, M.; Ranno, C.; Andre, C.; Carat, F.; Tosca, M.A.; Canonica, G.W. Double-blind placebo-controlled evaluation of sublingual-swallow immunotherapy with standardized Parietaria judaica extract in children with allergic rhinoconjunctivitis. J. Allergy Clin. Immunol. 1999, 104, 425–432. [Google Scholar] [CrossRef]
- Troise, C.; Voltolini, S.; Canessa, A.; Pecora, S.; Negrini, A.C. Sublingual immunotherapy in Parietaria pollen-induced rhinitis: A double-blind study. J. Investig. Allergol. Clin. Immunol. 1995, 5, 25–30. [Google Scholar]
- Bahceciler, N.N.; Arikan, C.; Taylor, A.; Akdis, M.; Blaser, K.; Barlan, I.B.; Akdis, C.A. Impact of sublingual immunotherapy on specific antibody levels in asthmatic children allergic to house dust mites. Int. Arch. Allergy Immunol. 2005, 136, 287–294. [Google Scholar] [CrossRef]
- James, L.K.; Shamji, M.H.; Walker, S.M.; Wilson, D.R.; Wachholz, P.A.; Francis, J.N.; Jacobson, M.R.; Kimber, I.; Till, S.J.; Durham, S.R. Long-term tolerance after allergen immunotherapy is accompanied by selective persistence of blocking antibodies. J. Allergy Clin. Immunol. 2011, 127, 509–516.e5. [Google Scholar] [CrossRef] [PubMed]
- Sahiner, U.M.; Durham, S.R. Hymenoptera Venom Allergy: How Does Venom Immunotherapy Prevent Anaphylaxis From Bee and Wasp Stings? Front. Immunol. 2019, 10, 1959. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Bird, J.A.; Kulis, M.; Laubach, S.; Pons, L.; Shreffler, W.; Steele, P.; Kamilaris, J.; Vickery, B.; Burks, A.W. Sublingual immunotherapy for peanut allergy: Clinical and immunologic evidence of desensitization. J. Allergy Clin. Immunol. 2011, 127, 640–646.e1. [Google Scholar] [CrossRef] [PubMed]
- Vickery, B.P.; Scurlock, A.M.; Kulis, M.; Steele, P.H.; Kamilaris, J.; Berglund, J.P.; Burk, C.; Hiegel, A.; Carlisle, S.; Christie, L.; et al. Sustained unresponsiveness to peanut in subjects who have completed peanut oral immunotherapy. J. Allergy Clin. Immunol. 2014, 133, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Burks, A.W.; Keet, C.; Vickery, B.P.; Scurlock, A.M.; Wood, R.A.; Liu, A.H.; Sicherer, S.H.; Henning, A.K.; Lindblad, R.W.; et al. Long-term treatment with egg oral immunotherapy enhances sustained unresponsiveness that persists after cessation of therapy. J. Allergy Clin. Immunol. 2016, 137, 1117–1127.e10. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Jones, S.M.; Scurlock, A.M.; Perry, T.T.; Kemper, A.; Steele, P.; Hiegel, A.; Kamilaris, J.; Carlisle, S.; Yue, X.; et al. A randomized controlled study of peanut oral immunotherapy: Clinical desensitization and modulation of the allergic response. J. Allergy Clin. Immunol. 2011, 127, 654–660. [Google Scholar] [CrossRef]
- Baloh, C.H.; Huffaker, M.F.; Laidlaw, T. Biomarkers and mechanisms of tolerance induction in food allergic patients drive new therapeutic approaches. Front. Immunol. 2022, 13, 972103. [Google Scholar] [CrossRef]
- Tsai, M.; Mukai, K.; Chinthrajah, R.S.; Nadeau, K.C.; Galli, S.J. Sustained successful peanut oral immunotherapy associated with low basophil activation and peanut-specific IgE. J. Allergy Clin. Immunol. 2020, 145, 885–896.e6. [Google Scholar] [CrossRef]
- Plewako, H.; Wosinska, K.; Arvidsson, M.; Bjorkander, J.; Skov, P.S.; Hakansson, L.; Rak, S. Basophil interleukin 4 and interleukin 13 production is suppressed during the early phase of rush immunotherapy. Int. Arch. Allergy Immunol. 2006, 141, 346–353. [Google Scholar] [CrossRef]
- Dugas-Breit, S.; Przybilla, B.; Dugas, M.; Arnold, A.; Pfundstein, G.; Kuchenhoff, H.; Ruëff, F. Serum concentration of baseline mast cell tryptase: Evidence for a decline during long-term immunotherapy for Hymenoptera venom allergy. Clin. Exp. Allergy 2010, 40, 643–649. [Google Scholar] [CrossRef]
- Kulis, M.; Yue, X.; Guo, R.; Zhang, H.; Orgel, K.; Ye, P.; Li, Q.; Liu, Y.; Kim, E.; Burks, A.W.; et al. High- and low-dose oral immunotherapy similarly suppress pro-allergic cytokines and basophil activation in young children. Clin. Exp. Allergy 2019, 49, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Sicherer, S.H.; Burks, A.W.; Leung, D.Y.; Lindblad, R.W.; Dawson, P.; Henning, A.K.; Berin, M.C.; Chiang, D.; Vickery, B.P.; et al. Epicutaneous immunotherapy for the treatment of peanut allergy in children and young adults. J. Allergy Clin. Immunol. 2017, 139, 1242–1252.e9. [Google Scholar] [CrossRef] [PubMed]
- Keet, C.A.; Frischmeyer-Guerrerio, P.A.; Thyagarajan, A.; Schroeder, J.T.; Hamilton, R.G.; Boden, S.; Steele, P.; Driggers, S.; Burks, A.W.; Wood, R.A. The safety and efficacy of sublingual and oral immunotherapy for milk allergy. J. Allergy Clin. Immunol. 2012, 129, 448–455.e5. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.U.; Steinbrecher, J.; Calatroni, A.; Smith, N.; Ma, A.; Ruiter, B.; Virkud, Y.; Schneider, M.; Shreffler, W.G. Early decrease in basophil sensitivity to Ara h 2 precedes sustained unresponsiveness after peanut oral immunotherapy. J. Allergy Clin. Immunol. 2019, 144, 1310–1319.e4. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, M.; Narisety, S.D.; Guerrerio, A.L.; Chichester, K.L.; Keet, C.A.; Bieneman, A.P.; Hamilton, R.G.; Wood, R.A.; Schroeder, J.T.; Frischmeyer-Guerrerio, P.A. Suppression of the immunologic response to peanut during immunotherapy is often transient. J. Allergy Clin. Immunol. 2015, 135, 1283–1292. [Google Scholar] [CrossRef]
- Jones, S.M.; Kim, E.H.; Nadeau, K.C.; Nowak-Wegrzyn, A.; Wood, R.A.; Sampson, H.A.; Scurlock, A.M.; Chinthrajah, S.; Wang, J.; Pesek, R.D.; et al. Efficacy and safety of oral immunotherapy in children aged 1–3 years with peanut allergy (the Immune Tolerance Network IMPACT trial): A randomised placebo-controlled study. Lancet 2022, 399, 359–371. [Google Scholar] [CrossRef]
- Jutel, M.; Agache, I.; Bonini, S.; Burks, A.W.; Calderon, M.; Canonica, W.; Cox, L.; Demoly, P.; Frew, A.J.; O’Hehir, R.; et al. International Consensus on Allergen Immunotherapy II: Mechanisms, standardization, and pharmacoeconomics. J. Allergy Clin. Immunol. 2016, 137, 358–368. [Google Scholar] [CrossRef]
- Akdis, C.A.; Blesken, T.; Akdis, M.; Wuthrich, B.; Blaser, K. Role of interleukin 10 in specific immunotherapy. J. Clin. Investig. 1998, 102, 98–106. [Google Scholar] [CrossRef]
- Nasser, S.M.; Ying, S.; Meng, Q.; Kay, A.B.; Ewan, P.W. Interleukin-10 levels increase in cutaneous biopsies of patients undergoing wasp venom immunotherapy. Eur. J. Immunol. 2001, 31, 3704–3713. [Google Scholar] [CrossRef]
- Patil, S.U.; Ogunniyi, A.O.; Calatroni, A.; Tadigotla, V.R.; Ruiter, B.; Ma, A.; Moon, J.; Love, J.C.; Shreffler, W.G. Peanut oral immunotherapy transiently expands circulating Ara h 2-specific B cells with a homologous repertoire in unrelated subjects. J Allergy Clin. Immunol. 2015, 136, 125–134.e12. [Google Scholar] [CrossRef]
- Hoh, R.A.; Joshi, S.A.; Liu, Y.; Wang, C.; Roskin, K.M.; Lee, J.Y.; Pham, T.; Looney, T.J.; Jackson, K.J.; Dixit, V.P.; et al. Single B-cell deconvolution of peanut-specific antibody responses in allergic patients. J. Allergy Clin. Immunol. 2016, 137, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Komlosi, Z.I.; Kovacs, N.; Sokolowska, M.; van de Veen, W.; Akdis, M.; Akdis, C.A. Mechanisms of Subcutaneous and Sublingual Aeroallergen Immunotherapy: What is New? Immunol. Allergy Clin. N Am. 2020, 40, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Golebski, K.; Layhadi, J.A.; Sahiner, U.; Steveling-Klein, E.H.; Lenormand, M.M.; Li, R.C.Y.; Bal, S.M.; Heesters, B.A.; Vilà-Nadal, G.; Hunewald, O.; et al. Induction of IL-10-producing type 2 innate lymphoid cells by allergen immunotherapy is associated with clinical response. Immunity 2021, 54, 291–307.e7. [Google Scholar] [CrossRef] [PubMed]
- Globinska, A.; Boonpiyathad, T.; Satitsuksanoa, P.; Kleuskens, M.; van de Veen, W.; Sokolowska, M.; Akdis, M. Mechanisms of allergen-specific immunotherapy: Diverse mechanisms of immune tolerance to allergens. Ann. Allergy Asthma Immunol. 2018, 121, 306–312. [Google Scholar] [CrossRef]
- Keskin, O.; Tuncer, A.; Adalioglu, G.; Sekerel, B.E.; Sackesen, C.; Kalayci, O. The effects of grass pollen allergoid immunotherapy on clinical and immunological parameters in children with allergic rhinitis. Pediatr. Allergy Immunol. 2006, 17, 396–407. [Google Scholar] [CrossRef]
- Durham, S.R.; Yang, W.H.; Pedersen, M.R.; Johansen, N.; Rak, S. Sublingual immunotherapy with once-daily grass allergen tablets: A randomized controlled trial in seasonal allergic rhinoconjunctivitis. J. Allergy Clin. Immunol. 2006, 117, 802–809. [Google Scholar] [CrossRef]
- Gehlhar, K.; Schlaak, M.; Becker, W.; Bufe, A. Monitoring allergen immunotherapy of pollen-allergic patients: The ratio of allergen-specific IgG4 to IgG1 correlates with clinical outcome. Clin. Exp. Allergy 1999, 29, 497–506. [Google Scholar] [CrossRef]
- Rolinck-Werninghaus, C.; Kopp, M.; Liebke, C.; Lange, J.; Wahn, U.; Niggemann, B. Lack of detectable alterations in immune responses during sublingual immunotherapy in children with seasonal allergic rhinoconjunctivitis to grass pollen. Int. Arch. Allergy Immunol. 2005, 136, 134–141. [Google Scholar] [CrossRef]
- Di Lorenzo, G.; Mansueto, P.; Pacor, M.L.; Rizzo, M.; Castello, F.; Martinelli, N.; Ditta, V.; Bianco, C.L.; Leto-Barone, M.S.; D’Alcamo, A.; et al. Evaluation of serum s-IgE/total IgE ratio in predicting clinical response to allergen-specific immunotherapy. J. Allergy Clin. Immunol. 2009, 123, 1103–1110.e4. [Google Scholar] [CrossRef]
- Shamji, M.H.; Ljorring, C.; Francis, J.N.; Calderon, M.A.; Larche, M.; Kimber, I.; Frew, A.J.; Ipsen, H.; Lund, K.; Würtzen, P.A.; et al. Functional rather than immunoreactive levels of IgG4 correlate closely with clinical response to grass pollen immunotherapy. Allergy 2012, 67, 217–226. [Google Scholar] [CrossRef]
- Sanchez-Ruano, L.; de la Hoz, B.; Martinez-Botas, J. Clinical utility of microarray B-cell epitope mapping in food allergies: A systematic review. Pediatr. Allergy Immunol. 2020, 31, 175–185. [Google Scholar] [CrossRef]
- Vickery, B.P.; Lin, J.; Kulis, M.; Fu, Z.; Steele, P.H.; Jones, S.M.; Scurlock, A.M.; Gimenez, G.; Bardina, L.; Sampson, H.A.; et al. Peanut oral immunotherapy modifies IgE and IgG4 responses to major peanut allergens. J. Allergy Clin. Immunol. 2013, 131, 128–134.e3. [Google Scholar] [CrossRef] [PubMed]
- Savilahti, E.M.; Kuitunen, M.; Valori, M.; Rantanen, V.; Bardina, L.; Gimenez, G.; Makela, M.J.; Hautaniemi, S.; Savilahti, E.; Sampson, H.A. Use of IgE and IgG4 epitope binding to predict the outcome of oral immunotherapy in cow’s milk allergy. Pediatr. Allergy Immunol. 2014, 25, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Botas, J.; Rodriguez-Alvarez, M.; Cerecedo, I.; Vlaicu, C.; Dieguez, M.C.; Gomez-Coronado, D.; Fernandez-Rivas, M.; de la Hoz, B. Identification of novel peptide biomarkers to predict safety and efficacy of cow’s milk oral immunotherapy by peptide microarray. Clin. Exp. Allergy 2015, 45, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.H.; Wilcock, L.K.; Wachholz, P.A.; Dearman, R.J.; Kimber, I.; Wurtzen, P.A.; Larché, M.; Durham, S.R.; Francis, J.N. The IgE-facilitated allergen binding (FAB) assay: Validation of a novel flow-cytometric based method for the detection of inhibitory antibody responses. J. Immunol. Methods 2006, 317, 71–79. [Google Scholar] [CrossRef]
- Shamji, M.H.; Francis, J.N.; Wurtzen, P.A.; Lund, K.; Durham, S.R.; Till, S.J. Cell-free detection of allergen-IgE cross-linking with immobilized phase CD23: Inhibition by blocking antibody responses after immunotherapy. J. Allergy Clin. Immunol. 2013, 132, 1003–1005.e4. [Google Scholar] [CrossRef]
- Petersen, A.B.; Gudmann, P.; Milvang-Gronager, P.; Morkeberg, R.; Bogestrand, S.; Linneberg, A.; Johansen, N. Performance evaluation of a specific IgE assay developed for the ADVIA centaur immunoassay system. Clin. Biochem. 2004, 37, 882–892. [Google Scholar] [CrossRef]
- Blaiss, M.; Maloney, J.; Nolte, H.; Gawchik, S.; Yao, R.; Skoner, D.P. Efficacy and safety of timothy grass allergy immunotherapy tablets in North American children and adolescents. J. Allergy Clin. Immunol. 2011, 127, 64–71.e4. [Google Scholar] [CrossRef]
- Corzo, J.L.; Carrillo, T.; Pedemonte, C.; Plaza Martin, A.M.; Martin Hurtado, S.; Dige, E.; Calderon, M.A. Tolerability during double-blind randomized phase I trials with the house dust mite allergy immunotherapy tablet in adults and children. J. Investig. Allergol. Clin. Immunol. 2014, 24, 154–161. [Google Scholar]
- Wachholz, P.A.; Soni, N.K.; Till, S.J.; Durham, S.R. Inhibition of allergen-IgE binding to B cells by IgG antibodies after grass pollen immunotherapy. J. Allergy Clin. Immunol. 2003, 112, 915–922. [Google Scholar] [CrossRef]
- van Neerven, R.J.; Wikborg, T.; Lund, G.; Jacobsen, B.; Brinch-Nielsen, A.; Arnved, J.; Ipsen, H. Blocking antibodies induced by specific allergy vaccination prevent the activation of CD4+ T cells by inhibiting serum-IgE-facilitated allergen presentation. J. Immunol. 1999, 163, 2944–2952. [Google Scholar] [CrossRef] [PubMed]
- Kepley, C.L.; Cambier, J.C.; Morel, P.A.; Lujan, D.; Ortega, E.; Wilson, B.S.; Oliver, J.M. Negative regulation of FcepsilonRI signaling by FcgammaRII costimulation in human blood basophils. J. Allergy Clin. Immunol. 2000, 106, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Ebo, D.G.; Bridts, C.H.; Mertens, C.H.; Hagendorens, M.M.; Stevens, W.J.; De Clerck, L.S. Analyzing histamine release by flow cytometry (HistaFlow): A novel instrument to study the degranulation patterns of basophils. J. Immunol. Methods 2012, 375, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Nullens, S.; Sabato, V.; Faber, M.; Leysen, J.; Bridts, C.H.; De Clerck, L.S.; Falcone, F.H.; Maurer, M.; Ebo, D.G. Basophilic histamine content and release during venom immunotherapy: Insights by flow cytometry. Cytom. B Clin. Cytom. 2013, 84, 173–178. [Google Scholar] [CrossRef]
- Cosmi, L.; Santarlasci, V.; Angeli, R.; Liotta, F.; Maggi, L.; Frosali, F.; Rossi, O.; Falagiani, P.; Riva, G.; Romagnani, S.; et al. Sublingual immunotherapy with Dermatophagoides monomeric allergoid down-regulates allergen-specific immunoglobulin E and increases both interferon-gamma- and interleukin-10-production. Clin. Exp. Allergy 2006, 36, 261–272. [Google Scholar] [CrossRef]
- Fanta, C.; Bohle, B.; Hirt, W.; Siemann, U.; Horak, F.; Kraft, D.; Ebner, H.; Ebner, C. Systemic immunological changes induced by administration of grass pollen allergens via the oral mucosa during sublingual immunotherapy. Int. Arch. Allergy Immunol. 1999, 120, 218–224. [Google Scholar] [CrossRef]
- Jutel, M.; Akdis, M.; Budak, F.; Aebischer-Casaulta, C.; Wrzyszcz, M.; Blaser, K.; Akdis, C.A. IL-10 and TGF-beta cooperate in the regulatory T cell response to mucosal allergens in normal immunity and specific immunotherapy. Eur. J. Immunol. 2003, 33, 1205–1214. [Google Scholar] [CrossRef]
- Bohle, B.; Kinaciyan, T.; Gerstmayr, M.; Radakovics, A.; Jahn-Schmid, B.; Ebner, C. Sublingual immunotherapy induces IL-10-producing T regulatory cells, allergen-specific T-cell tolerance, and immune deviation. J. Allergy Clin. Immunol. 2007, 120, 707–713. [Google Scholar] [CrossRef]
- Francis, J.N.; Till, S.J.; Durham, S.R. Induction of IL-10+CD4+CD25+ T cells by grass pollen immunotherapy. J. Allergy Clin. Immunol. 2003, 111, 1255–1261. [Google Scholar] [CrossRef]
- Wachholz, P.A.; Nouri-Aria, K.T.; Wilson, D.R.; Walker, S.M.; Verhoef, A.; Till, S.J.; Durham, S.R. Grass pollen immunotherapy for hayfever is associated with increases in local nasal but not peripheral Th1:Th2 cytokine ratios. Immunology 2002, 105, 56–62. [Google Scholar] [CrossRef]
- Plewako, H.; Holmberg, K.; Oancea, I.; Gotlib, T.; Samolinski, B.; Rak, S. A follow-up study of immunotherapy-treated birch-allergic patients: Effect on the expression of chemokines in the nasal mucosa. Clin. Exp. Allergy. 2008, 38, 1124–1131. [Google Scholar] [CrossRef]
- Makino, Y.; Noguchi, E.; Takahashi, N.; Matsumoto, Y.; Kubo, S.; Yamada, T.; Imoto, Y.; Ito, Y.; Osawa, Y.; Shibasaki, M.; et al. Apolipoprotein A-IV is a candidate target molecule for the treatment of seasonal allergic rhinitis. J. Allergy Clin. Immunol. 2010, 126, 1163–1169.e5. [Google Scholar] [CrossRef]
- Salmivesi, S.; Paassilta, M.; Huhtala, H.; Nieminen, R.; Moilanen, E.; Korppi, M. Changes in biomarkers during a six-month oral immunotherapy intervention for cow’s milk allergy. Acta Paediatr. 2016, 105, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, E.; He, M. Cytokine Responses to Specific Immunotherapy in House Dust Mite-Induced Allergic Rhinitis Patients. Inflammation 2015, 38, 2216–2223. [Google Scholar] [CrossRef] [PubMed]
- Scadding, G.W.; Eifan, A.O.; Lao-Araya, M.; Penagos, M.; Poon, S.Y.; Steveling, E.; Yan, R.; Switzer, A.; Phippard, D.; Togias, A.; et al. Effect of grass pollen immunotherapy on clinical and local immune response to nasal allergen challenge. Allergy 2015, 70, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; De Amici, M.; Murdaca, G.; Filaci, G.; Fenoglio, D.; Marseglia, G.L. Adipokines and sublingual immunotherapy: Preliminary report. Hum. Immunol. 2009, 70, 73–78. [Google Scholar] [CrossRef]
- Ciprandi, G.; Continia, P.; Fenoglio, D.; Sormani, M.P.; Negrini, S.; Puppo, F.; Indiveri, F. Relationship between soluble HLA-G and HLA-A,-B,-C serum levels, and interferon-gamma production after sublingual immunotherapy in patients with allergic rhinitis. Hum. Immunol. 2008, 69, 409–413. [Google Scholar] [CrossRef] [PubMed]
- van de Veen, W. The role of regulatory B cells in allergen immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 447–452. [Google Scholar] [CrossRef]
- Sindher, S.B.; Long, A.; Acharya, S.; Sampath, V.; Nadeau, K.C. The Use of Biomarkers to Predict Aero-Allergen and Food Immunotherapy Responses. Clin. Rev. Allergy Immunol. 2018, 55, 190–204. [Google Scholar] [CrossRef]
- Rosner-Friese, K.; Kaul, S.; Vieths, S.; Pfaar, O. Environmental exposure chambers in allergen immunotherapy trials: Current status and clinical validation needs. J. Allergy Clin. Immunol. 2015, 135, 636–643. [Google Scholar] [CrossRef]
- Zieglmayer, P.U.; Pfaar, O. Update on the use of allergen challenge chambers in immunotherapy: Clinical implications. Curr. Opin. Allergy Clin. Immunol. 2022, 22, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Pfaar, O.; Bergmann, K.C.; Bonini, S.; Compalati, E.; Domis, N.; de Blay, F.; de Kam, P.; Devillier, P.; Durham, S.R.; Ellis, A.K.; et al. Technical standards in allergen exposure chambers worldwide—An EAACI Task Force Report. Allergy 2021, 76, 3589–3612. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Leon, B.; Martinez San Ireneo, M.; de la Roca, F.; Arenas, L.; Alfaya Arias, T.; Cordobes, C.; Marqués, L.; Vega, A.; Moreno-Aguila, C. The Lights and the Shadows of Controlled Sting Challenge With Hymenoptera. J. Investig. Allergol. Clin. Immunol. 2022, 32, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, R.; Kemeny, D.M.; Richards, D. Sub-class of IgG anti-bee venom antibody produced during bee venom immunotherapy and its relationship to long-term protection from bee stings and following termination of venom immunotherapy. Clin. Allergy 1986, 16, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Golden, D.B.; Kwiterovich, K.A.; Kagey-Sobotka, A.; Valentine, M.D.; Lichtenstein, L.M. Discontinuing venom immunotherapy: Outcome after five years. J. Allergy Clin. Immunol. 1996, 97, 579–587. [Google Scholar] [CrossRef] [PubMed]
- van Halteren, H.K.; van der Linden, P.W.; Burgers, J.A.; Bartelink, A.K. Discontinuation of yellow jacket venom immunotherapy: Follow-up of 75 patients by means of deliberate sting challenge. J. Allergy Clin. Immunol. 1997, 100, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Teufel, M.; Feidt, A.; Giel, K.E.; Zipfel, S.; Biedermann, T. Tolerated wasp sting challenge improves health-related quality of life in patients allergic to wasp venom. J. Allergy Clin. Immunol. 2013, 132, 489–490. [Google Scholar] [CrossRef]
- Swamy, R.S.; Reshamwala, N.; Hunter, T.; Vissamsetti, S.; Santos, C.B.; Baroody, F.M.; Hwang, P.H.; Hoyte, E.G.; Garcia, M.A.; Nadeau, K.C. Epigenetic modifications and improved regulatory T-cell function in subjects undergoing dual sublingual immunotherapy. J. Allergy Clin. Immunol. 2012, 130, 215–224.e7. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.Y.; Pan, C.; Ma, T.T.; Chen, Y.L.; Yan, W.J.; Liu, J.G.; Cao, M.-D.; Huang, H.-D.; Wang, D.-Y.; Wang, X.-Y.; et al. Clinical Efficacy Evaluation of 1-Year Subcutaneous Immunotherapy for Artemisia sieversiana Pollen Allergic Rhinitis by Serum Metabolomics. Front. Pharmacol. 2020, 11, 305. [Google Scholar] [CrossRef]
- Zheng, P.; Bian, X.; Zhai, Y.; Li, C.; Li, N.; Hao, C.; Huang, H.; Luo, W.; Huang, Z.; Liao, C.; et al. Metabolomics reveals a correlation between hydroxyeicosatetraenoic acids and allergic asthma: Evidence from three years’ immunotherapy. Pediatr. Allergy Immunol. 2021, 32, 1654–1662. [Google Scholar] [CrossRef]
- Pfaar, O.; Bonini, S.; Cardona, V.; Demoly, P.; Jakob, T.; Jutel, M.; Kleine-Tebbe, J.; Klimek, L.; Klysner, S.; Kopp, M.V.; et al. Perspectives in allergen immunotherapy: 2017 and beyond. Allergy 2018, 73 (Suppl. 104), 5–23. [Google Scholar] [CrossRef] [PubMed]
- Durham, S.R.; Walker, S.M.; Varga, E.M.; Jacobson, M.R.; O’Brien, F.; Noble, W.; Till, S.J.; Hamid, Q.A.; Nouri-Aria, K.T. Long-term clinical efficacy of grass-pollen immunotherapy. N. Engl. J. Med. 1999, 341, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Pilette, C.; Nouri-Aria, K.T.; Jacobson, M.R.; Wilcock, L.K.; Detry, B.; Walker, S.M.; Francis, J.N.; Durham, S.R. Grass pollen immunotherapy induces an allergen-specific IgA2 antibody response associated with mucosal TGF-beta expression. J. Immunol. 2007, 178, 4658–4666. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Yonekura, S.; Horiguchi, S.; Taniguchi, Y.; Saito, A.; Yasueda, H.; Inamine, A.; Nakayama, T.; Takemori, T.; Taniguchi, M.; et al. Increase of regulatory T cells and the ratio of specific IgE to total IgE are candidates for response monitoring or prognostic biomarkers in 2-year sublingual immunotherapy (SLIT) for Japanese cedar pollinosis. Clin. Immunol. 2011, 139, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Eifan, A.O.; Akkoc, T.; Yildiz, A.; Keles, S.; Ozdemir, C.; Bahceciler, N.N.; Barlan, I.B. Clinical efficacy and immunological mechanisms of sublingual and subcutaneous immunotherapy in asthmatic/rhinitis children sensitized to house dust mite: An open randomized controlled trial. Clin. Exp. Allergy 2010, 40, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, M.; Yue, W.; Zhou, J.; Li, R.; Lin, J.; Li, Y. Predictive factors for clinical response to allergy immunotherapy in children with asthma and rhinitis. Int. Arch. Allergy Immunol. 2014, 164, 210–217. [Google Scholar] [CrossRef]
- Gomez, E.; Fernandez, T.D.; Dona, I.; Rondon, C.; Campo, P.; Gomez, F.; Salas, M.; Gonzalez, M.; Perkins, J.R.; Palomares, F.; et al. Initial immunological changes as predictors for house dust mite immunotherapy response. Clin. Exp. Allergy 2015, 45, 1542–1553. [Google Scholar] [CrossRef] [PubMed]
- Moverare, R.; Elfman, L.; Vesterinen, E.; Metso, T.; Haahtela, T. Development of new IgE specificities to allergenic components in birch pollen extract during specific immunotherapy studied with immunoblotting and Pharmacia CAP System. Allergy 2002, 57, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.S.; Nolte, H.; Creticos, P.; Maloney, J.; Wu, J.; Bernstein, D.I. Efficacy and safety of timothy grass allergy immunotherapy tablet treatment in North American adults. J. Allergy Clin. Immunol. 2011, 127, 72–80.e2. [Google Scholar] [CrossRef]
- Wollmann, E.; Lupinek, C.; Kundi, M.; Selb, R.; Niederberger, V.; Valenta, R. Reduction in allergen-specific IgE binding as measured by microarray: A possible surrogate marker for effects of specific immunotherapy. J. Allergy Clin. Immunol. 2015, 136, 806–809.e7. [Google Scholar] [CrossRef]
- Nouri-Aria, K.T.; Wachholz, P.A.; Francis, J.N.; Jacobson, M.R.; Walker, S.M.; Wilcock, L.K.; Staple, S.Q.; Aalberse, R.C.; Till, S.J.; Durham, S.R. Grass pollen immunotherapy induces mucosal and peripheral IL-10 responses and blocking IgG activity. J. Immunol. 2004, 172, 3252–3559. [Google Scholar] [CrossRef] [PubMed]
- Durham, S.R.; Emminger, W.; Kapp, A.; de Monchy, J.G.; Rak, S.; Scadding, G.K.; Wurtzen, P.A.; Andersen, J.S.; Tholstrup, B.; Riis, B.; et al. SQ-standardized sublingual grass immunotherapy: Confirmation of disease modification 2 years after 3 years of treatment in a randomized trial. J. Allergy Clin. Immunol. 2012, 129, 717–725.e5. [Google Scholar] [CrossRef] [PubMed]
- Wurtzen, P.A.; Lund, G.; Lund, K.; Arvidsson, M.; Rak, S.; Ipsen, H. A double-blind placebo-controlled birch allergy vaccination study II: Correlation between inhibition of IgE binding, histamine release and facilitated allergen presentation. Clin. Exp. Allergy 2008, 38, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Reich, K.; Gessner, C.; Kroker, A.; Schwab, J.A.; Pohl, W.; Villesen, H.; Wüstenberg, E.; Emminger, W. Immunologic effects and tolerability profile of in-season initiation of a standardized-quality grass allergy immunotherapy tablet: A phase III, multicenter, randomized, double-blind, placebo-controlled trial in adults with grass pollen-induced rhinoconjunctivitis. Clin. Ther. 2011, 33, 828–840. [Google Scholar]
- Van Overtvelt, L.; Baron-Bodo, V.; Horiot, S.; Moussu, H.; Ricarte, C.; Horak, F.; Zieglmayer, P.; Zieglmayer, R.; Montagut, A.; Galvain, S.; et al. Changes in basophil activation during grass-pollen sublingual immunotherapy do not correlate with clinical efficacy. Allergy 2011, 66, 1530–1537. [Google Scholar] [CrossRef]
- Buhring, H.J.; Streble, A.; Valent, P. The basophil-specific ectoenzyme E-NPP3 (CD203c) as a marker for cell activation and allergy diagnosis. Int. Arch. Allergy Immunol. 2004, 133, 317–329. [Google Scholar] [CrossRef]
- Knol, E.F.; Mul, F.P.; Jansen, H.; Calafat, J.; Roos, D. Monitoring human basophil activation via CD63 monoclonal antibody 435. J. Allergy Clin. Immunol. 1991, 88, 328–338. [Google Scholar] [CrossRef]
- Hennersdorf, F.; Florian, S.; Jakob, A.; Baumgartner, K.; Sonneck, K.; Nordheim, A.; Biedermann, T.; Valent, P.; Bühring, H.-J. Identification of CD13, CD107a, and CD164 as novel basophil-activation markers and dissection of two response patterns in time kinetics of IgE-dependent upregulation. Cell Res. 2005, 15, 325–335. [Google Scholar] [CrossRef]
- Kepil Ozdemir, S.; Sin, B.A.; Guloglu, D.; Ikinciogullari, A.; Gencturk, Z.; Misirligil, Z. Short-term preseasonal immunotherapy: Is early clinical efficacy related to the basophil response? Int. Arch. Allergy Immunol. 2014, 164, 237–245. [Google Scholar] [CrossRef]
- Aasbjerg, K.; Backer, V.; Lund, G.; Holm, J.; Nielsen, N.C.; Holse, M.; Wagtmann, V.R.; Würtzen, P.A. Immunological comparison of allergen immunotherapy tablet treatment and subcutaneous immunotherapy against grass allergy. Clin. Exp. Allergy 2014, 44, 417–428. [Google Scholar] [CrossRef]
- Ceuppens, J.L.; Bullens, D.; Kleinjans, H.; van der Werf, J.; The PURETHAL Birch Efficacy Study Group. Immunotherapy with a modified birch pollen extract in allergic rhinoconjunctivitis: Clinical and immunological effects. Clin. Exp. Allergy 2009, 39, 1903–1909. [Google Scholar] [CrossRef] [PubMed]
- Gueguen, C.; Bouley, J.; Moussu, H.; Luce, S.; Duchateau, M.; Chamot-Rooke, J.; Pallardy, M.; Lombardi, V.; Nony, E.; Baron-Bodo, V.; et al. Changes in markers associated with dendritic cells driving the differentiation of either TH2 cells or regulatory T cells correlate with clinical benefit during allergen immunotherapy. J. Allergy Clin. Immunol. 2016, 137, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Scadding, G.W.; Calderon, M.A.; Shamji, M.H.; Eifan, A.O.; Penagos, M.; Dumitru, F.; Sever, M.L.; Bahnson, H.T.; Lawson, K.; Harris, K.M.; et al. Effect of 2 Years of Treatment With Sublingual Grass Pollen Immunotherapy on Nasal Response to Allergen Challenge at 3 Years among Patients With Moderate to Severe Seasonal Allergic Rhinitis: The GRASS Randomized Clinical Trial. JAMA 2017, 317, 615–625. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahiner, U.M.; Giovannini, M.; Escribese, M.M.; Paoletti, G.; Heffler, E.; Alvaro Lozano, M.; Barber, D.; Canonica, G.W.; Pfaar, O. Mechanisms of Allergen Immunotherapy and Potential Biomarkers for Clinical Evaluation. J. Pers. Med. 2023, 13, 845. https://doi.org/10.3390/jpm13050845
Sahiner UM, Giovannini M, Escribese MM, Paoletti G, Heffler E, Alvaro Lozano M, Barber D, Canonica GW, Pfaar O. Mechanisms of Allergen Immunotherapy and Potential Biomarkers for Clinical Evaluation. Journal of Personalized Medicine. 2023; 13(5):845. https://doi.org/10.3390/jpm13050845
Chicago/Turabian StyleSahiner, Umit M., Mattia Giovannini, Maria M. Escribese, Giovanni Paoletti, Enrico Heffler, Montserrat Alvaro Lozano, Domingo Barber, Giorgio Walter Canonica, and Oliver Pfaar. 2023. "Mechanisms of Allergen Immunotherapy and Potential Biomarkers for Clinical Evaluation" Journal of Personalized Medicine 13, no. 5: 845. https://doi.org/10.3390/jpm13050845
APA StyleSahiner, U. M., Giovannini, M., Escribese, M. M., Paoletti, G., Heffler, E., Alvaro Lozano, M., Barber, D., Canonica, G. W., & Pfaar, O. (2023). Mechanisms of Allergen Immunotherapy and Potential Biomarkers for Clinical Evaluation. Journal of Personalized Medicine, 13(5), 845. https://doi.org/10.3390/jpm13050845