
High Angiotensin-Converting Enzyme and Low Carboxypeptidase N Serum Activity Correlate with Disease Severity in COVID-19 Patients



Abstract
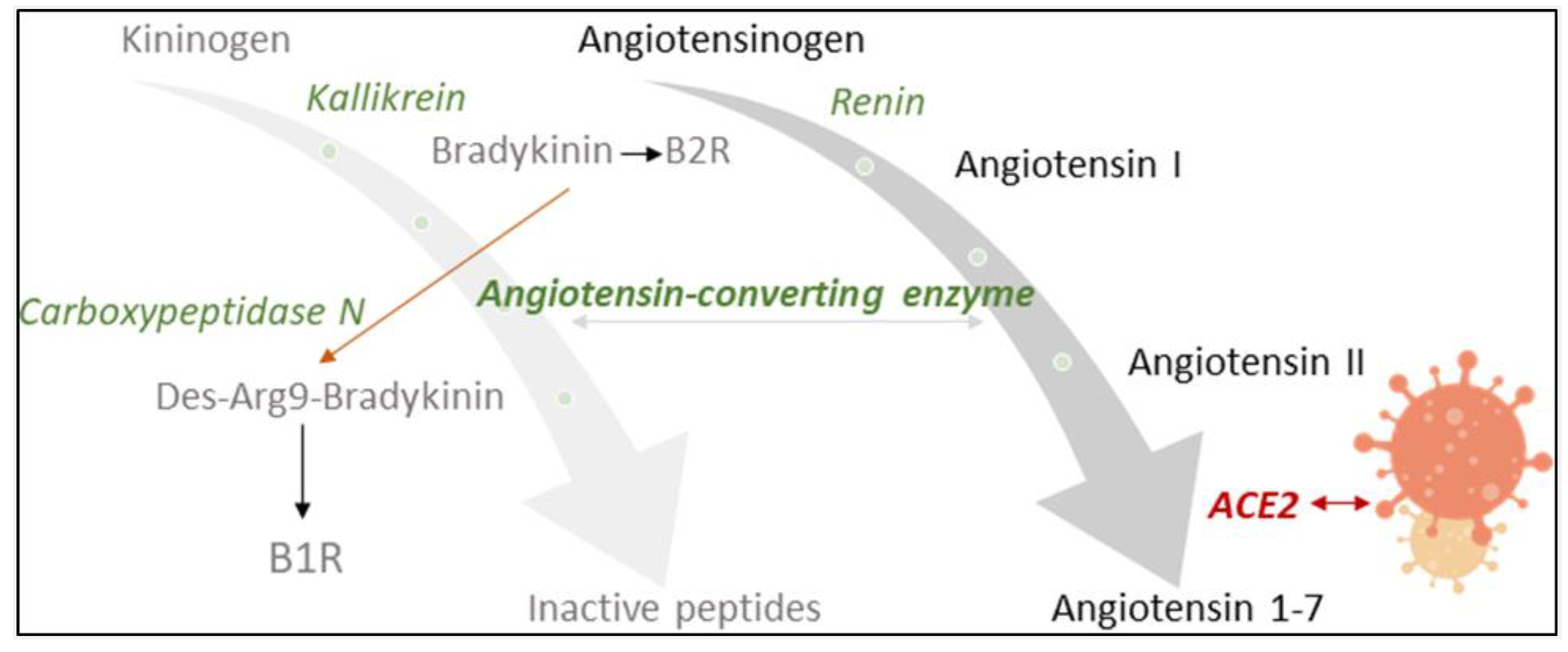
:1. Introduction
2. Materials and Methods
2.1. Participant Selection and Patient Samples
2.2. Laboratory Measurements and Validation of the Clinical Status
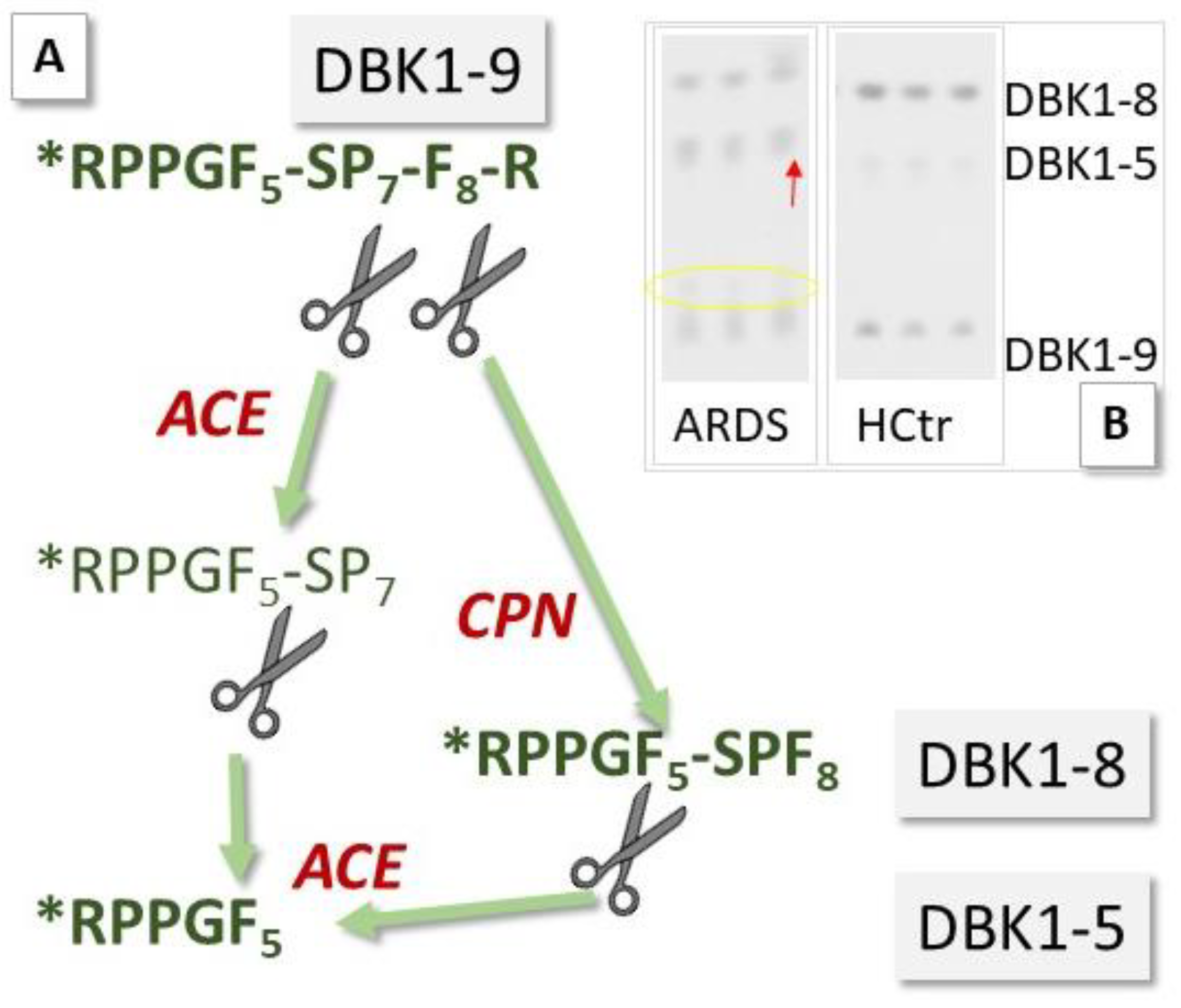
2.3. Neuropeptide Reporter Assay
2.4. Data Analysis/Statistics
3. Results
3.1. Patient Characteristics
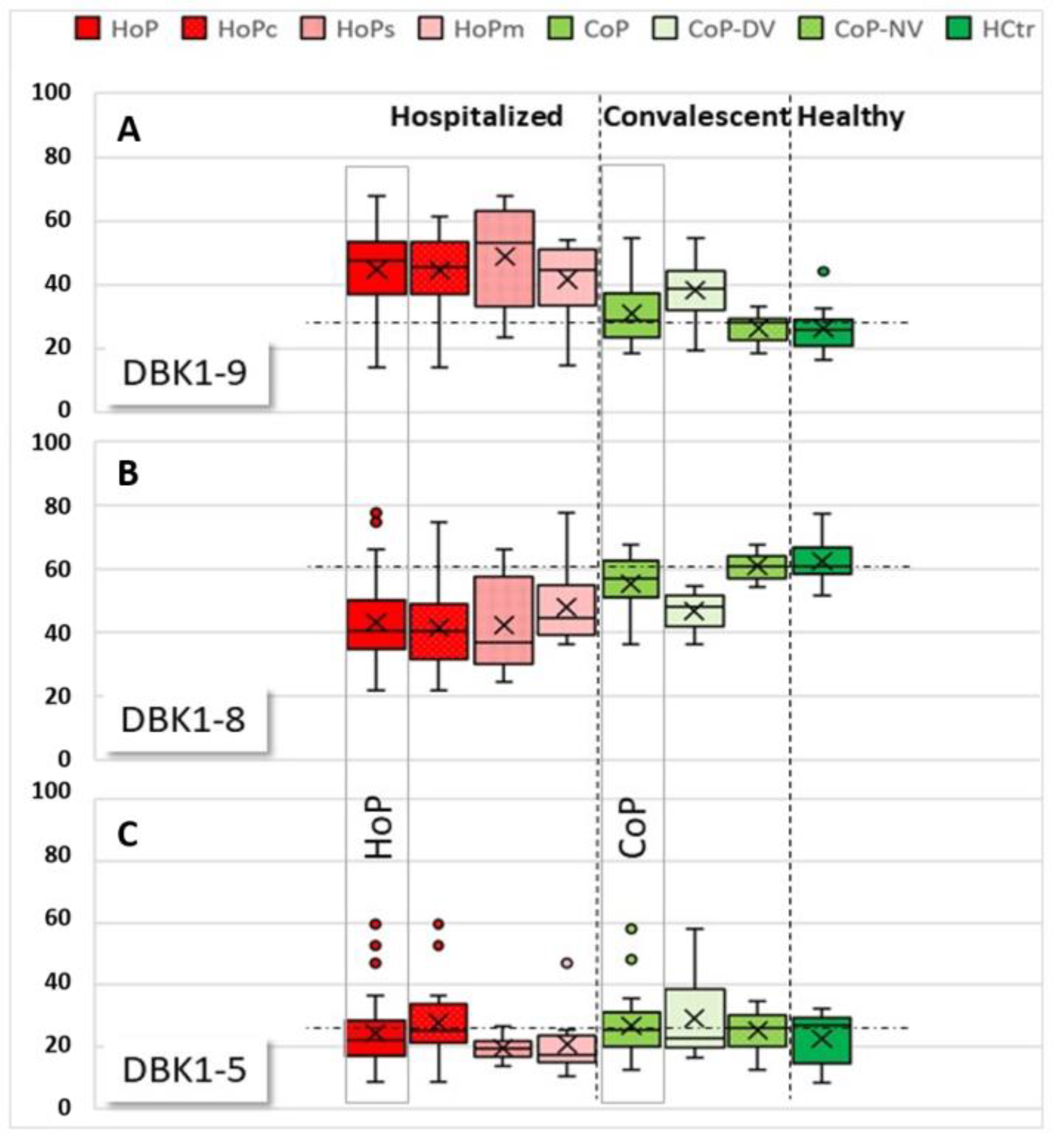
3.2. Neuropeptide Reporter Assay Data
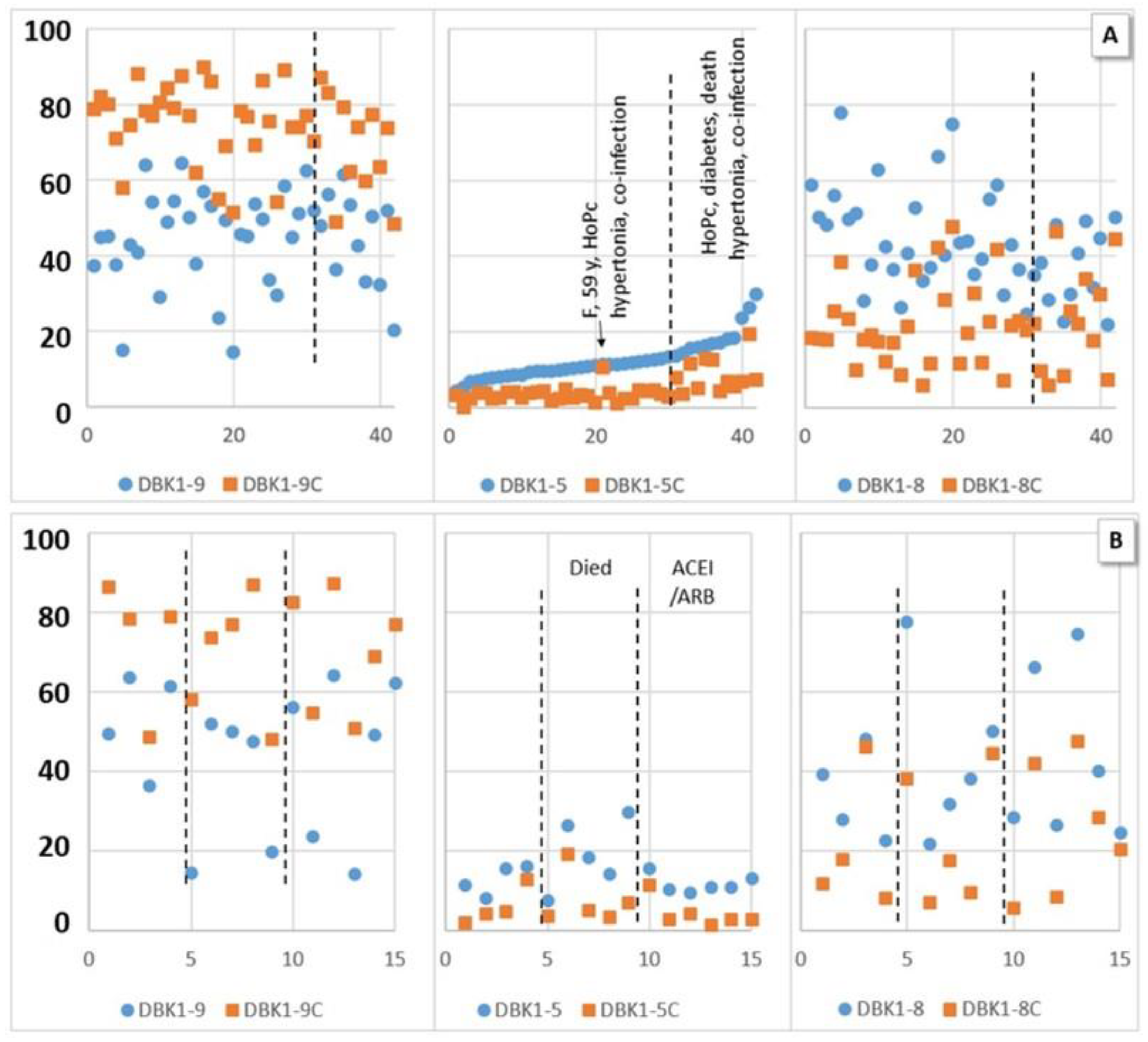
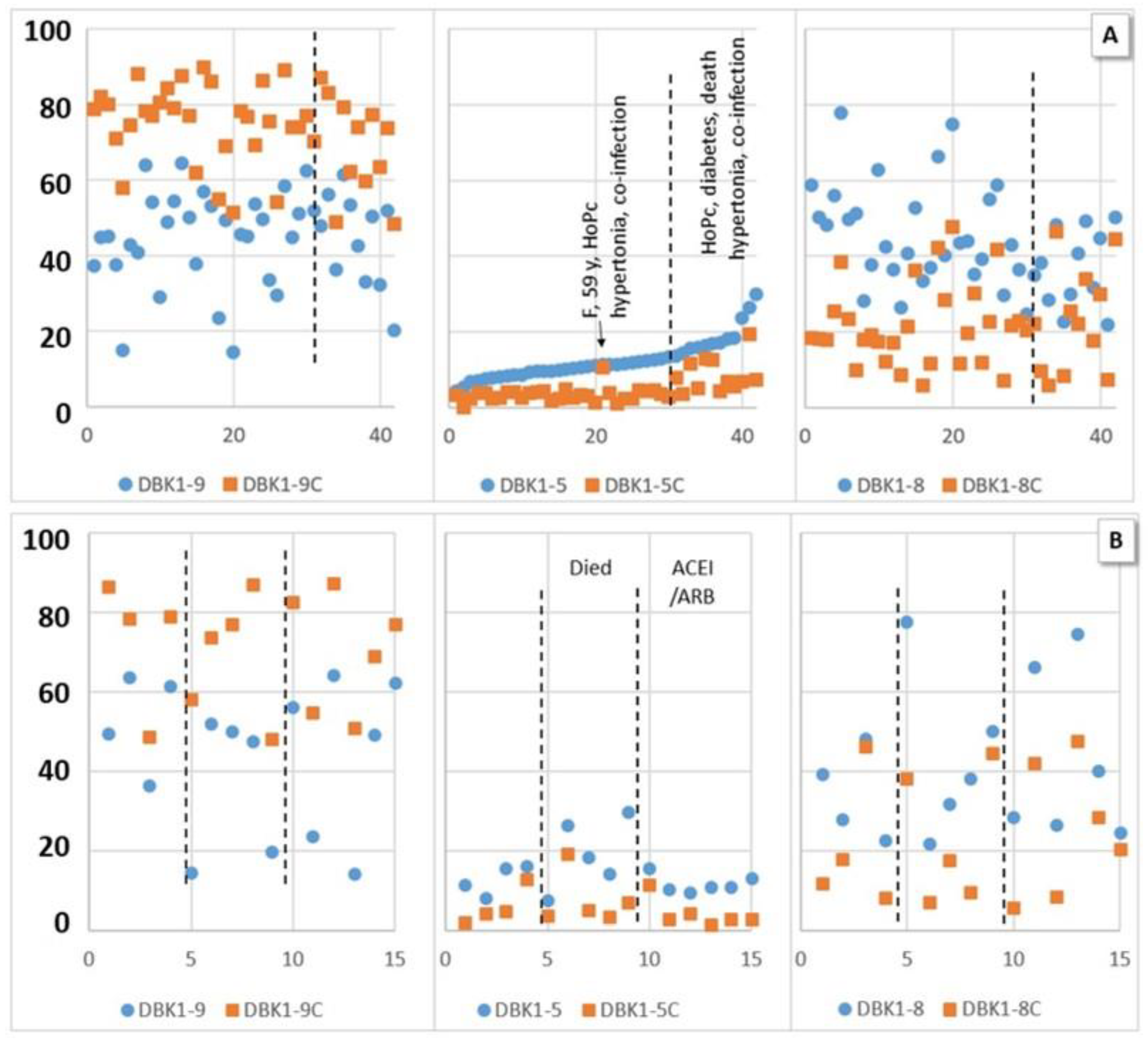
3.2.1. Experiments without Inhibition
3.2.2. Experiments with Inhibition
3.3. Correlations of NRA Values and Clinical Parameters
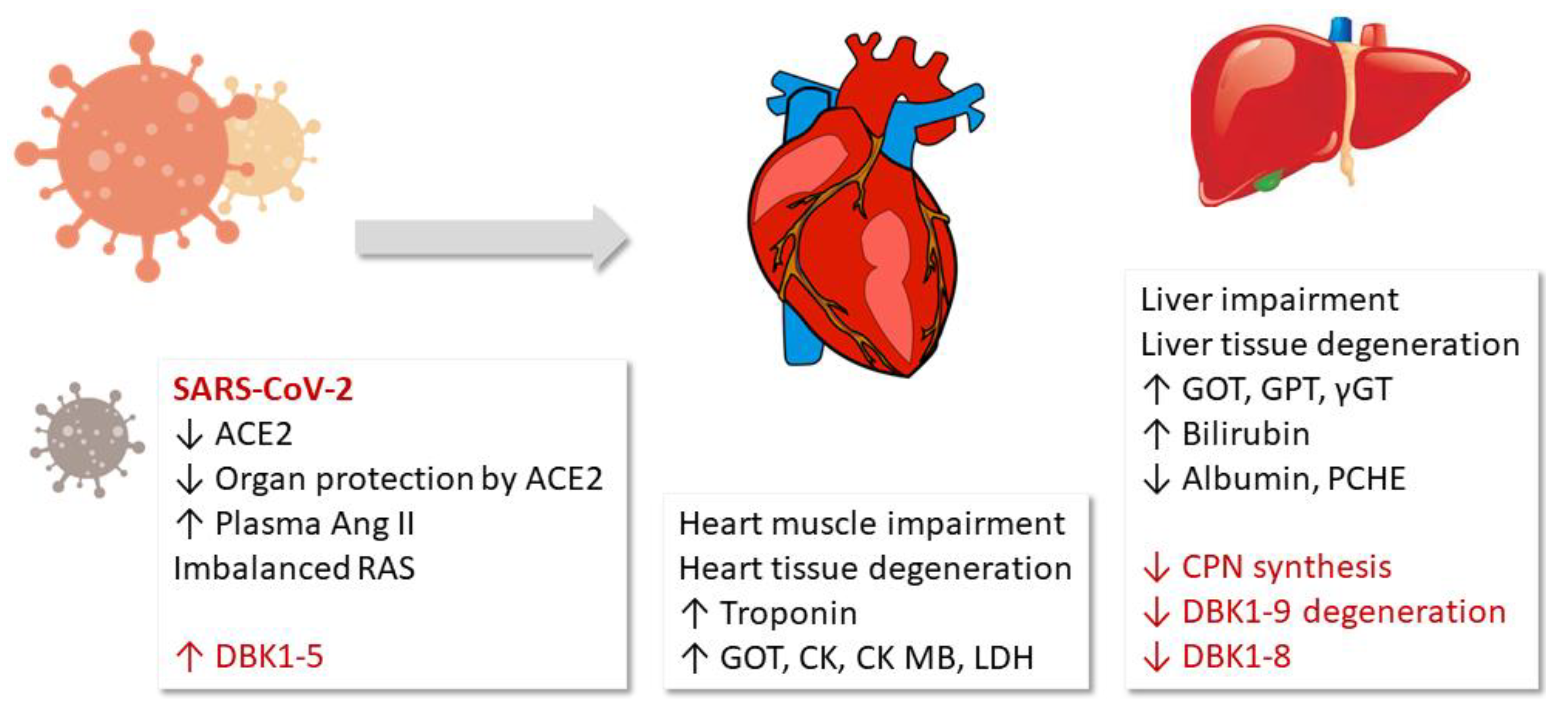
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guang, C.; Phillips, R.D.; Jiang, B.; Milani, F. Three key proteases—Angiotensin-I-converting enzyme (ACE), ACE2 and renin—Within and beyond the renin-angiotensin system. Arch. Cardiovasc. Dis. 2012, 105, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz Ocaranza, M.; Riquelme, J.A.; García, L.; Jalil, J.E.; Chiong, M.; Santos, R.A.S.; Lavandero, S. Counter-regulatory renin–angiotensin system in cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 116–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, R.G. COVID-19—Does this disease kill due to imbalance of the renin angiotensin system (RAS) caused by genetic and gender differences in the response to viral ACE2 attack? Heart Lung Circ. 2020, 29, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Kai, H.; Kai, M. Interactions of coronaviruses with ACE2, angiotensin II, and RAS inhibitors—Lessons from available evidence and insights into COVID-19. Hypertens. Res. 2020, 43, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.; Kali, M.S.K. Association of angiotensin-converting enzyme inhibitors and angiotensin-receptor blockers with risk of mortality, severity or SARS-CoV-2 test positivity in COVID-19 patients: Meta-analysis. Sci. Rep. 2021, 11, 5012. [Google Scholar] [CrossRef]
- Ciaglia, E.; Vecchione, C.; Puca, A.A. COVID-19 infection and circulating ACE2 Levels: Protective role in women and children. Front. Pediatr. 2020, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Skarstein Kolberg, E. ACE2, COVID19 and serum ACE as a possible biomarker to predict severity of disease. J. Clin. Virol. 2020, 126, 104350. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef]
- Bayer, M.; König, S. A vote for robustness: Monitoring serum enzyme activity by thin-layer chromatography of dabsylated bradykinin products. J. Pharmaceut. Biomed. Anal. 2017, 143, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L. Labor und Diagnose, 4th ed.; Medizinische Verlagsges: Marburg, Germany, 1992; pp. 74–76. [Google Scholar]
- Regoli, D.; Barabe, J. Pharmacology of bradykinin and related kinins. Pharm. Rev. 1980, 32, 1–46. [Google Scholar] [PubMed]
- Molinaro, G.; Gervais, N.; Adam, A. Biochemical basis of angioedema associated with recombinant tissue plasminogen activator treatment—An in vitro experimental approach. Stroke 2002, 33, 1712–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, S.; Steinebrey, N.; Herrnberger, M.; Escolano-Lozano, F.; Schlereth, T.; Rebhorn, C.; Birklein, F. Reduced serum protease activity in Complex Regional Pain Syndrome: The impact of angiotensin-converting enzyme and carboxypeptidases. JPBA 2021, 205, 114307. [Google Scholar] [CrossRef] [PubMed]
- König, S.; Bayer, M.; Dimova, V.; Herrnberger, M.; Escolano-Lozano, F.; Bednarik, J.; Vlckova, E.; Rittner, H.; Schlereth, T.; Birklein, F. The serum protease network-one key to understand complex regional pain syndrome pathophysiology. Pain 2019, 160, 1402–1409. [Google Scholar] [CrossRef]
- Birklein, F.; Ajit, S.K.; Goebel, A.; Perez, R.S.G.M.; Sommer, C. Complex regional pain syndrome—Phenotypic characteris-tics and potential biomarkers. Nat. Rev. Neurol. 2018, 14, 272–284. [Google Scholar] [CrossRef]
- König, S.; Schlereth, T.; Birklein, F. Molecular signature for complex regional pain syndrome and its analysis. Expert Rev. Proteom. 2017, 14, 857–867. [Google Scholar] [CrossRef]
- Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute respiratory distress syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar]
- Lucena, J.F.; Alegre, F.; Martinez-Urbistondo, D.; Landecho, M.F.; Huerta, A.; Garcia-Mouriz, A.; Garcia, N.; Quiroga, J. Performance of SAPS II and SAPS 3 in intermediate care. PLoS ONE 2013, 8, e77229. [Google Scholar] [CrossRef]
- Zisman, L.S. ACE and ACE2: A tale of two enzymes. Europ. Heart J. 2005, 26, 322–324. [Google Scholar] [CrossRef]
- Kuoppala, A.; Lindstedt, K.A.; Saarinen, J.; Kovanen, P.T.; Kokkonen, J.O. Inactivation of bradykinin by angiotensin-converting enzyme and by carboxypeptidase N in human plasma. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H1069–H1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteonofrio, L.; Florio, M.C.; AIGhatrif, M.; Lakatta, E.G.; Capogrossi, M.C. Aging- and gender-related mod-ulation of RAAS: Potential implications in COVID-19 disease. Vasc. Biol. 2021, 3, R1–R14. [Google Scholar] [CrossRef] [PubMed]
- Bellis, A.; Mauro, C.; Barbato, E.; Trimarco, B.; Morisco, C. The rationale for angiotensin receptor neprilysin inhibitors in a multi-targeted therapeutic approach to COVID-19. Int. J. Mol. Sci. 2020, 21, 8612. [Google Scholar] [CrossRef] [PubMed]
- Skidgel, R.A. Bradykinin-degrading enzymes: Structure, function, distribution, and potential roles in cardiovascular pharmacology. J. Card. Pharm. 1992, 20 (Suppl. 9), S4–S9. [Google Scholar] [CrossRef]
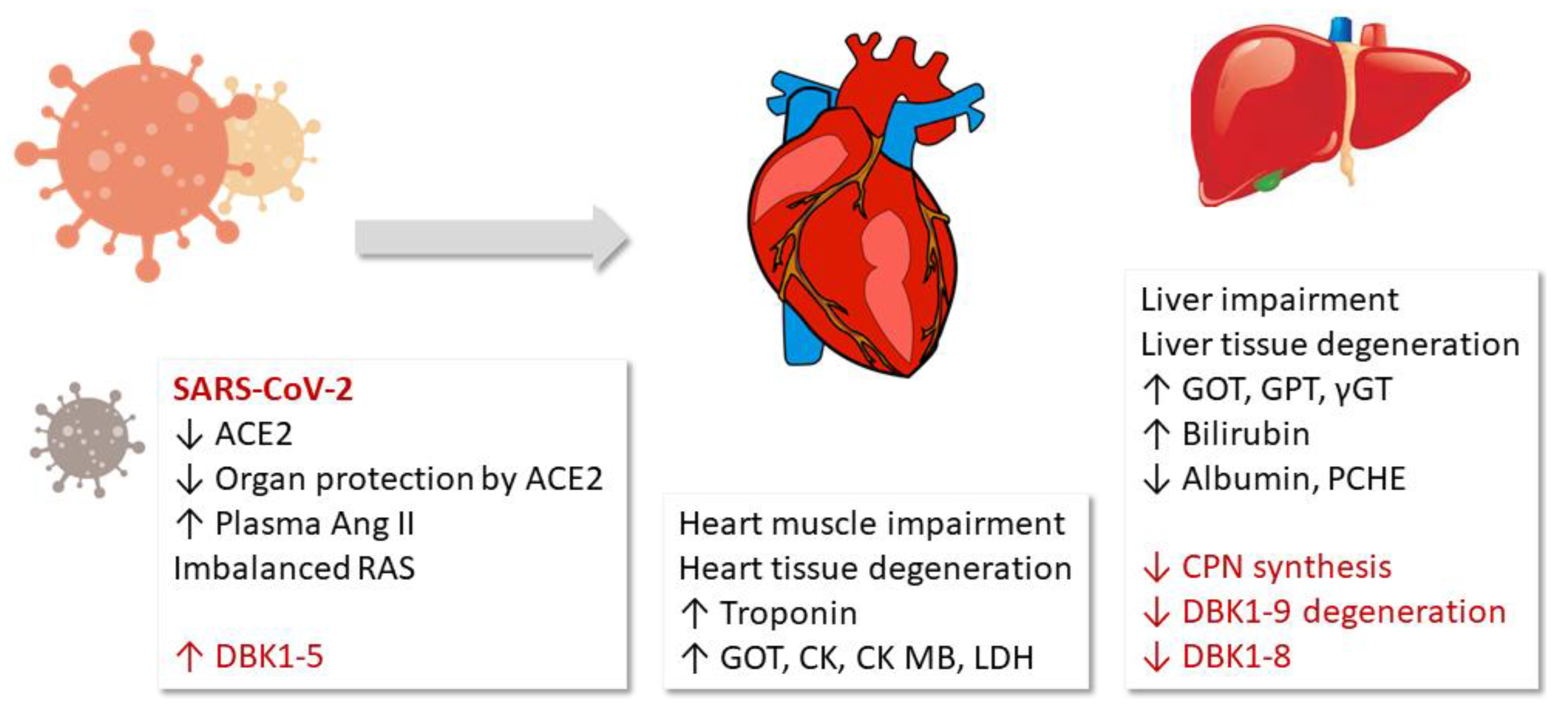

{kind=link}
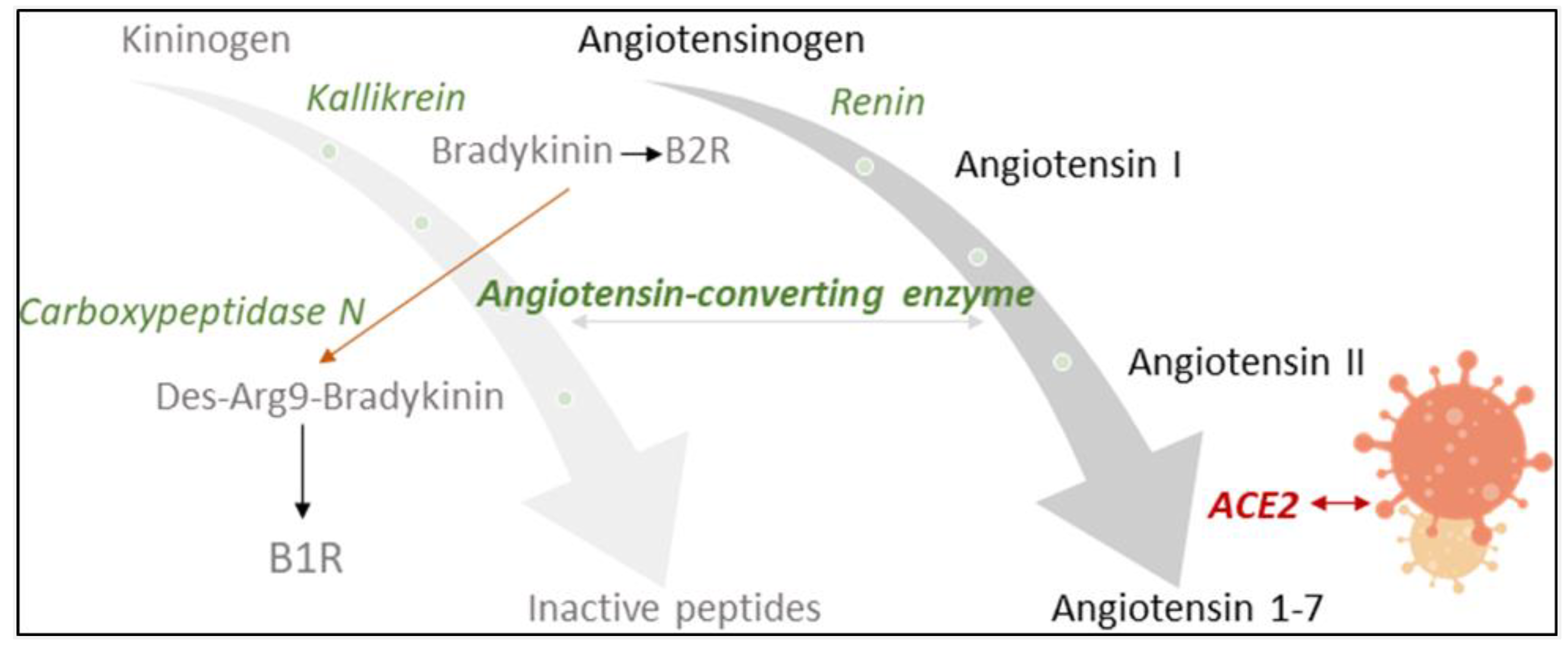
{kind=link}
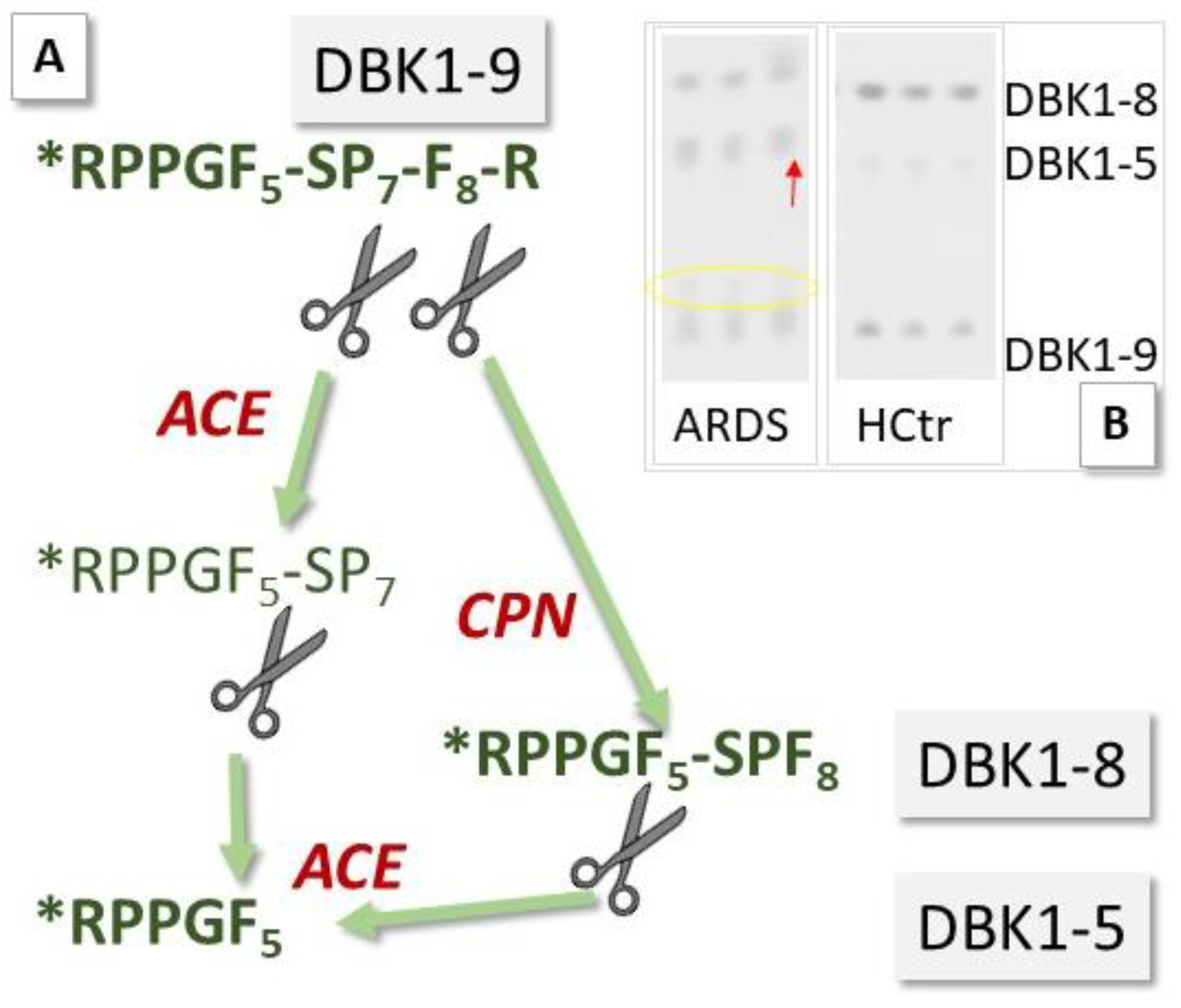
{kind=link}
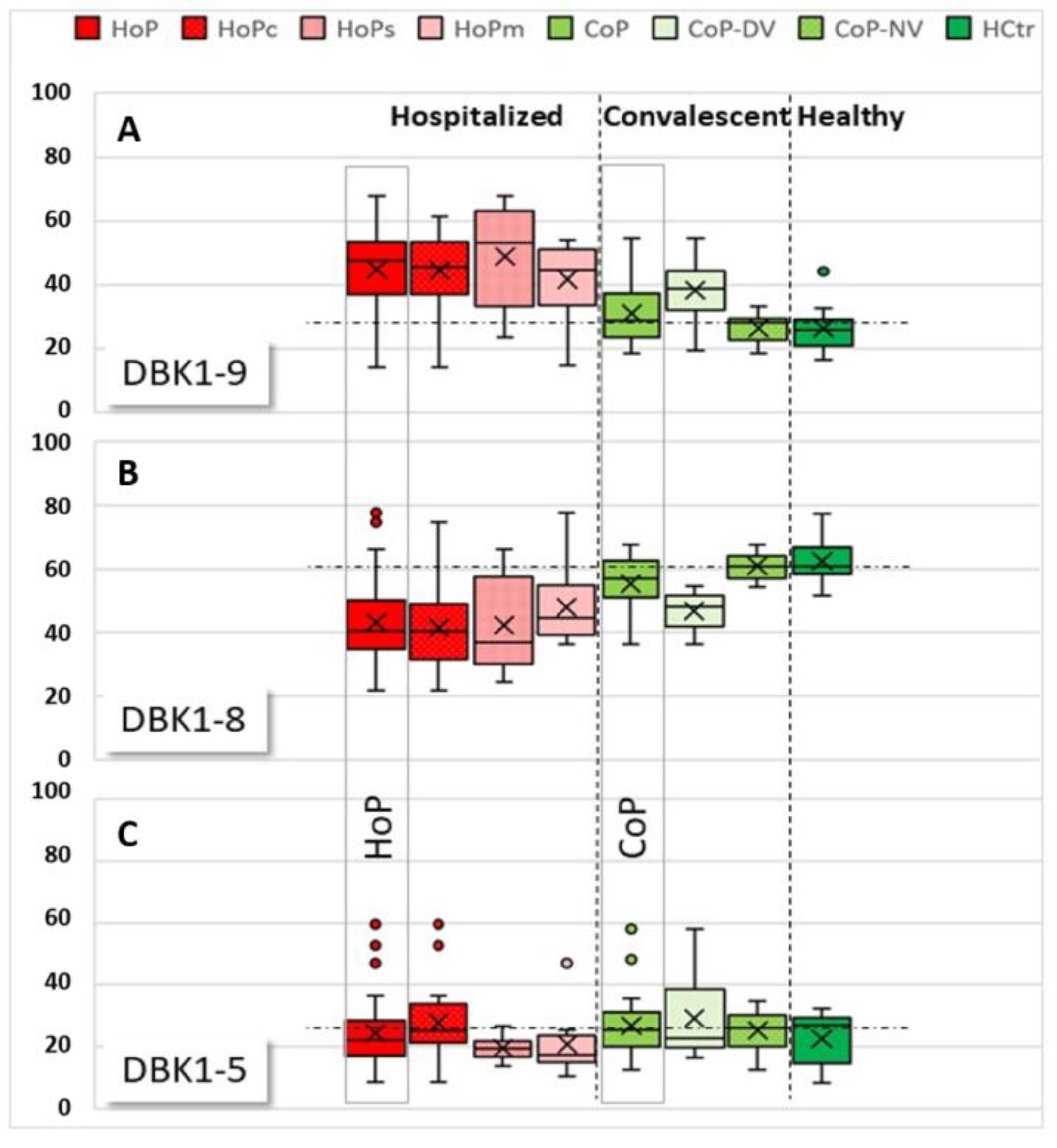
{kind=link}
{kind=link}
| Patients (Samples) | HoP | CoP | ||||||
|---|---|---|---|---|---|---|---|---|
| HoPc (n = 22/45) | HoPs (n = 12/45) | HoPm (n = 11/45) | p-Value | CoP (n = 26) | CoP-NV (n = 16/26) | HCtr (n = 8) | ||
| Patient Characteristics | Age, years median (IQR) | 57 (49–63) | 57 (49–66) | 55 (38–65) | 0.516 | 62 (55–70) | 60 (55–65) | 31 (25–38) |
| Gender, male, n (%) | 22 (100) | 19 (86) | 9 (82) | 24 (92) | 15 (94) | 5 (63) | ||
| BMI | 27.3 (25–31.2) | 23 (22.8–25.2) | 23.8 (22.8–27.7) | 0.005 | n/a | n/a | n/a | |
| Death (abs.) | 2 | 7 | 2 | 0 | 0 | 0 | ||
| SARS CoV-2 therapy | Cortison | 9 | 0 | 2 | 0 | 0 | 0 | |
| Pre-existing conditions | Chronic inflammatory disease (abs.) | 0 | 3 | 1 | 0 | 0 | 0 | |
| Respiratory disease (abs.) | 0 | 1 | 0 | 0 | 0 | 0 | ||
| Kidney insufficiency (abs.) | 0 | 0 | 1 | 0 | 0 | 0 | ||
| Metastatic neoplasm (abs.) | 2 | 1 | 3 | 0 | 0 | 0 | ||
| Diabetes (abs.) | 0 | 3 | 0 | 0 | 0 | 0 | ||
| Arterial hypertension (abs.) | 4 | 10 | 5 | 0 | 0 | 0 | ||
| Coronary heart disease (abs.) | 1 | 3 | 2 | 0 | 0 | 0 | ||
| Medication | Angiotensin-1 receptor antagonist (abs.) | 2 | 1 | 0 | 0 | 0 | 0 | |
| Angiotensin converting enzyme inhibitor (abs.) | 1 | 2 | 1 | 0 | 0 | 0 | ||
| HoP (n = 45) | p-Value | CoP (n = 26) | |||
|---|---|---|---|---|---|
| HoPc (n = 22) | HoPs (n = 12) | HoPm (n = 11) | |||
| Erythrocytes (Mio./µL) | 3.4 (2.7–4.1) | 4.8 (3.9–5.1) | 4.7 (4.1–5.1) | p < 0.001 | 4.8 (4.6–5.2) |
| Hemoglobin (g/dL) | 9.4 (7.9–11.6) | 13.7 (11.9–15) | 13.6 (11.8–14.9) | p < 0.001 | 14.6 (13.8–15.7) |
| Leukocytes (×109/L) | 9.1 (6.8–11.4) | 5.3 (3.7–7) | 4.1 (3.5–5.9) | p = 0.001 | 5.5 (4.96–6.9) |
| Lymphocytes (rel., %) | 10.5 (7.2–14.1) | 21.3 (13.1–22.5) | 22.1 (14.6–21.1) | p = 0.001 | 28.2 (24.8–31.4) |
| AST (U/L) | 71 (45.8–103) | 41 (34–75) | 38.5 (27–53.5) | p = 0.008 | 30.5 (24.3–34.8) |
| ALT (U/L) | 42.5 (28.3–67) | 34 (28–43) | 25.5 (18.3–46.8) | p = 0.204 | 28.5 (24.5–34.8) |
| ɣ-GT (U/L) | 127 (60–204) | 54 (26–60) | 42 (26–134.5) | p = 0.003 | 27.5 (21–50) |
| PCHE (U/L) | 3457 (2749–5026) | 6291 (4175–7998) | 7695 (5806–8711) | p < 0.001 | 8956 (7948–10408) |
| Bilirubin (mg/dL) | 0.6 (0.4–1.3) | 0.5 (0.3–0.6) | 0.4 (0.3–0.75) | p = 0.107 | 0.5 (0.4–0.7) |
| LDH (E/L) | 424 (329–543) | 392 (264–418) | 268.5 (229.5–412) | p = 0.005 | 217.5 (194.3–239) |
| Creatinine (mg/dL) | 1 (0.68–1.8) | 0.8 (0.7–1.1) | 1 (0.8–1) | p = 0.69 | 0.9 (0.8–1) |
| Urea (mg/dL) | 19.5 (13–51) | 11 (9–22) | 11 (9–23.3) | p = 0.065 | 14 (13–16.5) |
| CK (U/L) | 148.5 (57–394) | 132 (46–206) | 81 (59–186) | p = 0.515 | 131.5 (78.5–148) |
| CK-MB (U/L), | 20 (16–27) | 15 (8.8–17.8) | 11 (5–19) | p = 0.012 | 5 (5–5) |
| Troponin T (ng/L) | 32 (15.5–118) | 12.5 (6.4–23) | 8.5 (5.3–18.6) | p = 0.003 | n.d. |
| D-Dimer (mg/L) | 2.6 (1.9–8.7) | 0.8 (0.7–1.4) | 0.7 (0.4–1.5) | p < 0.001 | n.d. |
| CRP (mg/dL) | 14.8 (5.5–24.8) | 6 (3.7–9.4) | 1.8 (0.8–3.3) | p < 0.001 | 0.5 (0.5–0.5) |
| Ferritin (µg/L) | 898 (685–1561) | 692 (358–1065) | 371 (197–617) | p < 0.001 | 200.5 (92–402.8) |
| Procalcitonin (ng/mL) | 0.53 (0.16–2.04) | 0.11 (0.07–0.25) | 0.09 (0.06–0.13) | p < 0.001 | 0.05 (0.04–0.07) |
| SAPS-II Score | 56 (34–73) | 22 (15–32) | 17 (13–24) | p < 0.001 | n.d. |
| DBK1-9 | DBK1-9C | DBK1-9Diff | DBK1-8 | DBK1-8C | DBK1-8Diff | DBK1-5 | DBK1-5C | DBK1-5Diff | |
|---|---|---|---|---|---|---|---|---|---|
| SAPS II | −0.387 * | −0.339 * | 0.289 * | −0.371 * | 0.519 ** | 0.526 ** | |||
| Troponin | −0.313 * | −0.538 ** | 0.436 ** | −0.347 * | 0.485 ** | 0.373 * | |||
| Erythrocytes | 0.285 * | −0.343 | |||||||
| Hb | 0.296 * | 0.304 * | 0.317 * | −0.337 * | |||||
| Ferritin | 0.346 * | −0.542 ** | −0.392 ** | −0.476 ** | 0.411 ** | ||||
| GPT | −0.346 * | −0.454 ** | −0.503 ** | 0.350 * | 0.317 * | ||||
| GOT | −0.357 * | −0.664 ** | −0.403 ** | −0.654 ** | 0.389 ** | 0.562 ** | |||
| γGT | −0.283 * | −0.338 * | 0.499 ** | 0.318 * | |||||
| PCHE | 0.291 * | 0.363 * | −0.343 * | ||||||
| Bilirubin | −0.294 * | 0.578 ** | 0.302 * | 0.451 ** | |||||
| Albumin | −0.299 * | 0.541 ** | 0.383 ** | 0.499 ** | −0.441 ** | ||||
| PCT | −0.340 * | 0.296 * | |||||||
| LDH | 0.291 * | −0.468 ** | −0.358 * | 0.280 * | |||||
| CK-MB | 0.384 ** | −0.586 ** | −0.429 ** | −0.468 ** | 0.586 ** | −0.330 * | |||
| CRP | −0.429 ** | −0.325 * | −0.383 ** | 0.451 ** | |||||
| Creatinine | −0.465 ** | 0.429 ** | |||||||
| Lymphocytes, relative | −0.287 * | 0.385 ** | 0.289 * | −0.371 ** | 0.303 * | ||||
| Lymphocytes, absolute | −0.356 * | 0.282 * | |||||||
| D-Dimers | 0.333 * | ||||||||
| Leukocytes | 0.490 ** | ||||||||
| Urea | 0.317 * | ||||||||
| CK | 0.495 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tepasse, P.-R.; Vollenberg, R.; Steinebrey, N.; König, S. High Angiotensin-Converting Enzyme and Low Carboxypeptidase N Serum Activity Correlate with Disease Severity in COVID-19 Patients. J. Pers. Med. 2022, 12, 406. https://doi.org/10.3390/jpm12030406
Tepasse P-R, Vollenberg R, Steinebrey N, König S. High Angiotensin-Converting Enzyme and Low Carboxypeptidase N Serum Activity Correlate with Disease Severity in COVID-19 Patients. Journal of Personalized Medicine. 2022; 12(3):406. https://doi.org/10.3390/jpm12030406
Chicago/Turabian StyleTepasse, Phil-Robin, Richard Vollenberg, Nico Steinebrey, and Simone König. 2022. "High Angiotensin-Converting Enzyme and Low Carboxypeptidase N Serum Activity Correlate with Disease Severity in COVID-19 Patients" Journal of Personalized Medicine 12, no. 3: 406. https://doi.org/10.3390/jpm12030406
APA StyleTepasse, P.-R., Vollenberg, R., Steinebrey, N., & König, S. (2022). High Angiotensin-Converting Enzyme and Low Carboxypeptidase N Serum Activity Correlate with Disease Severity in COVID-19 Patients. Journal of Personalized Medicine, 12(3), 406. https://doi.org/10.3390/jpm12030406