
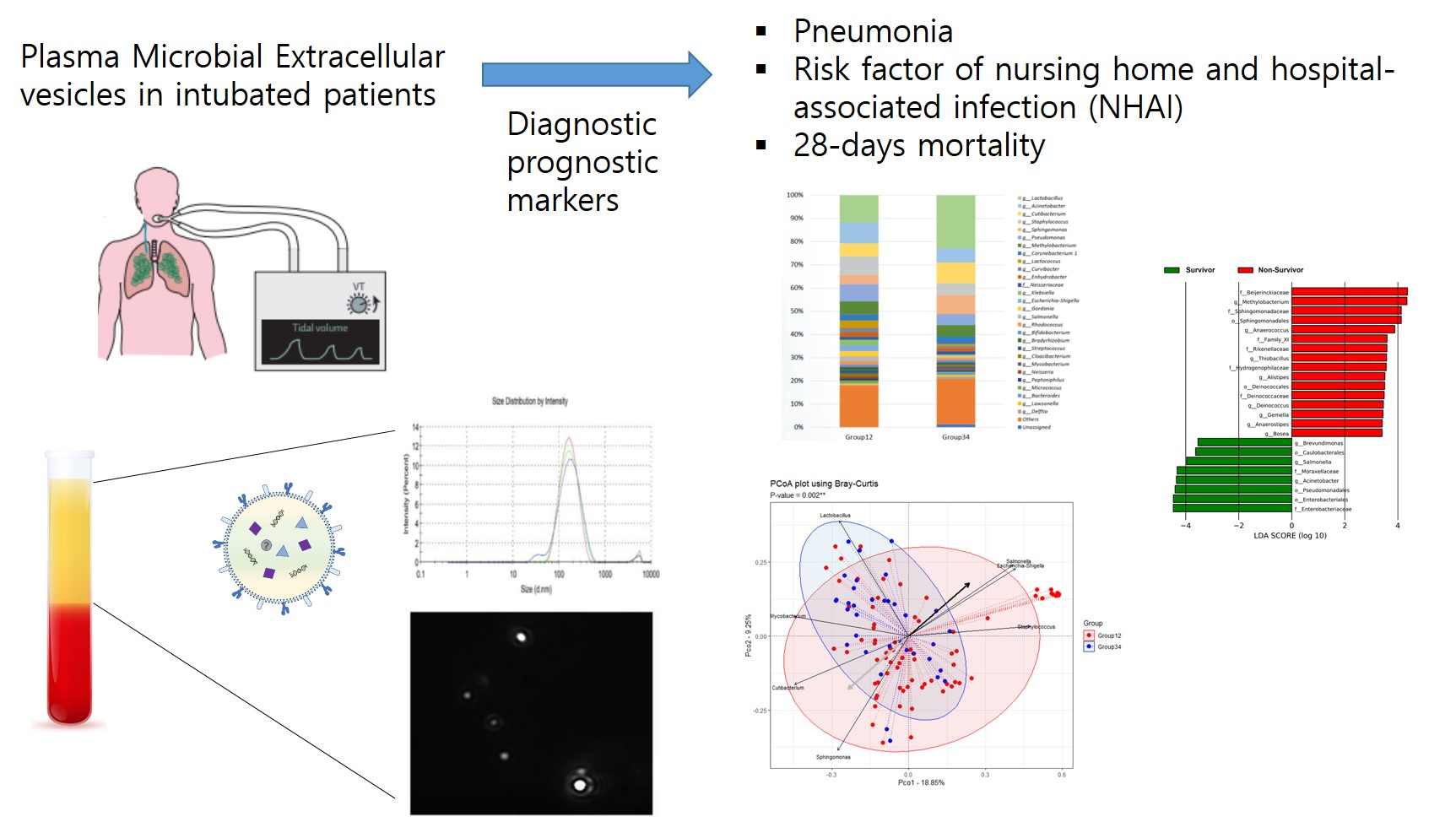
Metagenomic Analysis of Plasma Microbial Extracellular Vesicles in Patients Receiving Mechanical Ventilation: A Pilot Study

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Population and Plasma Sample Collection
2.2. EV Isolation and DNA Extraction from Human Plasma
2.3. Amplicon Sequencing Using EV DNA from Human Plasma Samples and Data Processing
2.4. Statistical Analysis
3. Results
3.1. Demographic Characteristics of the Study Population
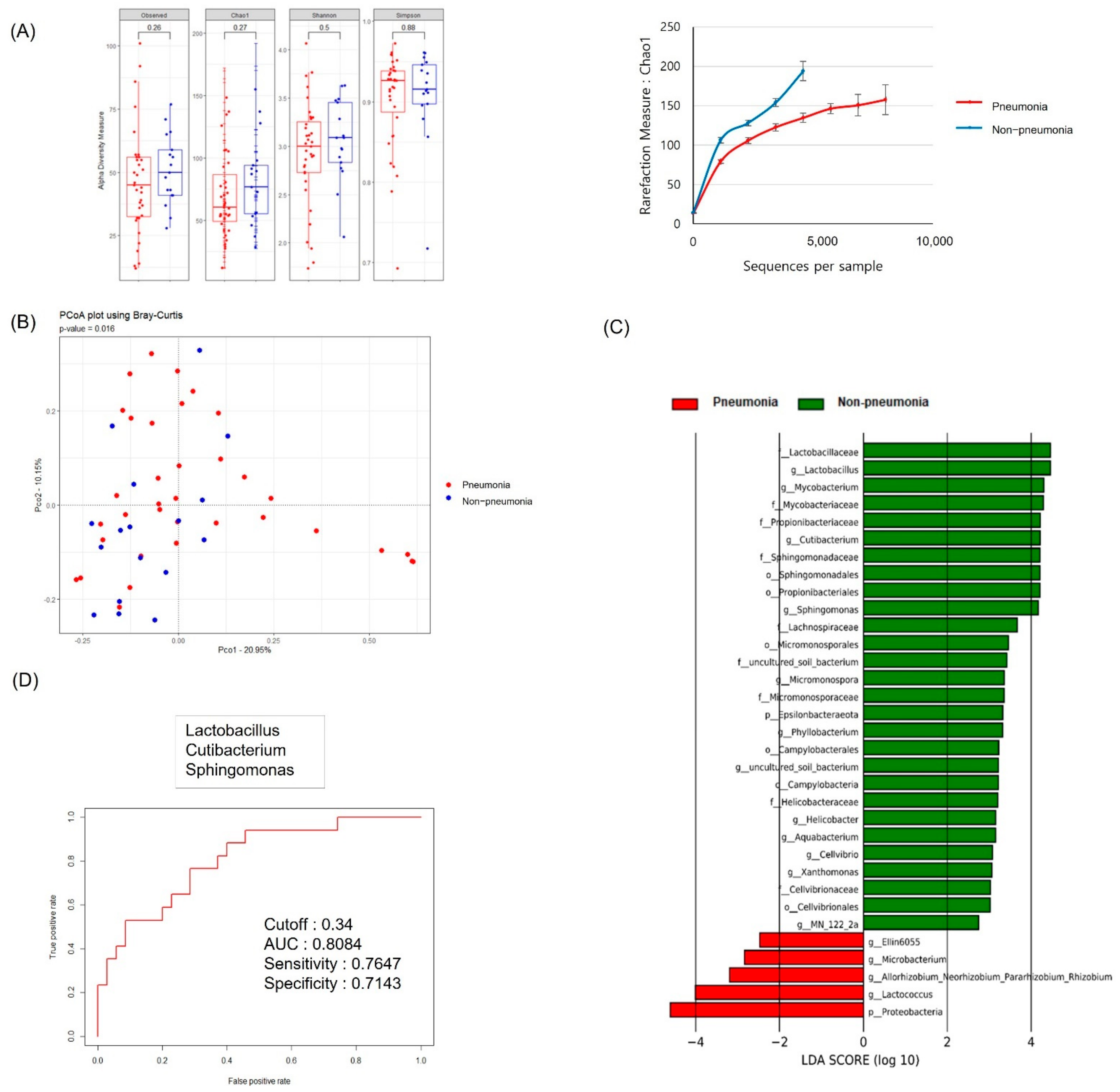
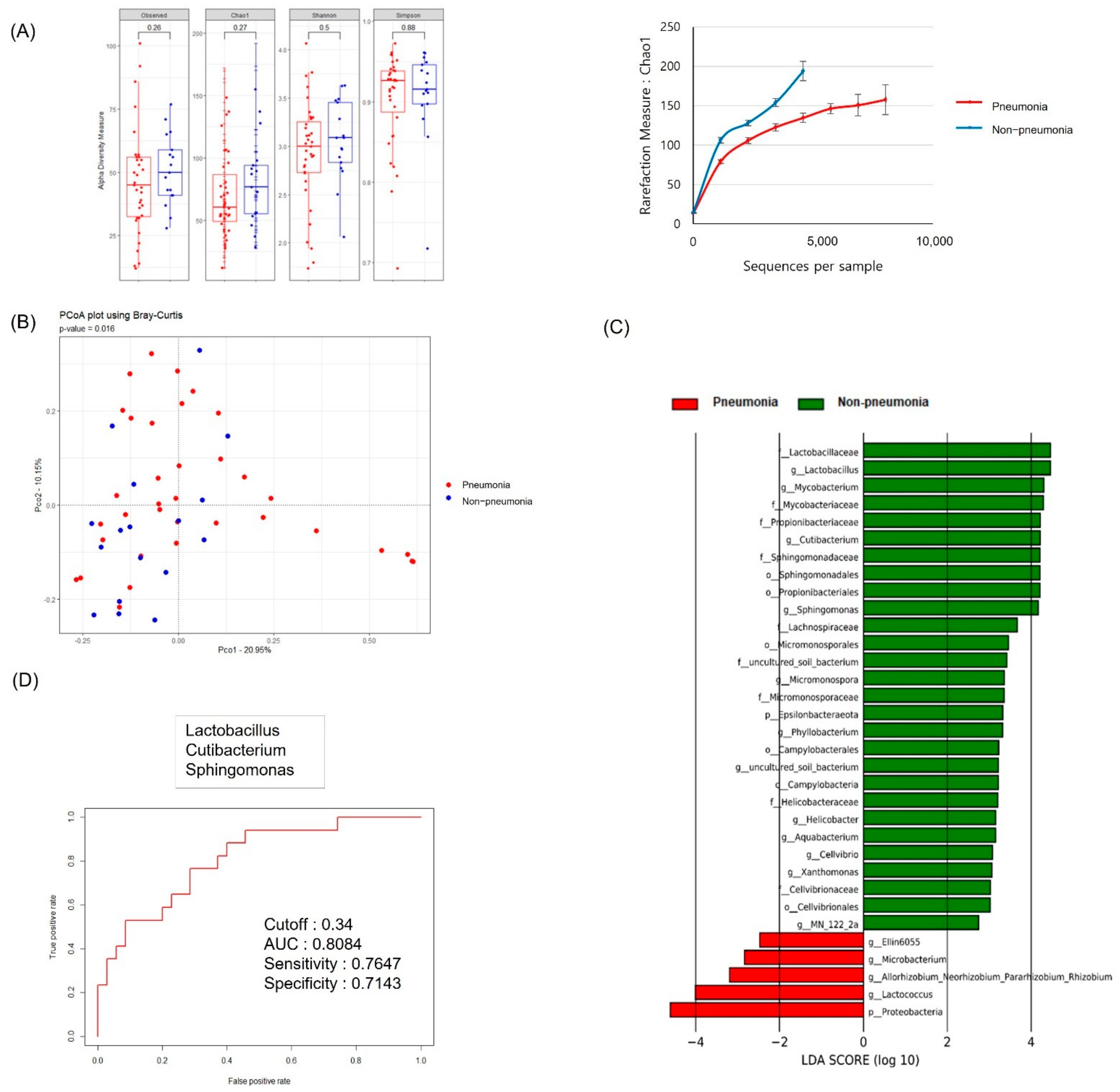
3.2. Serum MEV Diversity by Clinical Group
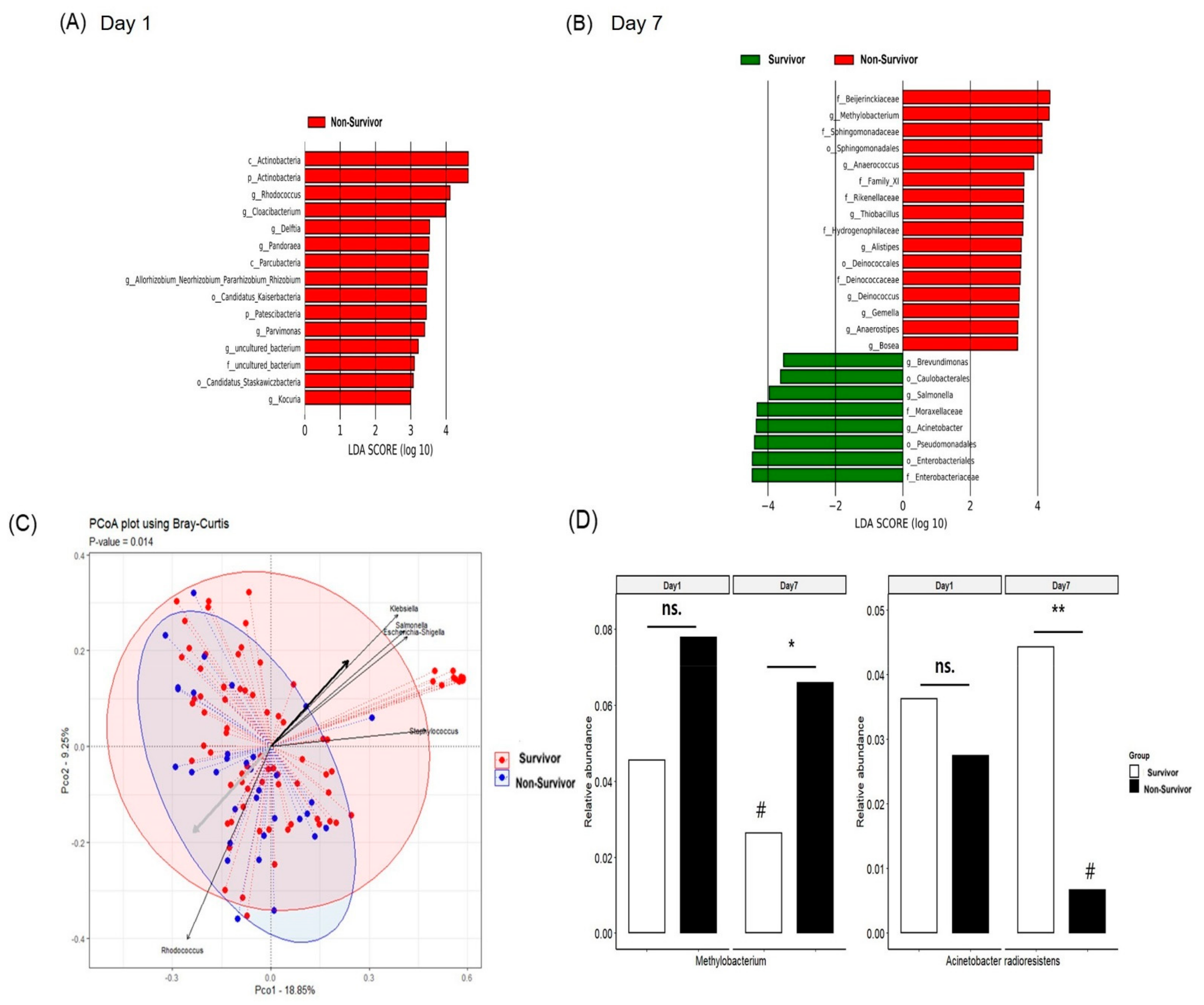
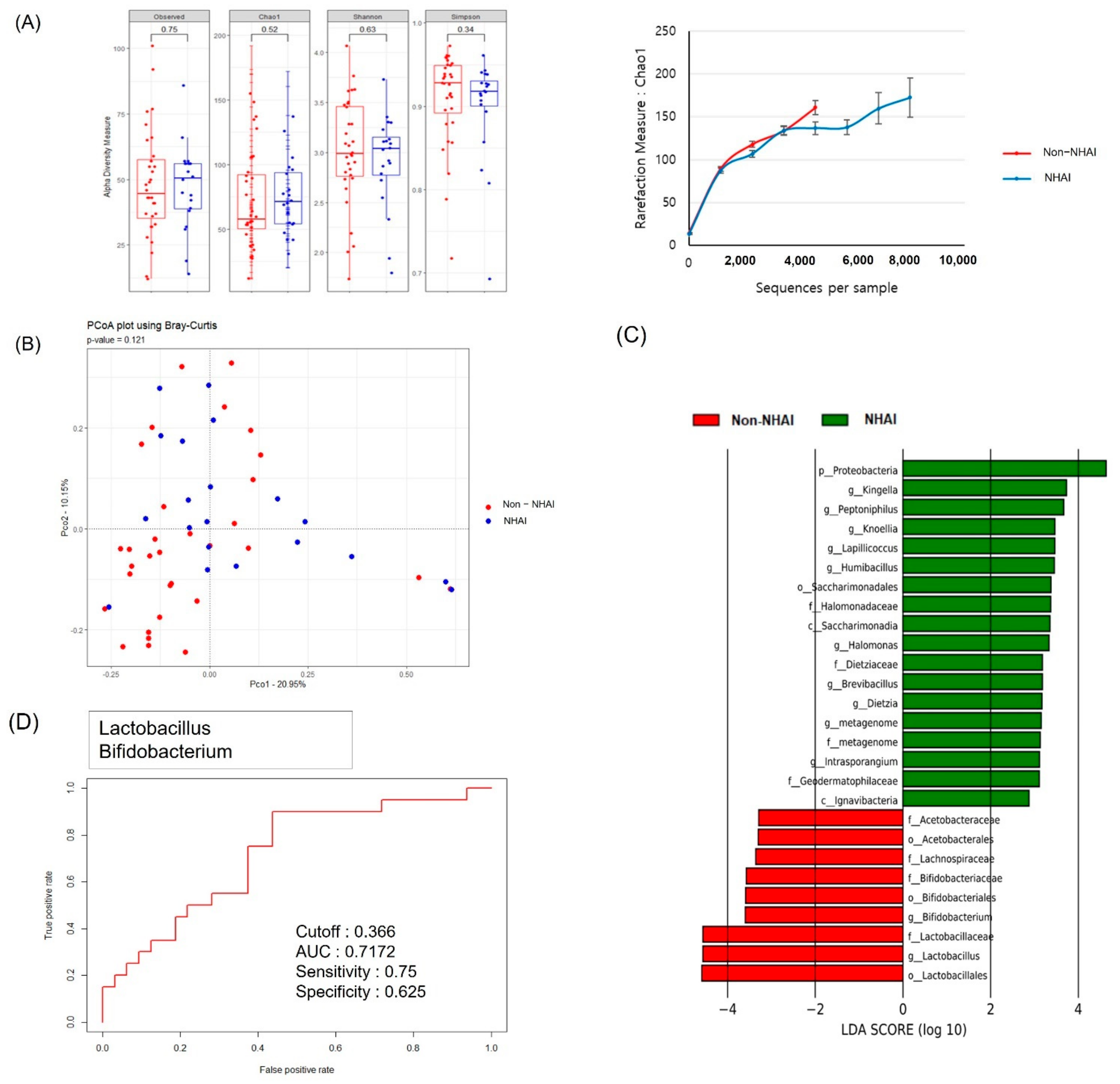
3.3. Comparison of the Diversity of Serum MEV Composition between Survivors and Non-Survivors
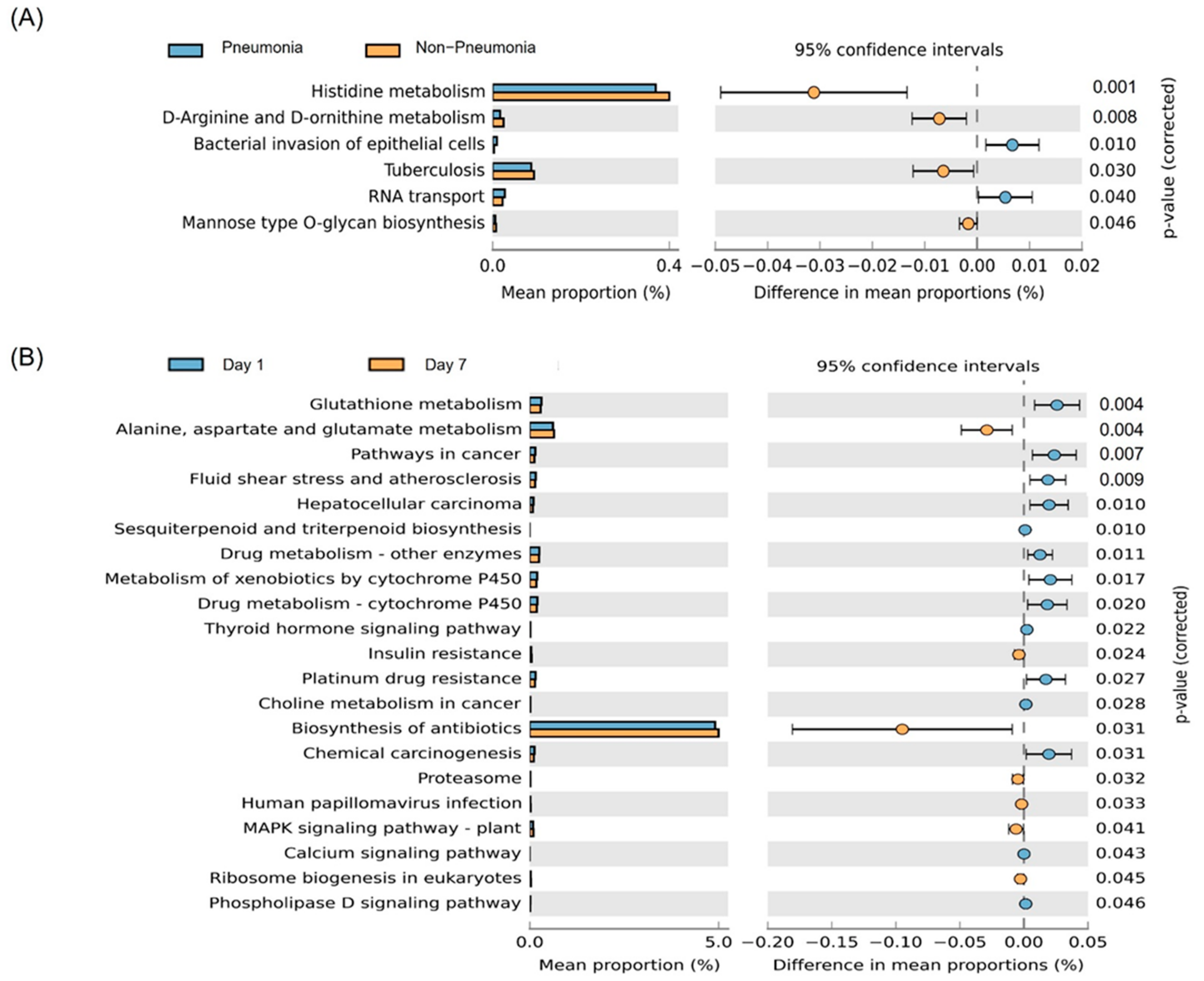
3.4. Predicted Functions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaparakis-Liaskos, M.; Ferrero, R.L. Immune modulation by bacterial outer membrane vesicles. Nat. Rev. Immunol. 2015, 15, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Kadota, T.; Araya, J.; Ochiya, T.; Kuwano, K. Extracellular Vesicles: New Players in Lung Immunity. Am. J. Respir. Cell Mol. Biol. 2018, 58, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-negative and Gram-positive bacterial extracellular vesicles. Semin. Cell Dev. Biol. 2015, 40, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.R.; Hong, S.W.; Choi, E.B.; Lee, W.H.; Kim, Y.S.; Jeon, S.G.; Jang, M.H.; Gho, Y.S.; Kim, Y.K. Staphylococcus aureus-derived extracellular vesicles induce neutrophilic pulmonary inflammation via both Th1 and Th17 cell responses. Allergy 2012, 67, 1271–1281. [Google Scholar] [CrossRef]
- Kim, Y.S.; Choi, E.J.; Lee, W.H.; Choi, S.J.; Roh, T.Y.; Park, J.; Jee, Y.K.; Zhu, Z.; Koh, Y.Y.; Gho, Y.S.; et al. Extracellular vesicles, especially derived from Gram-negative bacteria, in indoor dust induce neutrophilic pulmonary inflammation associated with both Th1 and Th17 cell responses. Clin. Exp. Allergy 2013, 43, 443–454. [Google Scholar] [CrossRef]
- Shukla, S.D.; Budden, K.F.; Neal, R.; Hansbro, P.M. Microbiome effects on immunity, health and disease in the lung. Clin. Transl. Immunol. 2017, 6, e133. [Google Scholar] [CrossRef]
- Choi, Y.; Park, H.; Park, H.S.; Kim, Y.K. Extracellular Vesicles, a Key Mediator to Link Environmental Microbiota to Airway Immunity. Allergy Asthma Immunol. Res. 2017, 9, 101–106. [Google Scholar] [CrossRef]
- Codemo, M.; Muschiol, S.; Iovino, F.; Nannapaneni, P.; Plant, L.; Wai, S.N.; Henriques-Normark, B. Immunomodulatory Effects of Pneumococcal Extracellular Vesicles on Cellular and Humoral Host Defenses. mBio 2018, 9, e00559-18. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Self, W.H.; Wunderink, R.G.; Fakhran, S.; Balk, R.; Bramley, A.M.; Reed, C.; Grijalva, C.G.; Anderson, E.J.; Courtney, D.M.; et al. Community-Acquired Pneumonia Requiring Hospitalization among U.S. Adults. N. Engl. J. Med. 2015, 373, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Morris, A.C. Management of pneumonia in intensive care. J. Emerg. Crit. Care Med. 2018, 2, 101. [Google Scholar] [CrossRef]
- Shindo, Y.; Ito, R.; Kobayashi, D.; Ando, M.; Ichikawa, M.; Shiraki, A.; Goto, Y.; Fukui, Y.; Iwaki, M.; Okumura, J.; et al. Risk factors for drug-resistant pathogens in community-acquired and healthcare-associated pneumonia. Am. J. Respir. Crit. Care Med. 2013, 188, 985–995. [Google Scholar] [CrossRef]
- Woo, S.; Park, S.Y.; Kim, Y.; Jeon, J.P.; Lee, J.J.; Hong, J.Y. The Dynamics of Respiratory Microbiota during Mechanical Ventilation in Patients with Pneumonia. J. Clin. Med. 2020, 9, 638. [Google Scholar] [CrossRef] [Green Version]
- Shin, T.S.; Kim, J.H.; Kim, Y.S.; Jeon, S.G.; Zhu, Z.; Gho, Y.S.; Kim, Y.K. Extracellular vesicles are key intercellular mediators in the development of immune dysfunction to allergens in the airways. Allergy 2010, 65, 1256–1265. [Google Scholar] [CrossRef] [Green Version]
- Metlay, J.P.; Waterer, G.W.; Long, A.C.; Anzueto, A.; Brozek, J.; Crothers, K.; Cooley, L.A.; Dean, N.C.; Fine, M.J.; Flanders, S.A.; et al. Diagnosis and Treatment of Adults with Community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 2019, 200, e45–e67. [Google Scholar] [CrossRef]
- Kalil, A.C.; Metersky, M.L.; Klompas, M.; Muscedere, J.; Sweeney, D.A.; Palmer, L.B.; Napolitano, L.M.; O’Grady, N.P.; Bartlett, J.G.; Carratala, J.; et al. Management of Adults with Hospital-acquired and Ventilator-associated Pneumonia: 2016 Clinical Practice Guidelines by the Infectious Diseases Society of America and the American Thoracic Society. Clin. Infect. Dis. 2016, 63, e61–e111. [Google Scholar] [CrossRef]
- Baek, M.G.; Woo, S.J.; Kim, N.E.; Baek, C.; Won, S.; Kim, Y.; Lee, J.J.; Yi, H.; Hong, J.Y. Respiratory microbiome profiles differ by recent hospitalization and nursing home residence in patients on mechanical ventilation. J. Transl. Med. 2020, 18, 464. [Google Scholar] [CrossRef]
- Kim, J.R.; Han, K.; Han, Y.; Kang, N.; Shin, T.S.; Park, H.J.; Kim, H.; Kwon, W.; Lee, S.; Kim, Y.K.; et al. Microbiome Markers of Pancreatic Cancer Based on Bacteria-Derived Extracellular Vesicles Acquired from Blood Samples: A Retrospective Propensity Score Matching Analysis. Biology 2021, 10, 219. [Google Scholar] [CrossRef]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef]
- Yang, J.; McDowell, A.; Seo, H.; Kim, S.; Min, T.K.; Jee, Y.K.; Choi, Y.; Park, H.S.; Pyun, B.Y.; Kim, Y.K. Diagnostic Models for Atopic Dermatitis Based on Serum Microbial Extracellular Vesicle Metagenomic Analysis: A Pilot Study. Allergy Asthma Immunol. Res. 2020, 12, 792–805. [Google Scholar] [CrossRef]
- Schuijt, T.J.; Lankelma, J.M.; Scicluna, B.P.; de Sousa e Melo, F.; Roelofs, J.J.; de Boer, J.D.; Hoogendijk, A.J.; de Beer, R.; de Vos, A.; Belzer, C.; et al. The gut microbiota plays a protective role in the host defence against pneumococcal pneumonia. Gut 2016, 65, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Abston, E.; Zhang, D.; Rai, A.; Jin, Y. Extracellular Vesicle: An Emerging Mediator of Intercellular Crosstalk in Lung Inflammation and Injury. Front. Immunol. 2018, 9, 924. [Google Scholar] [CrossRef]
- Guervilly, C.; Lacroix, R.; Forel, J.M.; Roch, A.; Camoin-Jau, L.; Papazian, L.; Dignat-George, F. High levels of circulating leukocyte microparticles are associated with better outcome in acute respiratory distress syndrome. Crit. Care 2011, 15, R31. [Google Scholar] [CrossRef] [Green Version]
- Shaver, C.M.; Woods, J.; Clune, J.K.; Grove, B.S.; Wickersham, N.E.; McNeil, J.B.; Shemancik, G.; Ware, L.B.; Bastarache, J.A. Circulating microparticle levels are reduced in patients with ARDS. Crit. Care 2017, 21, 120. [Google Scholar] [CrossRef] [Green Version]
- Paisse, S.; Valle, C.; Servant, F.; Courtney, M.; Burcelin, R.; Amar, J.; Lelouvier, B. Comprehensive description of blood microbiome from healthy donors assessed by 16S targeted metagenomic sequencing. Transfusion 2016, 56, 1138–1147. [Google Scholar] [CrossRef]
- Zakharkina, T.; Martin-Loeches, I.; Matamoros, S.; Povoa, P.; Torres, A.; Kastelijn, J.B.; Hofstra, J.J.; de Wever, B.; de Jong, M.; Schultz, M.J.; et al. The dynamics of the pulmonary microbiome during mechanical ventilation in the intensive care unit and the association with occurrence of pneumonia. Thorax 2017, 72, 803–810. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.J.; Imai, I.; Bittinger, K.; Laughlin, A.; Fuchs, B.D.; Bushman, F.D.; Collman, R.G. Composition and dynamics of the respiratory tract microbiome in intubated patients. Microbiome 2016, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Corvec, S. Clinical and Biological Features of Cutibacterium (Formerly Propionibacterium) avidum, an Underrecognized Microorganism. Clin. Microbiol. Rev. 2018, 31, e00064-17. [Google Scholar] [CrossRef] [Green Version]
- White, D.C.; Sutton, S.D.; Ringelberg, D.B. The genus Sphingomonas: Physiology and ecology. Curr. Opin. Biotechnol. 1996, 7, 301–306. [Google Scholar] [CrossRef]
- Qi, X.; Qu, H.; Yang, D.; Zhou, L.; He, Y.W.; Yu, Y.; Qu, J.; Liu, J. Lower respiratory tract microbial composition was diversified in Pseudomonas aeruginosa ventilator-associated pneumonia patients. Respir. Res. 2018, 19, 139. [Google Scholar] [CrossRef]
- Shimizu, K.; Yamada, T.; Ogura, H.; Mohri, T.; Kiguchi, T.; Fujimi, S.; Asahara, T.; Yamada, T.; Ojima, M.; Ikeda, M.; et al. Synbiotics modulate gut microbiota and reduce enteritis and ventilator-associated pneumonia in patients with sepsis: A randomized controlled trial. Crit. Care 2018, 22, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2011, 10, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, J.; Degener, J.E.; van der Mei, H.C. Methylobacterium and its role in health care-associated infection. J. Clin. Microbiol. 2014, 52, 1317–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savov, E.; Pfeifer, Y.; Wilharm, G.; Trifonova, A.; Todorova, I.; Gergova, I.; Borisova, M.; Kjoseva, E. Isolation of Acinetobacter radioresistens from a clinical sample in Bulgaria. J. Glob. Antimicrob. Resist. 2016, 4, 57–59. [Google Scholar] [CrossRef]
- Wang, T.; Costa, V.; Jenkins, S.G.; Hartman, B.J.; Westblade, L.F. Acinetobacter radioresistens infection with bacteremia and pneumonia. IDCases 2019, 15, e00495. [Google Scholar] [CrossRef]
- Alvarez, L.; Aliashkevich, A.; de Pedro, M.A.; Cava, F. Bacterial secretion of D-arginine controls environmental microbial biodiversity. ISME J. 2018, 12, 438–450. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150, 2570S–2575S. [Google Scholar] [CrossRef]
- Ilaiwy, A.; Ten Have, G.A.M.; Bain, J.R.; Muehlbauer, M.J.; O’Neal, S.K.; Berthiaume, J.M.; Parry, T.L.; Deutz, N.E.; Willis, M.S. Identification of Metabolic Changes in Ileum, Jejunum, Skeletal Muscle, Liver, and Lung in a Continuous I.V. Pseudomonas aeruginosa Model of Sepsis Using Nontargeted Metabolomics Analysis. Am. J. Pathol. 2019, 189, 1797–1813. [Google Scholar] [CrossRef]
- Lv, C.; Huang, L. Xenobiotic receptors in mediating the effect of sepsis on drug metabolism. Acta Pharm. Sin. B 2020, 10, 33–41. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pneumonia | Non-Pneumonia | p Value | NHAI | Non NHAI | p Value | |
|---|---|---|---|---|---|---|
| n = 41 | n = 18 | n = 24 | n = 35 | |||
| Age | 73 (60.5, 79.5) | 76 (59, 81) | 0.52 | 77.5 (72.3, 85.0) | 70 (57, 77) | 0.005 |
| Male | 30 (73.2%) | 12 (66.7%) | 0.76 | 15 (62.5%) | 27 (77.1%) | 0.254 |
| ARDS | 8 (19.5%) | 0 (0%) | 0.092 | 4 (16.7%) | 4 (11.4%) | 0.704 |
| Charlson comorbidity index | 3 (1, 4) | 2 (1, 2.3) | 0.079 | 3 (2.3, 5.5) | 1 (0, 2) | <0.001 |
| Cause of intubation | <0.001 | <0.001 | ||||
| Cardiac arrest | 1 (2.4%) | 3 (16.7%) | 0 (0%) | 4 (11.4%) | ||
| Neurological distress | 5 (12.2%) | 13 (72.2%) | 1 (4.2%) | 17 (48.6%) | ||
| Postoperative status | 0 (0%) | 1 (5.6%) | 0 (0%) | 1 (2.9%) | ||
| Respiratory | 35 (85.4%) | 1 (5.6%) | 23 (95.8%) | 13 (37.1%) | ||
| PaO2/FiO2 | 212.5 (133.1, 299.0) | 427.4 (304.5, 463.8) | <0.001 | 222 (130.9, 301.0) | 321 (182.8, 435.0) | 0.047 |
| Severity | ||||||
| APACHE score | 20 (16, 24) | 22.5 (18.5, 25.3) | 0.121 | 20.5 (16.3, 24.0) | 21 (17, 25) | 0.551 |
| SOFA score | 7 (6, 9) | 6 (4.5, 9.3) | 0.08 | 7 (6, 9) | 7 (5, 10) | 0.852 |
| GCS | 8 (6, 11) | 6 (5, 8.3) | 0.038 | 8.5 (6, 10.8) | 7 (6.9) | 0.241 |
| 28-day mortality | 12 (29.3%) | 6 (33.3%) | 0.77 | 7 (29.2%) | 11 (31.4%) | 1 |
| In-Hospital mortality | 19 (46.3%) | 6 (33.3%) | 0.403 | 11 (45.8%) | 14 (40.0%) | 0.79 |
| MV duration | 13 (8.0, 18.0) | 10 (6.8, 14.5) | 0.248 | 13 (8, 17.5) | 10 (7, 16) | 0.349 |
| CRP (mg/dL) | 132.7 (45.0, 213.5) | 61.6 (7.2, 135.5) | 0.022 | 124.2 (62.8, 192.4) | 96 (8, 205.6) | 0.195 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Lee, J.J.; Hong, Y.; Seo, H.; Shin, T.-S.; Hong, J.Y. Metagenomic Analysis of Plasma Microbial Extracellular Vesicles in Patients Receiving Mechanical Ventilation: A Pilot Study. J. Pers. Med. 2022, 12, 564. https://doi.org/10.3390/jpm12040564
Park J, Lee JJ, Hong Y, Seo H, Shin T-S, Hong JY. Metagenomic Analysis of Plasma Microbial Extracellular Vesicles in Patients Receiving Mechanical Ventilation: A Pilot Study. Journal of Personalized Medicine. 2022; 12(4):564. https://doi.org/10.3390/jpm12040564
Chicago/Turabian StylePark, Jinkyeong, Jae Jun Lee, Yoonki Hong, Hochan Seo, Tae-Seop Shin, and Ji Young Hong. 2022. "Metagenomic Analysis of Plasma Microbial Extracellular Vesicles in Patients Receiving Mechanical Ventilation: A Pilot Study" Journal of Personalized Medicine 12, no. 4: 564. https://doi.org/10.3390/jpm12040564
APA StylePark, J., Lee, J. J., Hong, Y., Seo, H., Shin, T.-S., & Hong, J. Y. (2022). Metagenomic Analysis of Plasma Microbial Extracellular Vesicles in Patients Receiving Mechanical Ventilation: A Pilot Study. Journal of Personalized Medicine, 12(4), 564. https://doi.org/10.3390/jpm12040564