
Three Pathways of Cancer Cachexia: Inflammation, Changes in Adipose Tissue and Loss of Muscle Mass—The Role of miRNAs

 , and
, and 
Abstract
:1. Introduction
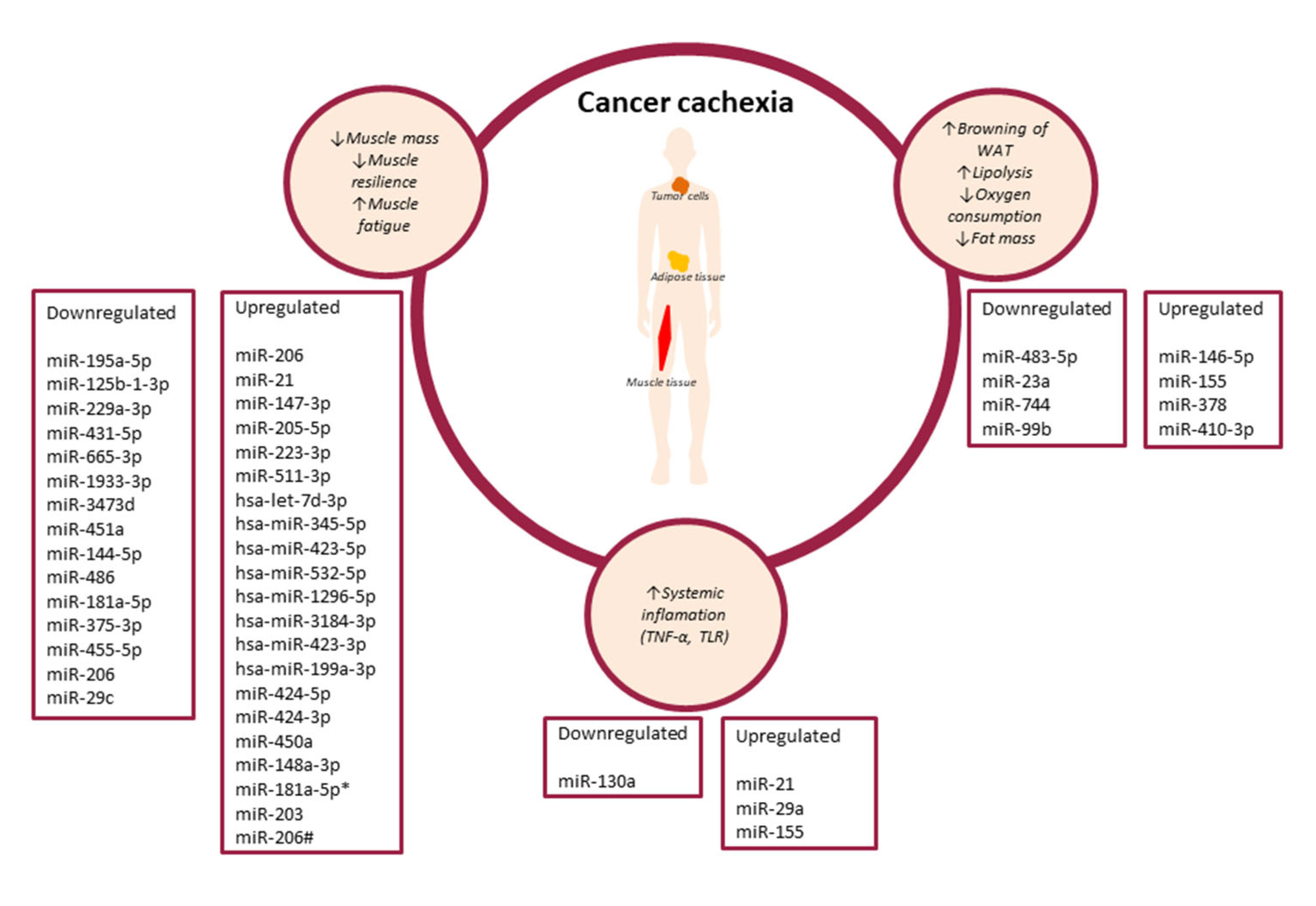
1.1. Cancer Cachexia
1.2. miRNA
1.2.1. The Role of miRNAs in the Regulation of Proinflammatory Cytokines Involved in Cancer Cachexia
1.2.2. Role of miRNAs in Cancer Cachexia-Related Adipose Tissue Remodelling
1.2.3. The Role of miRNAs in Cancer Cachexia-Related Muscle Tissue Remodelling
2. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Kottorou, A.; Dimitrakopoulos, F.I.; Tsezou, A. Non-coding RNAs in cancer-associated cachexia: Clinical implications and future perspectives. Transl. Oncol. 2021, 14, 101101. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet. Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Fearon, K.C.H.; Glass, D.J.; Guttridge, D.C. Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.D.; Smith, G.L.; Hurria, A.; Hortobagyi, G.N.; Buchholz, T.A. Future of cancer incidence in the United States: Burdens upon an aging, changing nation. J. Clin. Oncol. 2009, 27, 2758–2765. [Google Scholar] [CrossRef] [PubMed]
- Mattox, T.W. Cancer Cachexia: Cause, Diagnosis, and Treatment. Nutr. Clin. Pract. 2017, 32, 599–606. [Google Scholar] [CrossRef]
- Johns, N.; Stephens, N.A.; Fearon, K.C.H. Muscle wasting in cancer. Int. J. Biochem. Cell Biol. 2013, 45, 2215–2229. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M. Muscle wasting: The gut microbiota as a new therapeutic target? Int. J. Biochem. Cell Biol. 2013, 45, 2186–2190. [Google Scholar] [CrossRef]
- Loumaye, A.; Thissen, J.-P. Biomarkers of cancer cachexia. Clin. Biochem. 2017, 50, 1281–1288. [Google Scholar] [CrossRef]
- Fouladiun, M.; Körner, U.; Bosaeus, I.; Daneryd, P.; Hyltander, A.; Lundholm, K.G. Body composition and time course changes in regional distribution of fat and lean tissue in unselected cancer patients on palliative care--correlations with food intake, metabolism, exercise capacity, and hormones. Cancer 2005, 103, 2189–2198. [Google Scholar] [CrossRef]
- Hall, K.D.; Baracos, V.E. Computational modeling of cancer cachexia. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Brown, J.L.; Washington, T.A.; Greene, N.P. Development and progression of cancer cachexia: Perspectives from bench to bedside. Sport. Med. Health Sci. 2020, 2, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Porkka, K.P.; Pfeiffer, M.J.; Waltering, K.K.; Vessella, R.L.; Tammela, T.L.J.; Visakorpi, T. MicroRNA expression profiling in prostate cancer. Cancer Res. 2007, 67, 6130–6135. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Ding, Z.; Shen, L.; Yang, F.; Han, J.; Wu, G. miR-410-3P inhibits adipocyte differentiation by targeting IRS-1 in cancer-associated cachexia patients. Lipids Health Dis. 2021, 20, 115. [Google Scholar] [CrossRef]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Zhang, B.; Li, B.; Qin, F.; Bai, F.; Sun, C.; Liu, Q. Expression of serum microRNA-155 and its clinical importance in patients with heart failure after myocardial infarction. J. Int. Med. Res. 2019, 47, 6294–6302. [Google Scholar] [CrossRef]
- Shioya, M.; Obayashi, S.; Tabunoki, H.; Arima, K.; Saito, Y.; Ishida, T.; Satoh, J. Aberrant microRNA expression in the brains of neurodegenerative diseases: MiR-29a decreased in Alzheimer disease brains targets neurone navigator 3. Neuropathol. Appl. Neurobiol. 2010, 36, 320–330. [Google Scholar] [CrossRef]
- Ben-Dov, I.Z.; Tan, Y.-C.; Morozov, P.; Wilson, P.D.; Rennert, H.; Blumenfeld, J.D.; Tuschl, T. Urine microRNA as potential biomarkers of autosomal dominant polycystic kidney disease progression: Description of miRNA profiles at baseline. PLoS ONE 2014, 9, e86856. [Google Scholar] [CrossRef] [Green Version]
- Migdalska-Sęk, M.; Modrzewska, B.; Kordiak, J.; Pastuszak-Lewandoska, D.; Kiszałkiewicz, J.M.; Bielec, F.; Antczak, A.; Brzeziańska-Lasota, E. Diagnostic value of PPARδ and miRNA-17 expression levels in patients with non-small cell lung cancer. Sci. Rep. 2021, 11, 24136. [Google Scholar] [CrossRef] [PubMed]
- Hess, A.-K.; Müer, A.; Mairinger, F.D.; Weichert, W.; Stenzinger, A.; Hummel, M.; Budach, V.; Tinhofer, I. MiR-200b and miR-155 as predictive biomarkers for the efficacy of chemoradiation in locally advanced head and neck squamous cell carcinoma. Eur. J. Cancer 2017, 77, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Zhang, Q.; Zhao, S.; Wang, J.; Lu, K.; Song, Y.; Zhao, L.; Kang, X.; Wang, J.; Xu, S.; et al. The expression and clinical significance of microRNA-1258 and heparanase in human breast cancer. Clin. Biochem. 2013, 46, 926–932. [Google Scholar] [CrossRef]
- Cheng, H.H.; Plets, M.; Li, H.; Higano, C.S.; Tangen, C.M.; Agarwal, N.; Vogelzang, N.J.; Hussain, M.; Thompson, I.M.J.; Tewari, M.; et al. Circulating microRNAs and treatment response in the Phase II SWOG S0925 study for patients with new metastatic hormone-sensitive prostate cancer. Prostate 2018, 78, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhang, J.; Wang, J.; Lin, X.; Ding, F. Role of miRNA in Lung Cancer-Potential Biomarkers and Therapies. Curr. Pharm. Des. 2018, 23, 5997–6010. [Google Scholar] [CrossRef]
- Wang, L.Q.; Kumar, S.; Calin, G.A.; Li, Z.; Chim, C.S. Frequent methylation of the tumour suppressor miR-1258 targeting PDL1: Implication in multiple myeloma-specific cytotoxicity and prognostification. Br. J. Haematol. 2020, 190, 249–261. [Google Scholar] [CrossRef]
- Nagachinta, S.; Bouzo, B.L.; Vazquez-Rios, A.J.; Lopez, R.; Fuente, M. de la Sphingomyelin-Based Nanosystems (SNs) for the Development of Anticancer miRNA Therapeutics. Pharmaceutics 2020, 12, 189. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Valdecanas, D.; Niknam, S.; Peltier, H.J.; Diao, L.; Giri, U.; Komaki, R.; Calin, G.A.; Gomez, D.R.; Chang, J.Y.; et al. In Vivo Delivery of miR-34a Sensitizes Lung Tumors to Radiation Through RAD51 Regulation. Mol. Ther. Nucleic Acids 2015, 4, e270. [Google Scholar] [CrossRef]
- Raeisi, F.; Mahmoudi, E.; Dehghani-Samani, M.; Hosseini, S.S.E.; Ghahfarrokhi, A.M.; Arshi, A.; Forghanparast, K.; Ghazanfari, S. Differential Expression Profile of miR-27b, miR-29a, and miR-155 in Chronic Lymphocytic Leukemia and Breast Cancer Patients. Mol. Ther. Oncolytics 2020, 16, 230–237. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Sutandyo, N. The role of microRNA in cancer cachexia and muscle wasting: A review article. Casp. J. Intern. Med. 2021, 12, 124–128. [Google Scholar] [CrossRef]
- Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Fearon, K.; Hütterer, E.; Isenring, E.; Kaasa, S.; et al. ESPEN guidelines on nutrition in cancer patients. Clin. Nutr. 2017, 36, 11–48. [Google Scholar] [CrossRef] [PubMed]
- Powrózek, T.; Mlak, R.; Brzozowska, A.; Mazurek, M.; Gołębiowski, P.; Małecka-Massalska, T. miRNA-130a Significantly Improves Accuracy of SGA Nutritional Assessment Tool in Prediction of Malnutrition and Cachexia in Radiotherapy-Treated Head and Neck Cancer Patients. Cancers 2018, 10, 294. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.; Schaalan, M.; Abdallah, D.M.; Saad, A.S.; Sarhan, N.; Saleh, S. Impact of TNF-α Gene Polymorphisms on Pancreatic and Non-Small Cell Lung Cancer-Induced Cachexia in Adult Egyptian Patients: A Focus on Pathogenic Trajectories. Front. Oncol. 2021, 11, 4739. [Google Scholar] [CrossRef]
- Bing, C.; Russell, S.; Becket, E.; Pope, M.; Tisdale, M.J.; Trayhurn, P.; Jenkins, J.R. Adipose atrophy in cancer cachexia: Morphologic and molecular analysis of adipose tissue in tumour-bearing mice. Br. J. Cancer 2006, 95, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, S.; Lane, M.D. Regulating adipogenesis. J. Biol. Chem. 1997, 272, 5367–5370. [Google Scholar] [CrossRef]
- Chen, L.; Song, J.; Cui, J.; Hou, J.; Zheng, X.; Li, C.; Liu, L. microRNAs regulate adipocyte differentiation. Cell Biol. Int. 2013, 37, 533–546. [Google Scholar] [CrossRef]
- Blum, D.; Omlin, A.; Baracos, V.E.; Solheim, T.S.; Tan, B.H.L.; Stone, P.; Kaasa, S.; Fearon, K.; Strasser, F. Cancer cachexia: A systematic literature review of items and domains associated with involuntary weight loss in cancer. Crit. Rev. Oncol. Hematol. 2011, 80, 114–144. [Google Scholar] [CrossRef]
- Tsoli, M.; Moore, M.; Burg, D.; Painter, A.; Taylor, R.; Lockie, S.H.; Turner, N.; Warren, A.; Cooney, G.; Oldfield, B.; et al. Activation of thermogenesis in brown adipose tissue and dysregulated lipid metabolism associated with cancer cachexia in mice. Cancer Res. 2012, 72, 4372–4382. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Eder, S.; Schauer, S.; Diwoky, C.; Temmel, H.; Guertl, B.; Gorkiewicz, G.; Tamilarasan, K.P.; Kumari, P.; Trauner, M.; et al. Adipose triglyceride lipase contributes to cancer-associated cachexia. Science 2011, 333, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.P.; Koons, J.E.; Tan, E.T.; Grigor, M.R. Modified lipoprotein lipase activities, rates of lipogenesis, and lipolysis as factors leading to lipid depletion in C57BL mice bearing the preputial gland tumor, ESR-586. Cancer Res. 1981, 41, 3228–3232. [Google Scholar] [PubMed]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Di, W.; Zhang, W.; Zhu, B.; Li, X.; Tang, Q.; Zhou, Y. Colorectal cancer prompted adipose tissue browning and cancer cachexia through transferring exosomal miR-146b-5p. J. Cell. Physiol. 2021, 236, 5399–5410. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Sun, S.; Li, Z.; Yang, Q.; Li, B.; Zhu, S.; Wang, L.; Wu, J.; Yuan, J.; Wang, C.; et al. Breast cancer-released exosomes trigger cancer-associated cachexia to promote tumor progression. Adipocyte 2019, 8, 31–45. [Google Scholar] [CrossRef]
- Kulyté, A.; Lorente-Cebrián, S.; Gao, H.; Mejhert, N.; Agustsson, T.; Arner, P.; Rydén, M.; Dahlman, I. MicroRNA profiling links miR-378 to enhanced adipocyte lipolysis in human cancer cachexia. Am. J. Physiol. Endocrinol. Metab. 2014, 306, 267–274. [Google Scholar] [CrossRef]
- Güller, I.; Russell, A.P. MicroRNAs in skeletal muscle: Their role and regulation in development, disease and function. J. Physiol. 2010, 588, 4075–4087. [Google Scholar] [CrossRef]
- Armstrong, V.S.; Fitzgerald, L.W.; Bathe, O.F. Cancer-Associated Muscle Wasting—Candidate Mechanisms and Molecular Pathways. Int. J. Mol. Sci. 2020, 21, 9268. [Google Scholar] [CrossRef]
- Argilés, J.M.; Busquets, S.; Felipe, A.; López-Soriano, F.J. Molecular mechanisms involved in muscle wasting in cancer and ageing: Cachexia versus sarcopenia. Int. J. Biochem. Cell Biol. 2005, 37, 1084–1104. [Google Scholar] [CrossRef]
- Burckart, K.; Beca, S.; Urban, R.J.; Sheffield-Moore, M. Pathogenesis of muscle wasting in cancer cachexia: Targeted anabolic and anticatabolic therapies. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 410–416. [Google Scholar] [CrossRef] [Green Version]
- He, W.A.; Calore, F.; Londhe, P.; Canella, A.; Guttridge, D.C.; Croce, C.M. Microvesicles containing miRNAs promote muscle cell death in cancer cachexia via TLR7. Proc. Natl. Acad. Sci. USA 2014, 111, 4525–4529. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Zhang, W.; Feng, L.; Gu, X.; Shen, Q.; Lu, S.; Fan, M.; Li, Y.; Guo, X.; Ma, Y.; et al. Cancer-derived exosome miRNAs induce skeletal muscle wasting by Bcl-2-mediated apoptosis in colon cancer cachexia. Mol. Ther. Nucleic Acids 2021, 24, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.J.; Cagnin, S.; Chemello, F.; Silvestrin, M.; Musaro, A.; De Pitta, C.; Lanfranchi, G.; Sandri, M. Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions. J. Biol. Chem. 2014, 289, 21909–21925. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Brown, J.L.; Rosa-Caldwell, M.E.; Blackwell, T.A.; Perry, R.A.; Brown, L.A.; Khatri, B.; Seo, D.; Bottje, W.G.; Washington, T.A.; et al. Cancer cachexia-induced muscle atrophy: Evidence for alterations in microRNAs important for muscle size. Physiol. Genom. 2017, 49, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, A.; Ghosh, S.; Stretch, C.; Greiner, R.; Bathe, O.F.; Baracos, V.; Damaraju, S. Small RNAome profiling from human skeletal muscle: Novel miRNAs and their targets associated with cancer cachexia. J. Cachexia. Sarcopenia Muscle 2017, 8, 405–416. [Google Scholar] [CrossRef]
- Van de Worp, W.R.P.H.; Schols, A.M.W.J.; Dingemans, A.M.C.; Op den Kamp, C.M.H.; Degens, J.H.R.J.; Kelders, M.C.J.M.; Coort, S.; Woodruff, H.C.; Kratassiouk, G.; Harel-Bellan, A.; et al. Identification of microRNAs in skeletal muscle associated with lung cancer cachexia. J. Cachexia. Sarcopenia Muscle 2020, 11, 452–463. [Google Scholar] [CrossRef]
- Chen, D.; Goswami, C.P.; Burnett, R.M.; Anjanappa, M.; Bhat-Nakshatri, P.; Muller, W.; Nakshatri, H. Cancer affects microRNA expression, release, and function in cardiac and skeletal muscle. Cancer Res. 2014, 74, 4270–4281. [Google Scholar] [CrossRef]
- Okugawa, Y.; Yao, L.; Toiyama, Y.; Yamamoto, A.; Shigemori, T.; Yin, C.; Omura, Y.; Ide, S.; Kitajima, T.; Shimura, T.; et al. Prognostic impact of sarcopenia and its correlation with circulating miR-21 in colorectal cancer patients. Oncol. Rep. 2018, 39, 1555–1564. [Google Scholar] [CrossRef]
- Pin, F.; Beltrà, M.; Garcia-Castillo, L.; Pardini, B.; Birolo, G.; Matullo, G.; Penna, F.; Guttridge, D.; Costelli, P. Extracellular vesicles derived from tumour cells as a trigger of energy crisis in the skeletal muscle. J. Cachexia. Sarcopenia Muscle 2022, 13, 481–494. [Google Scholar] [CrossRef]
- Scott, I.C.; Tomlinson, W.; Walding, A.; Isherwood, B.; Dougall, I.G. Large-scale isolation of human skeletal muscle satellite cells from post-mortem tissue and development of quantitative assays to evaluate modulators of myogenesis. J. Cachexia. Sarcopenia Muscle 2013, 4, 157–169. [Google Scholar] [CrossRef]
- Okugawa, Y.; Toiyama, Y.; Hur, K.; Yamamoto, A.; Yin, C.; Ide, S.; Kitajima, T.; Fujikawa, H.; Yasuda, H.; Koike, Y.; et al. Circulating miR-203 derived from metastatic tissues promotes myopenia in colorectal cancer patients. J. Cachexia. Sarcopenia Muscle 2019, 10, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.L.P.; Tobias, G.C.; Fernandes, T.; Silveira, A.C.; Negrão, C.E.; Chammas, R.; Brum, P.C.; Oliveira, E.M. Effects of Aerobic Exercise Training on MyomiRs Expression in Cachectic and Non-Cachectic Cancer Mice. Cancers 2021, 13, 5728. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Xiong, H.; Xiao, W.; Xiong, Z.; Hu, W.; Ye, J.; Xu, N.; Shi, J.; Yuan, C.; Chen, Z.; et al. Downregulation of miR-29c promotes muscle wasting by modulating the activity of leukemia inhibitory factor in lung cancer cachexia. Cancer Cell Int. 2021, 21, 627. [Google Scholar] [CrossRef] [PubMed]
- Nicola, N.A.; Babon, J.J. Leukemia inhibitory factor (LIF). Cytokine Growth Factor Rev. 2015, 26, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Authors (Year of Publication) | Study Group/Cancer Type | Race/Nationality | Anticancer Treatment (Surgery, CTH, RT) | Study Material | Studied Mirna | Time-Point of miRNA Assessment (before or after Anticancer Treatment/Experimental Model Manipulation) | miRNA Change in Patients with Cachexia or in Cachectic Models | miRNA-Related Targets/Changes in the Studied Material |
|---|---|---|---|---|---|---|---|---|
| Powrozek et al. (2018) [33] | Patients with HNC (n = 70, without cachexia, n = 33, with cachexia, n = 37) | Poland | IMRT | Plasma | miR-130a | Before | Downregulated | TNF-α |
| Fabbri et al. (2012) [34] | B6 mice with LLC (n = 18) Cell Cultures (A549, SK-MES, HEK-293) | - | - | Cell Culture/animal model | miR-21 miR-29a | - | Upregulated | TLR (murine TLR7, human TLR8) |
| Yehia et al. (2021) [35] | Patients with pancreatic cancer or NSCLC (n = 203, without cachexia, n = 94, with cachexia, n = 109) | Egypt | CTH (GEM+CIS) | Serum | miR-155 | After | Upregulated | TNF-α (through TNF- α: SOCS1, TAB2, Foxp3) |
| Authors (Year of Publication) | Study Group/Cancer Type | Race/Nationality | Anticancer Treatment (Surgery, CTH, RT) | Study Material | Studied miRNA | Time-Point of miRNA Assessment (before or after Anticancer Treatment) | miRNA Change | miRNA-Related Targets/Changes in the Studied Material |
|---|---|---|---|---|---|---|---|---|
| Wenjuan Di et al. (2021) [44] | Patients with colorectal cancer (n = 48) Healthy volunteers (n = 48) Male C57BL/6 mice (n = 6) | Asian (China) | - | Cancerous tissue samples | miR-146-5p | - | Upregulated | Browning of WAT Accelerated lipolysis Decreased oxygen consumption Fat mass loss |
| Wu et al. (2019) [45] | Patients with breast cancer (n = 108) Cell culture (C2C12) | - | - | Cell culture | miR-155 | - | Upregulated | Browning of WAT Anomalous conversion and increased catabolism of muscle cells Lipolysis Muscle loss |
| Kulyté et al. (2014) [46] | Patients with gastrointestinal cancer (pancreas, stomach, liver metastasis) (n = 21, without cachexia, n = 11, with cachexia, n = 10) | Sweden | Surgery | Abdominal subcutaneous adipose tissue | miR-483–5p miR-23a miR-744 miR-99b miR-378 | Before | Upregulated (miR-378) Downregulated miR-483-5p miR-23a miR-744 miR-99b | Accelerated lipolysis (miR-378) |
| Sun et al. (2021) [16] | Patients with gastric cancer (n = 60, without cachexia, n = 30, with cachexia, n = 30) | Asian (China) | Surgery | Exosomes from serum/tissue | miR-410-3p | Before | Upregulated | Adipogenesis inhibition |
| Authors (Year of Publication) | Study Group | Race/Nationality | Anticancer Treatment (Surgery, CTH, RT) | Study Material | Studied miRNA | Time-Point of miRNA Assessment (before or after Anticancer Treatment) | miRNA Change | miRNA-Related Targets/Changes in the Studied Material |
|---|---|---|---|---|---|---|---|---|
| He et al. (2014) [51] | Wild-type C57B6 male mice with LLC (number of subjects not provided) Cell lines of lung cancer and pancreatic adenocarcinoma (A549, LCC, H460, AsPC-1, Panc-2, MEFs MDA-MB-231, MIA-PaCa, MCF7, C2C12) | - | - | Wild-type C57B6 male mice serum Cell lines | miR-21 | - | Upregulated in myoblasts (cell line) | Myoblast apoptosis |
| Miao et al. (2021) [52] | Male BALB/c mice without tumour (n = 8) and with C26 (n = 8) Patients with colon cancer (n = 21, without cachexia n = 13, with cachexia n = 8) Healthy volunteers (n = 19) Cell line (C2C12) | Asian (China) | - | Serum and exosomes from mice and human subjects Mice muscle tissue Cell line | miR-195a-5p, miR-125b-1-3p | - | Upregulated | Downregulation of Bcl-2 gene Muscle atrophy |
| Soares et al. (2014) [53] | BALB/c mice with colon carcinoma (C26) (n = 3) Cell line (C2C12) | - | - | Muscle tissue Cell line | miR-206 miR-21 | - | Upregulated | Numerous different genes regulated by both miRNAs Muscle atrophy |
| Lee et al. (2017) [54] | C57BL6/J mice with LLC (n = 8) or without LLC (control, n = 8) | - | - | Muscle tissue | miR-147-3p miR-299a-3p miR -1933-3p miR-511-3p miR-3473d miR-233-3p miR-431-5p miR-665-3p miR-205-3p | - | Downregulated: miR-229a-3p miR-431-5p miR-665-3p miR-1933-3p miR-3473d Upregulated: miR-147-3p miR-205-5p miR-223-3p miR-511-3p | Muscle atrophy |
| Narasimhan et al. (2017) [55] | Patients with pancreatic and colorectal cancer (with liver metastasis) (n = 42, without cachexia n = 20, with cachexia n = 22) | Canada | Surgery | Muscle tissues (biopsies) | hsa-let-7d-3p hsa-miR-345-5p hsa-miR-423-5p hsa-miR-532-5p hsa-miR-1296-5p hsa-miR-3184-3p hsa-miR-423-3p hsa-miR-199a-3p | Before treatment | Upregulated | Numerous different genes regulated by studied miRNAs Muscle atrophy |
| van de Worp et al. (2020) [56] | NSCLC patients (n = 26, 11 without cachexia, 15 with cachexia) Healthy volunteers (n = 22) | Netherlands | - | Muscle tissue (biopsies) | hsa-miR-424-5p hsa-miR-451a hsa-miR-144-5p hsa-miR-424-3p hsa-miR-450a-5p | Before treatment | Upregulated: miR-424-5p miR-424-3p miR-450a Downregulated:miR-451a miR-144-5p | Muscle atrophy |
| Chen et al. (2014) [57] | Patients with breast cancer (n = 79) MMTV-PyMT (n = 4) and MMTV-Her2 (n = 4) transgenic mice Cell culture (C2C12) | USA | - | Human serumTransgenic mice models (serum, cancerous tissue, cardiac and skeletal muscle tissue) Cell culture | miR-486 | - | Downregulated | Downregulation of PTEN and FOXO1A genes Decreased signalling through PI3K/Akt pathway Downregulation of MyoD transcription factor Muscle atrophy |
| Okugawa et al. (2018) [58] | Patients with colorectal cancer (serum, n = 153, cancerous tissue, n = 134) | Asian (Japan) | - | SerumCancerous tissue | miR-21 | Before surgery | Upregulated | Sarcopenia Low PMI |
| Pin et al. (2022) [59] | BALB/c mice with or without C26 (n = 8) Male Wistar rats with or without AH-130 (n = 8) Cell lines (C2C12, LCC, C26) | - | - | Microvesicles from C26 or LLC cells Plasma of mice and rats Cell line | miR-181a-5p miR-375-3p miR-455-5p miR-148a-3p miR-181a-5p | - | Downregulated miR-181a-5p miR-375-3p miR-455-5p Upregulated miR-148a-3p miR-181a-5p | Upregulated miRNAs lead to downregulation of Myf5, Myog, Myh4, Myh7 genes Delayed differentiation Disorganized mitochondrial system |
| Okugawa et al. (2019) [61] | Patients with colorectal cancer (n = 183) Cell line (SkMC) | Asian (Japan) | CTH (5-FU) | Cancerous tissue Serum Cell line | miR-203 | Before surgery | Upregulated | Downregulation of BIRC5 Decreased skeletal muscles mass (decreased PMI) Skeletal muscle cells proliferation inhibition and apoptosis |
| Gomes et al. (2021) [62] | BALB/c female healthy mice (n = 17) or with colon cancer (n = 19) c57 healthy female mice (n = 13) or non-cachectic transgenic mice with spontaneous breast cancer (n = 16) | - | - | Serum Skeletal muscle tissue | miRNA-486 miRNA -206 | - | Upregulated miR-206 (Breast cancer) Downregulated miR-206 (Colon cancer) miRNA-486 | Muscle wasting |
| Xie et al. (2021) [63] | C57BL/6 mice with (n = 3) or without (n = 3) LLC Cell culture (C2C12) | - | - | C57BL/6 mice tissue Cell culture | miR-29c | - | Downregulated | Upregulation of LIF, JAK/STAT and p38 MAPK pathwaysMuscle wasting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homa-Mlak, I.; Pigoń-Zając, D.; Wawrejko, P.; Małecka-Massalska, T.; Mlak, R. Three Pathways of Cancer Cachexia: Inflammation, Changes in Adipose Tissue and Loss of Muscle Mass—The Role of miRNAs. J. Pers. Med. 2022, 12, 1438. https://doi.org/10.3390/jpm12091438
Homa-Mlak I, Pigoń-Zając D, Wawrejko P, Małecka-Massalska T, Mlak R. Three Pathways of Cancer Cachexia: Inflammation, Changes in Adipose Tissue and Loss of Muscle Mass—The Role of miRNAs. Journal of Personalized Medicine. 2022; 12(9):1438. https://doi.org/10.3390/jpm12091438
Chicago/Turabian StyleHoma-Mlak, Iwona, Dominika Pigoń-Zając, Paweł Wawrejko, Teresa Małecka-Massalska, and Radosław Mlak. 2022. "Three Pathways of Cancer Cachexia: Inflammation, Changes in Adipose Tissue and Loss of Muscle Mass—The Role of miRNAs" Journal of Personalized Medicine 12, no. 9: 1438. https://doi.org/10.3390/jpm12091438
APA StyleHoma-Mlak, I., Pigoń-Zając, D., Wawrejko, P., Małecka-Massalska, T., & Mlak, R. (2022). Three Pathways of Cancer Cachexia: Inflammation, Changes in Adipose Tissue and Loss of Muscle Mass—The Role of miRNAs. Journal of Personalized Medicine, 12(9), 1438. https://doi.org/10.3390/jpm12091438