
Integrated Proteomics Based on 2D Gel Electrophoresis and Mass Spectrometry with Validations: Identification of a Biomarker Compendium for Oral Submucous Fibrosis—An Indian Study

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Tissue Samples
2.2. Protein Extraction from Tissue Samples
2.3. 2D Gel Electrophoresis
2.4. In-Gel Trypsin Digestion and MALDI-TOF
2.5. Pathway Analysis and Gene Ontology
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
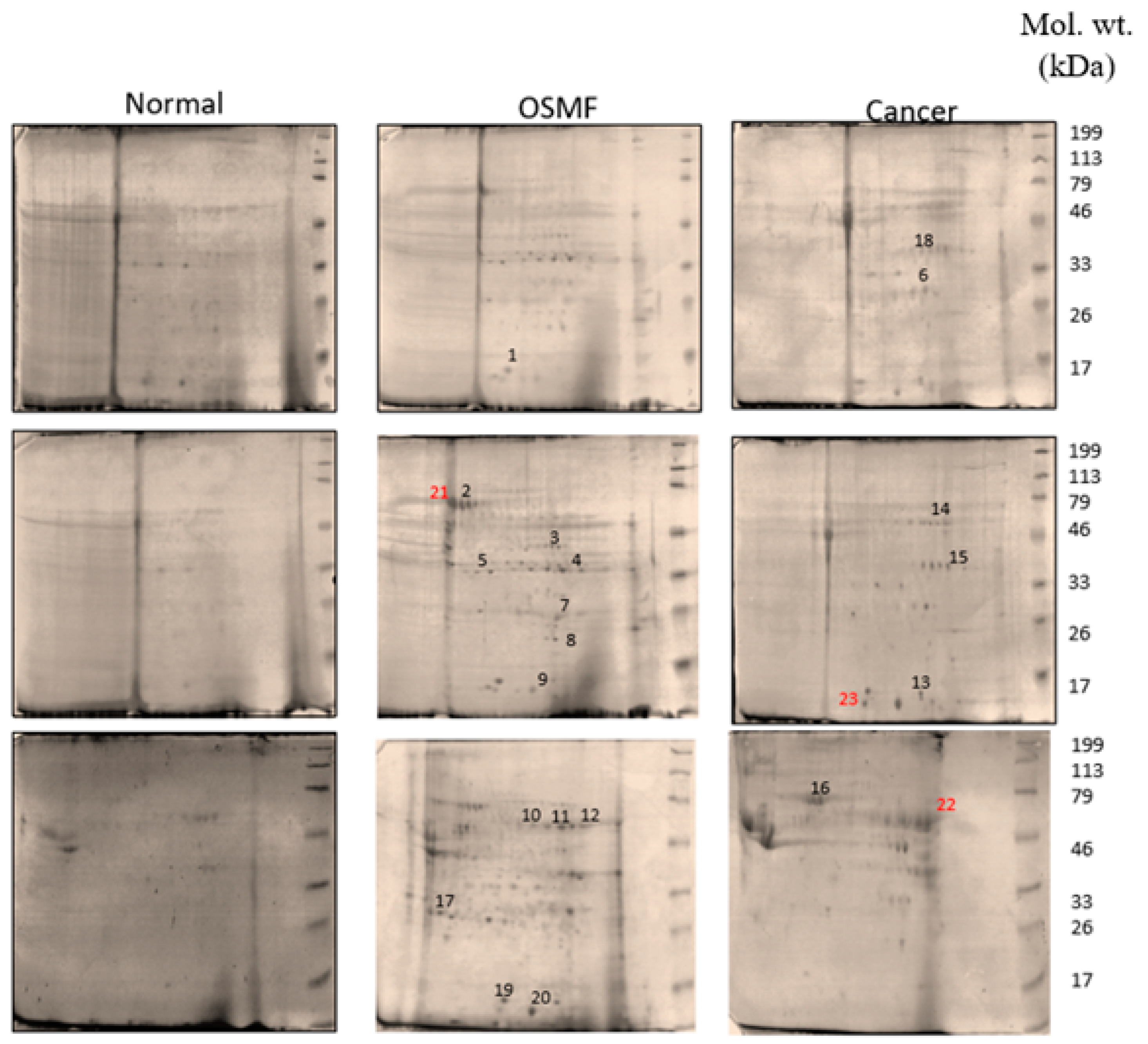
3.1. Quantitative Protein Profiling Using 2D Gel Electrophoresis and MALDI-TOF
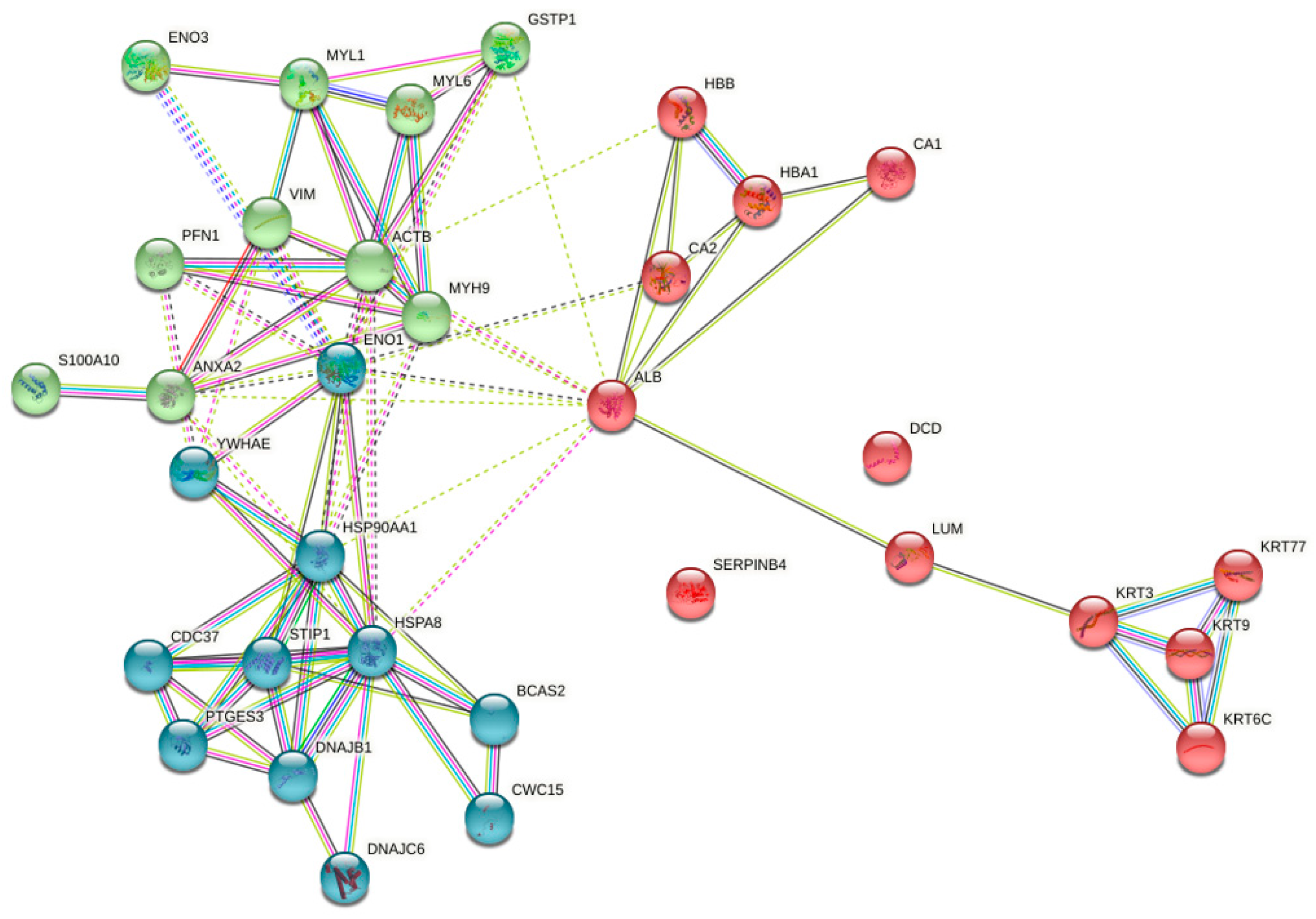
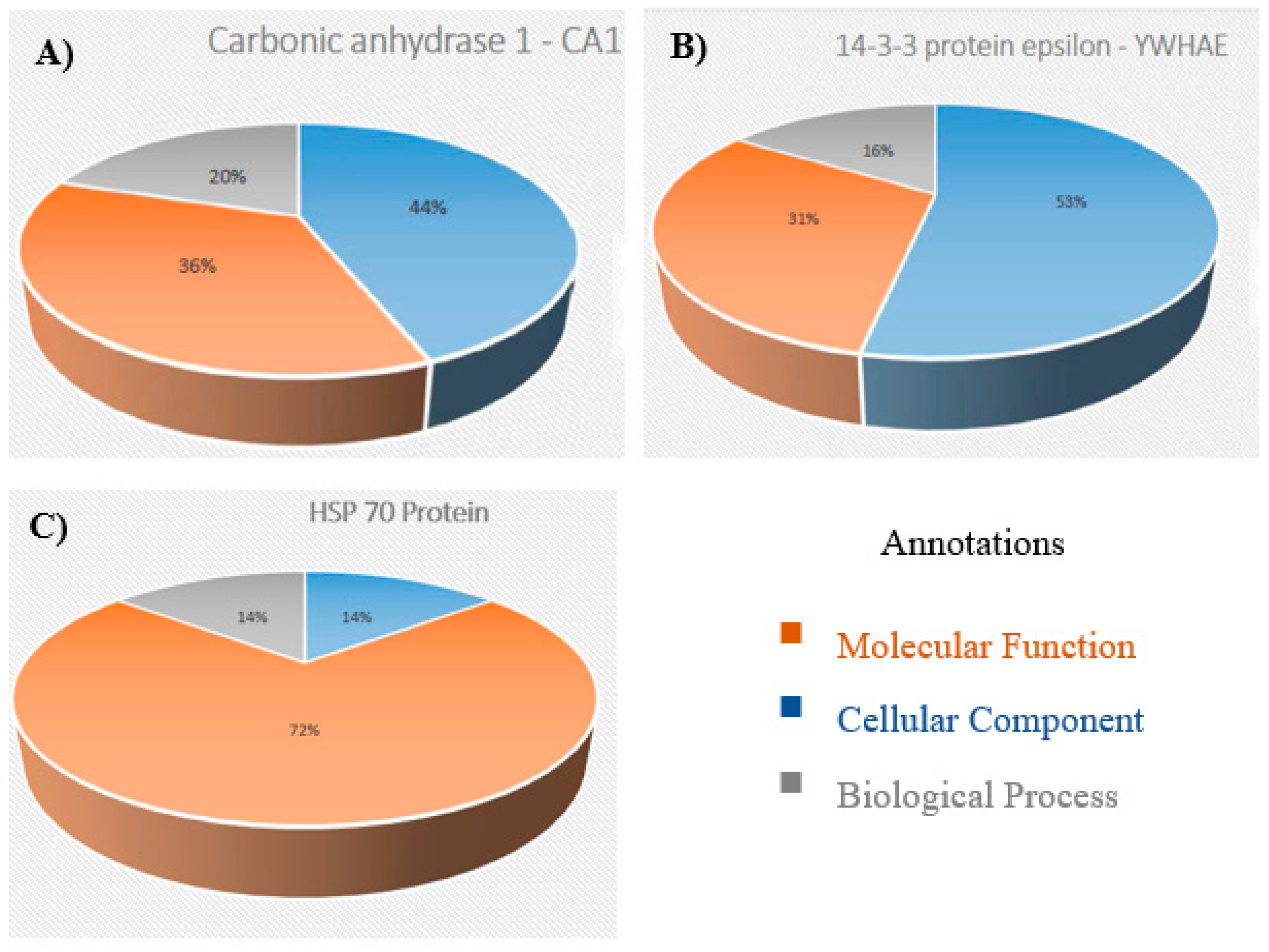
3.2. Functional Classification of Identified Proteins and Biological Network Analysis
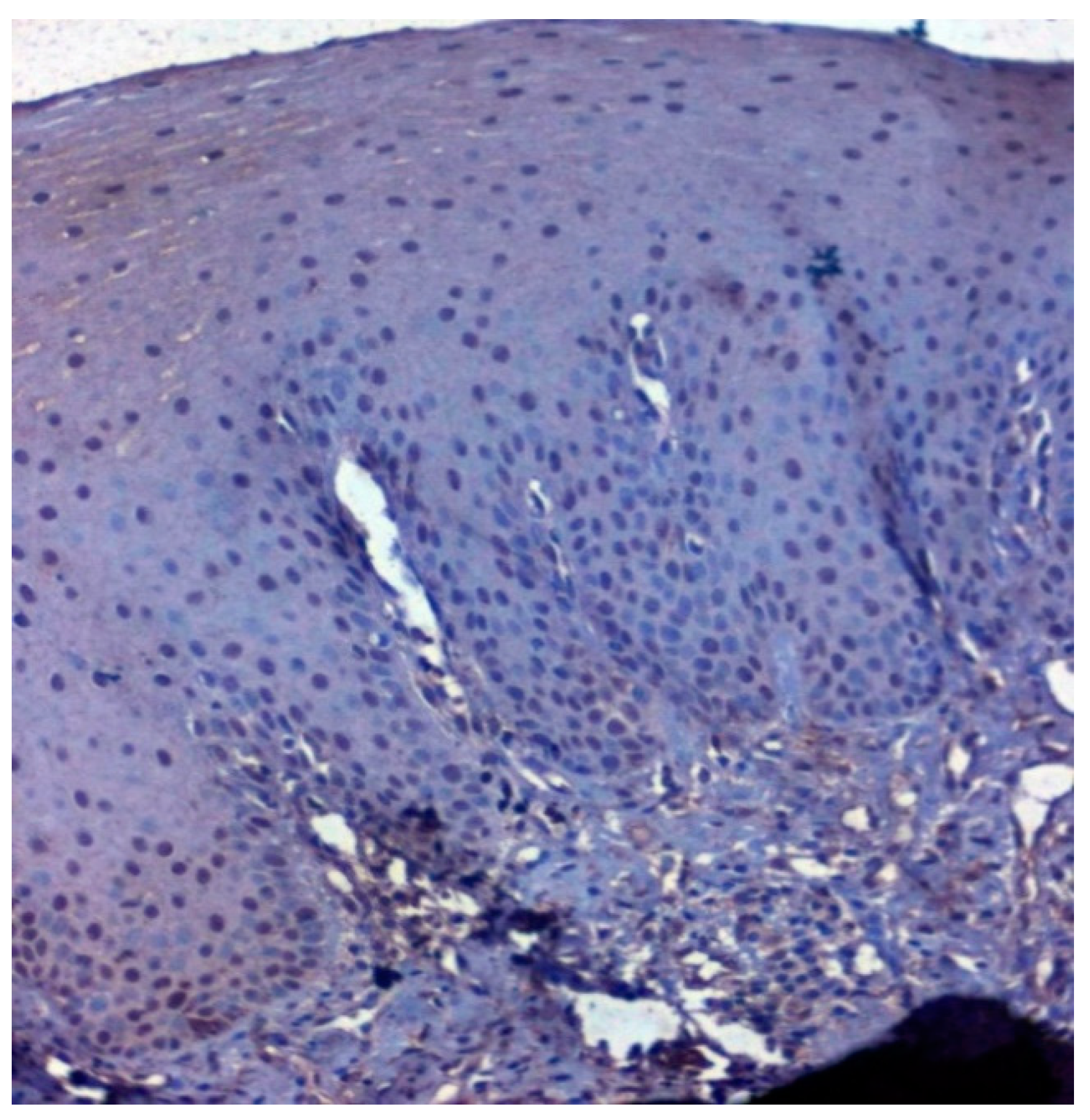
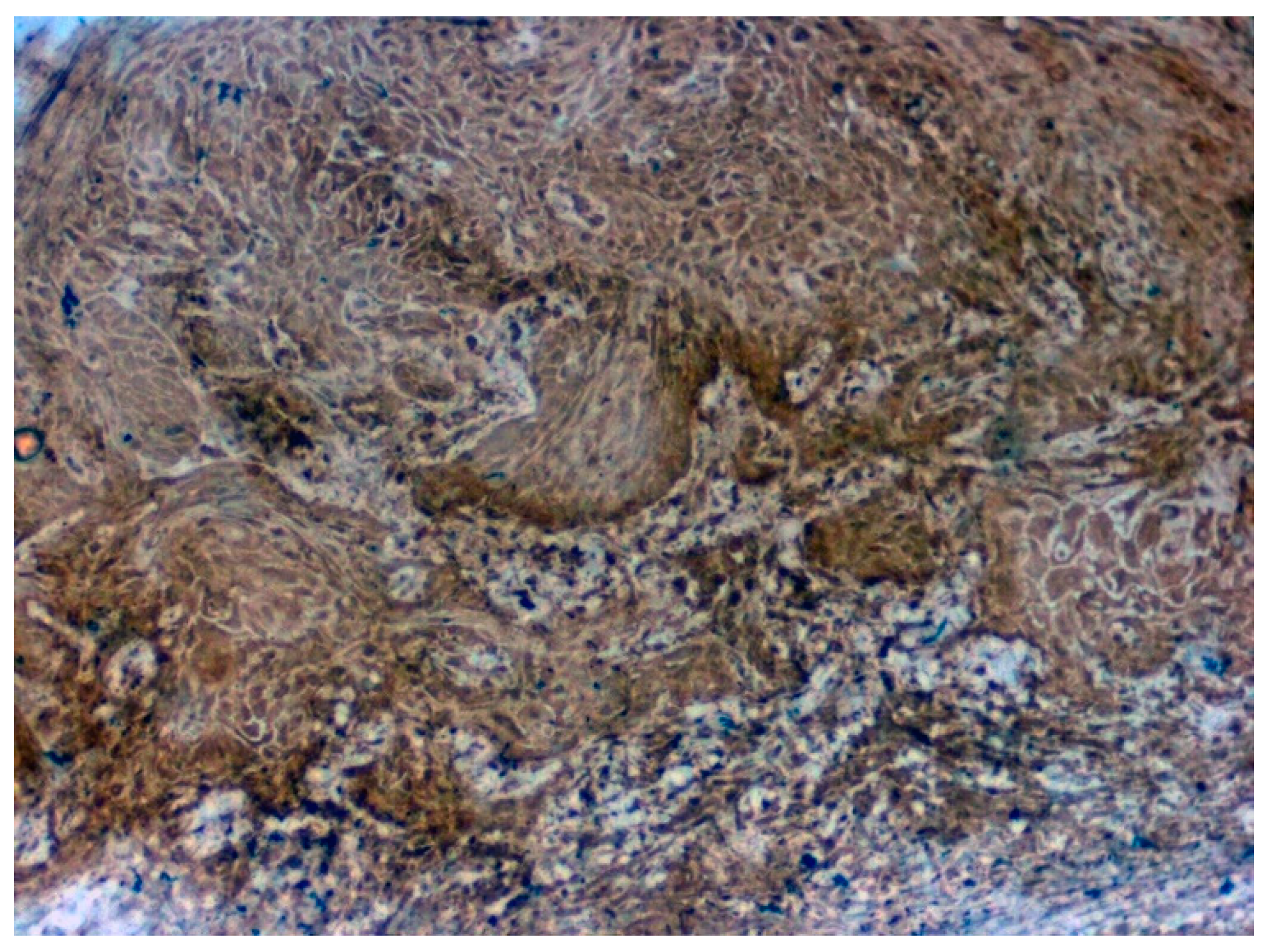
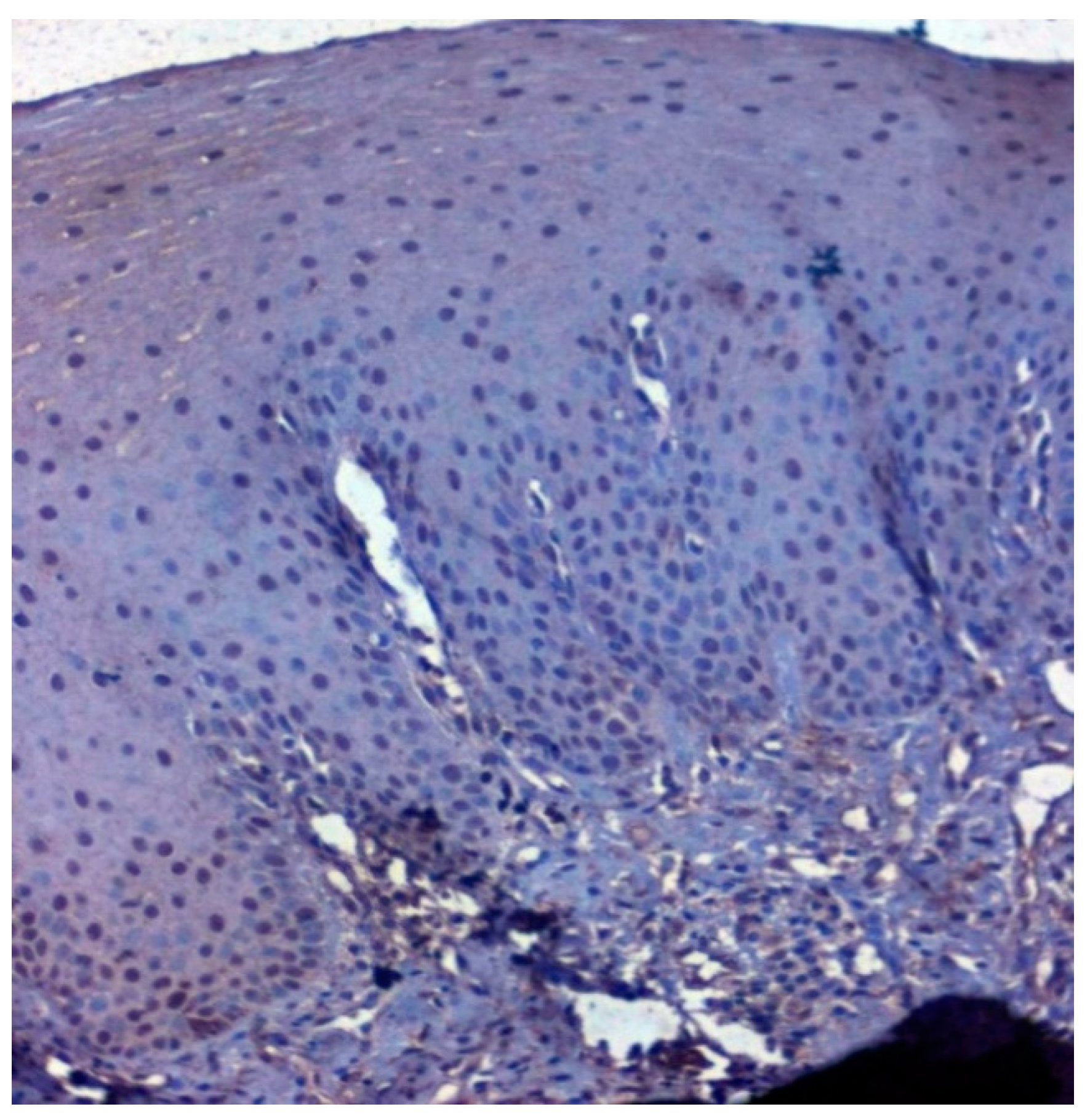
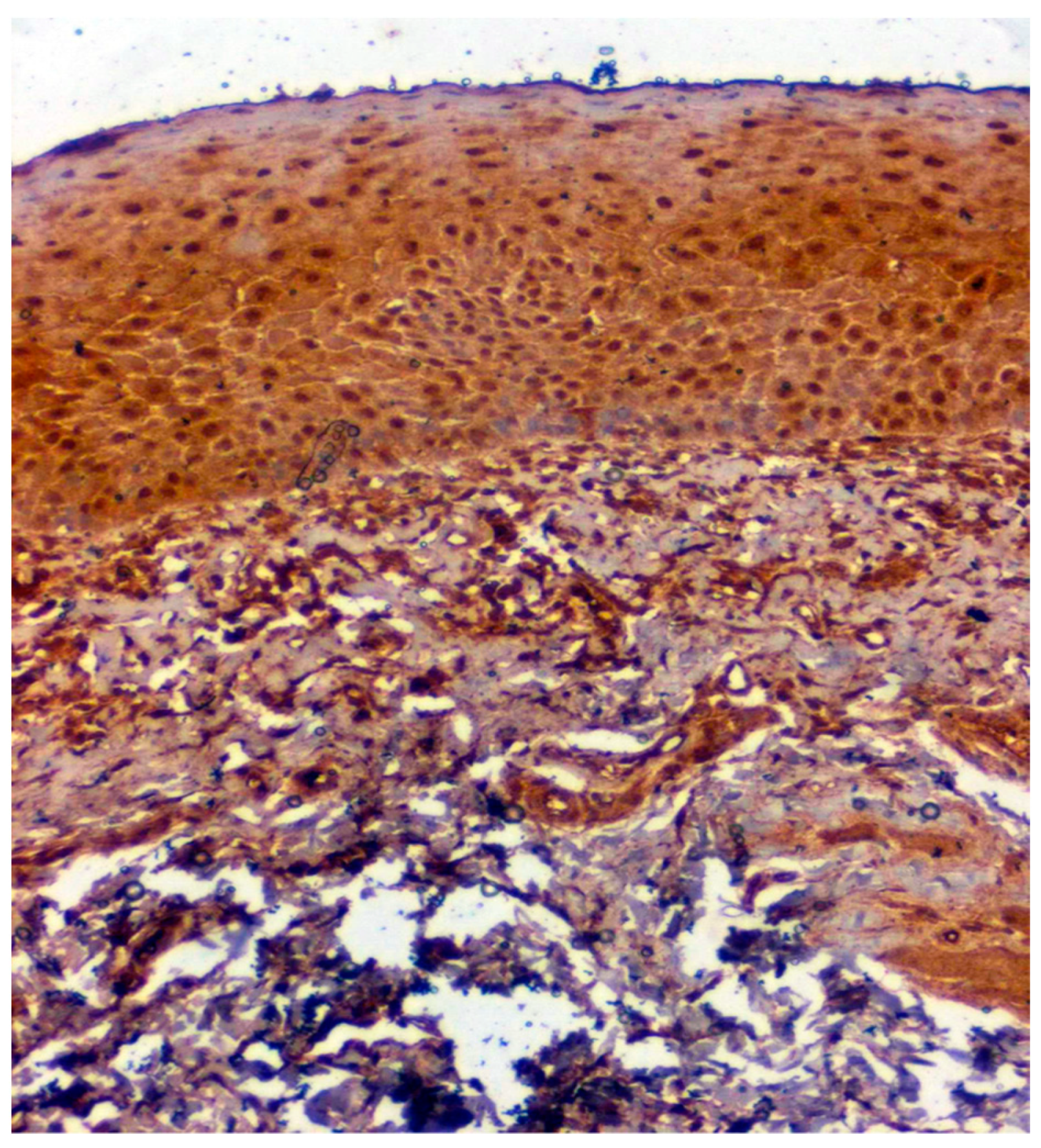
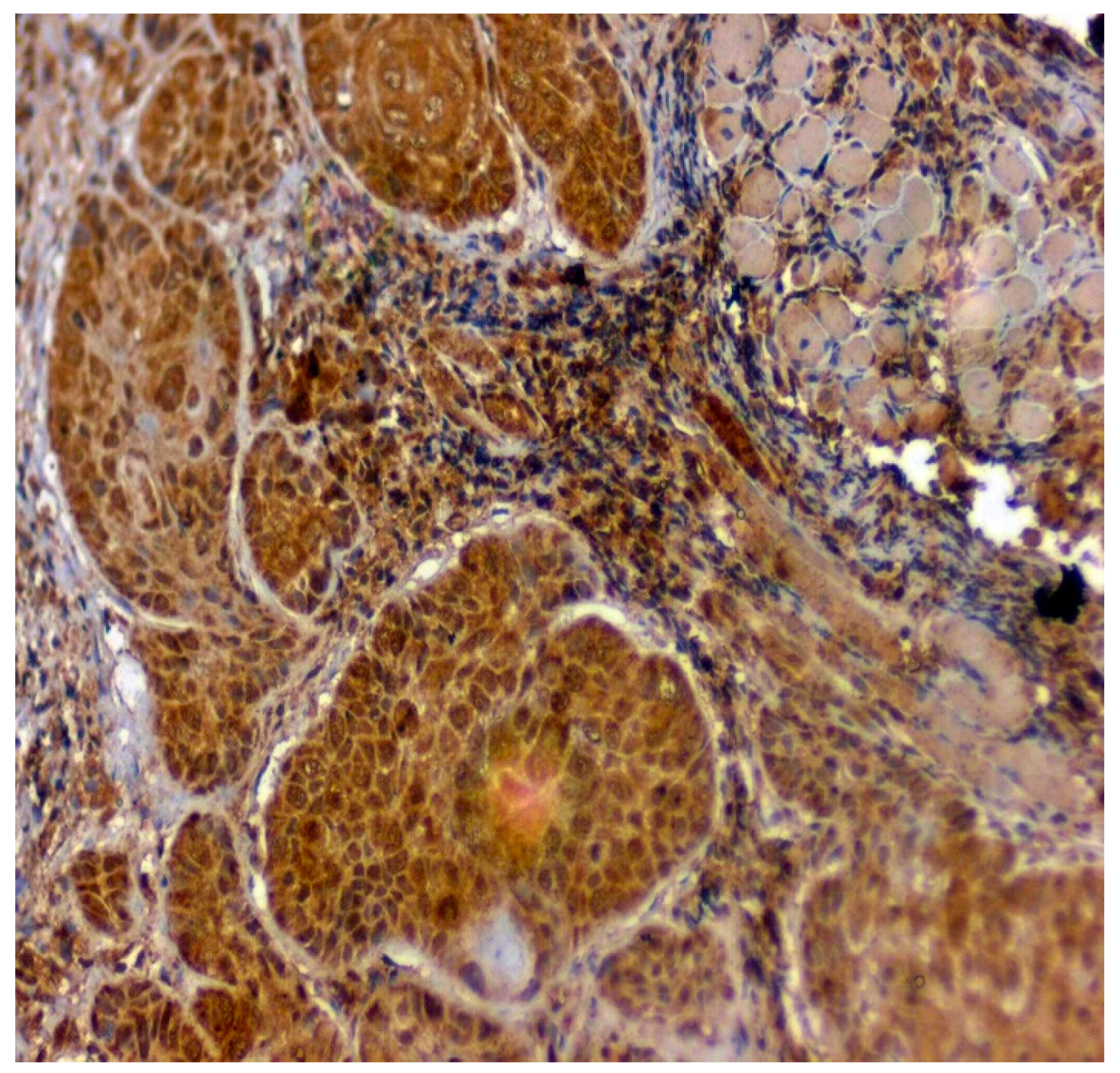
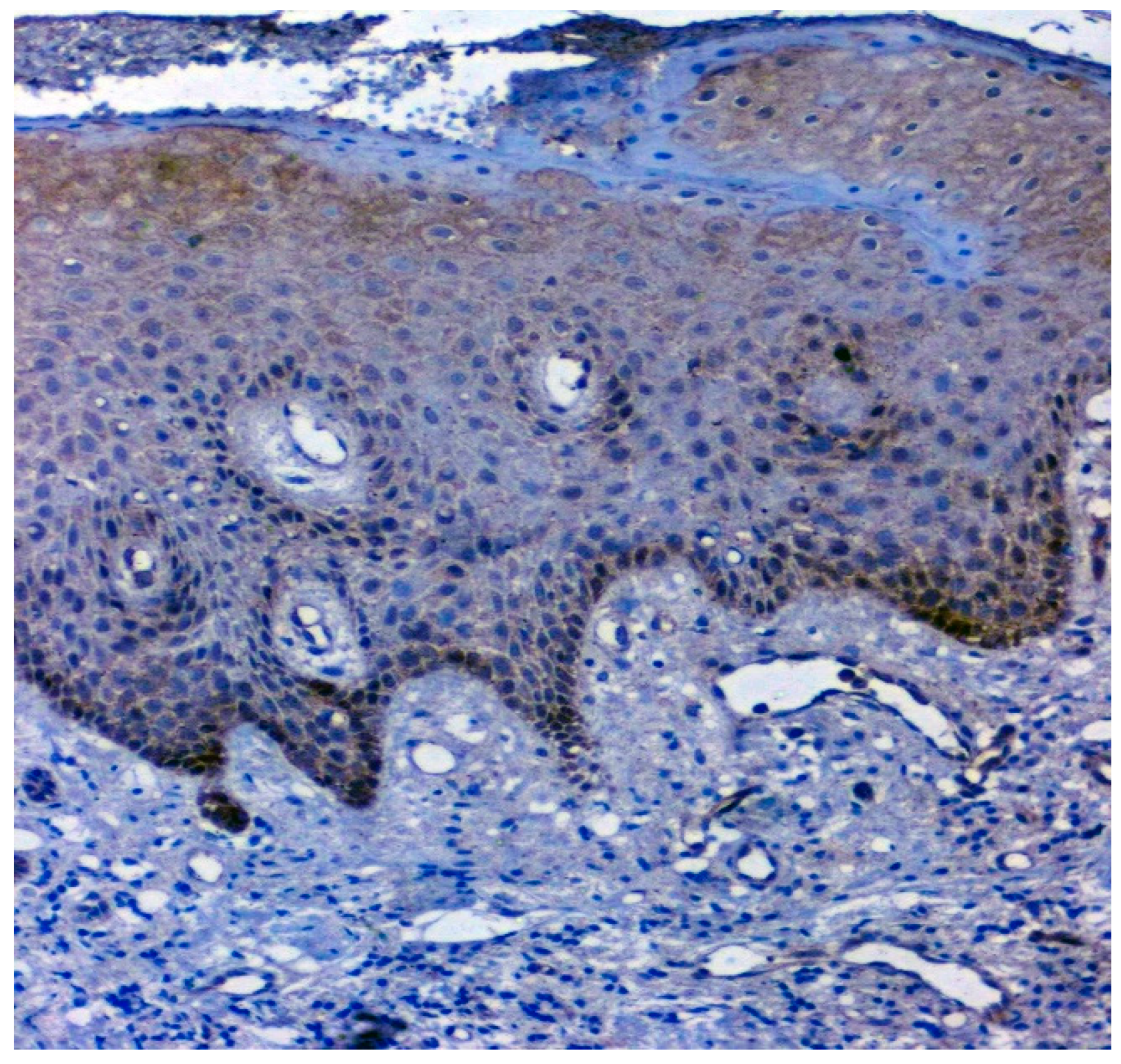
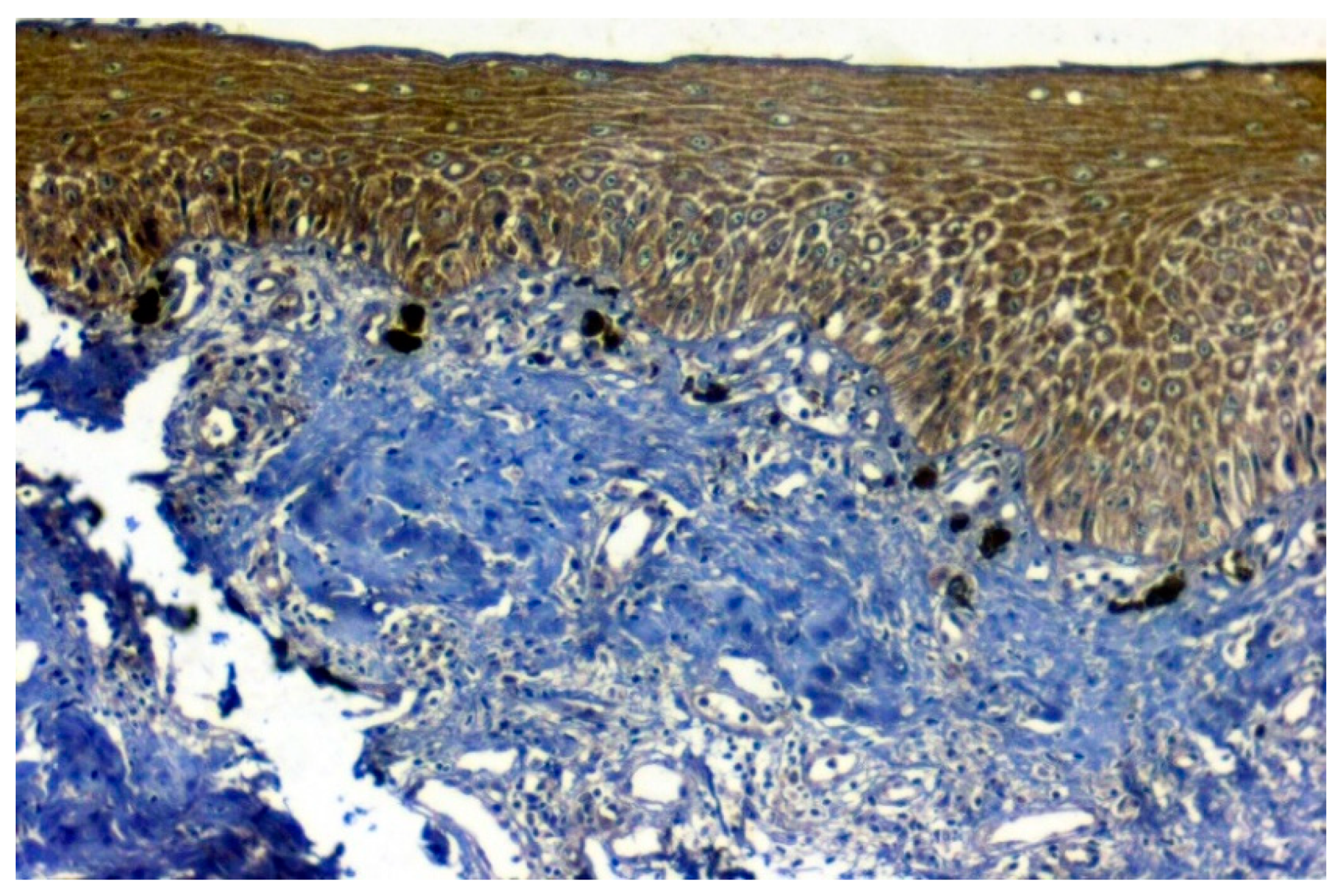
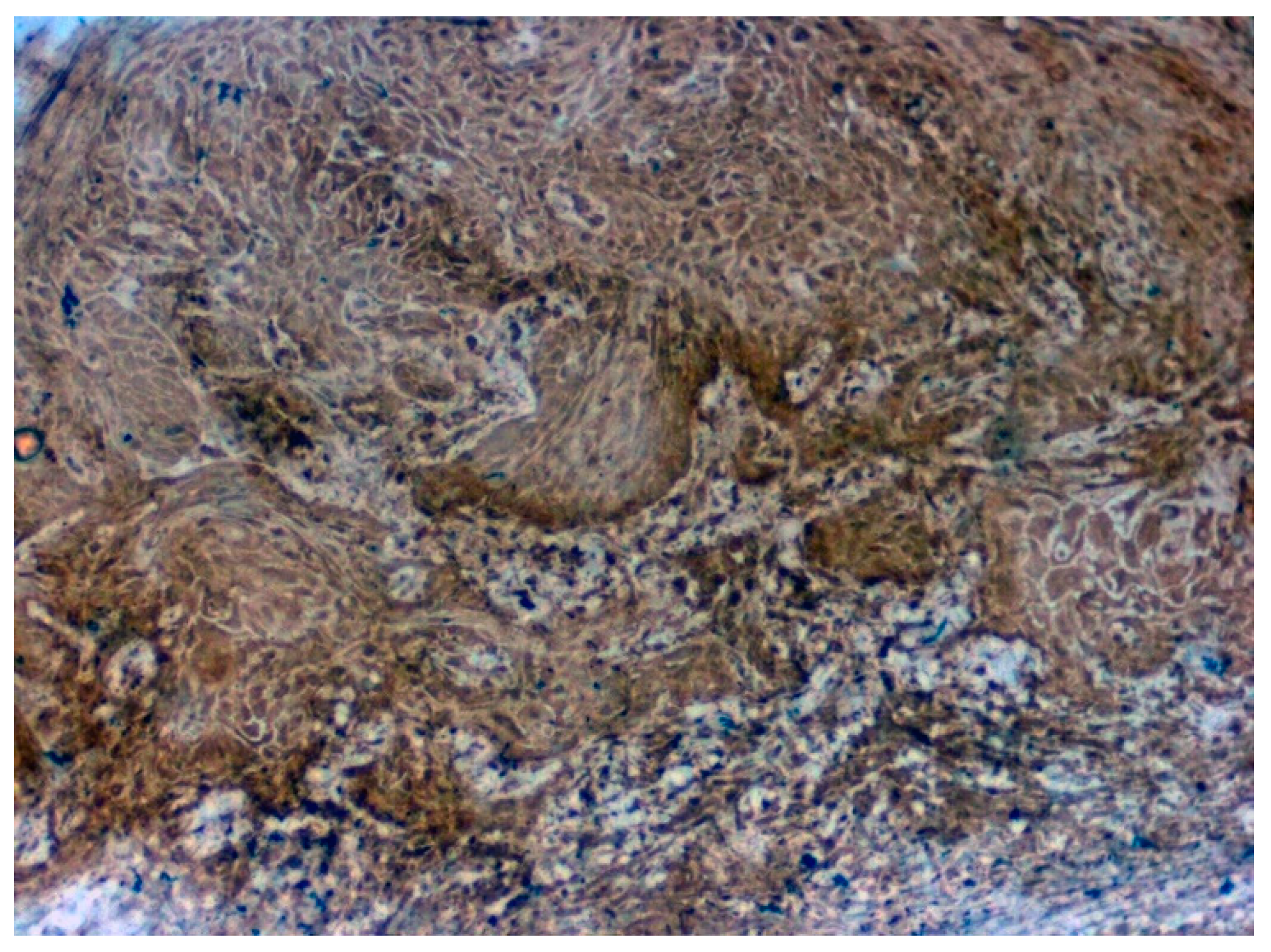
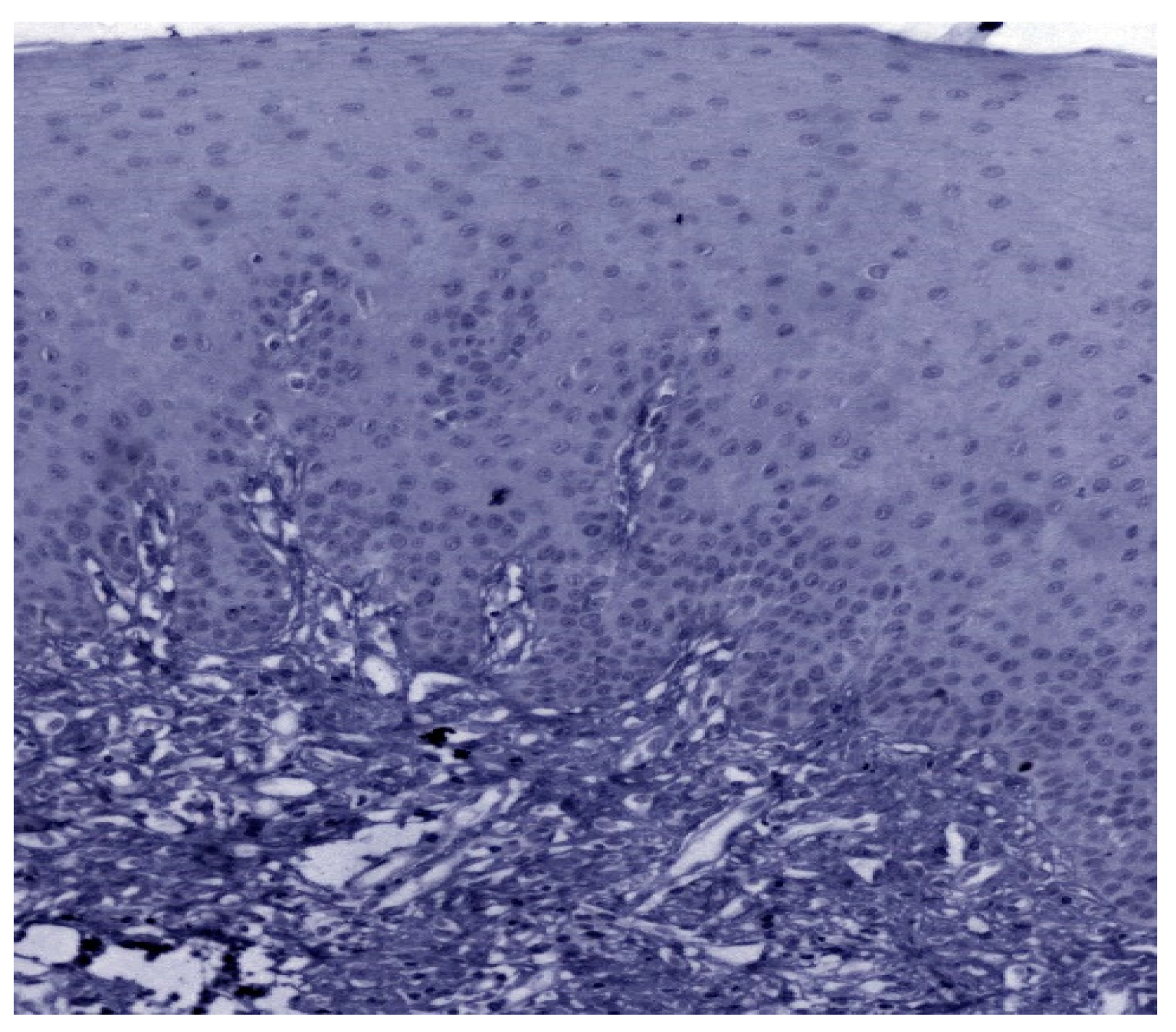
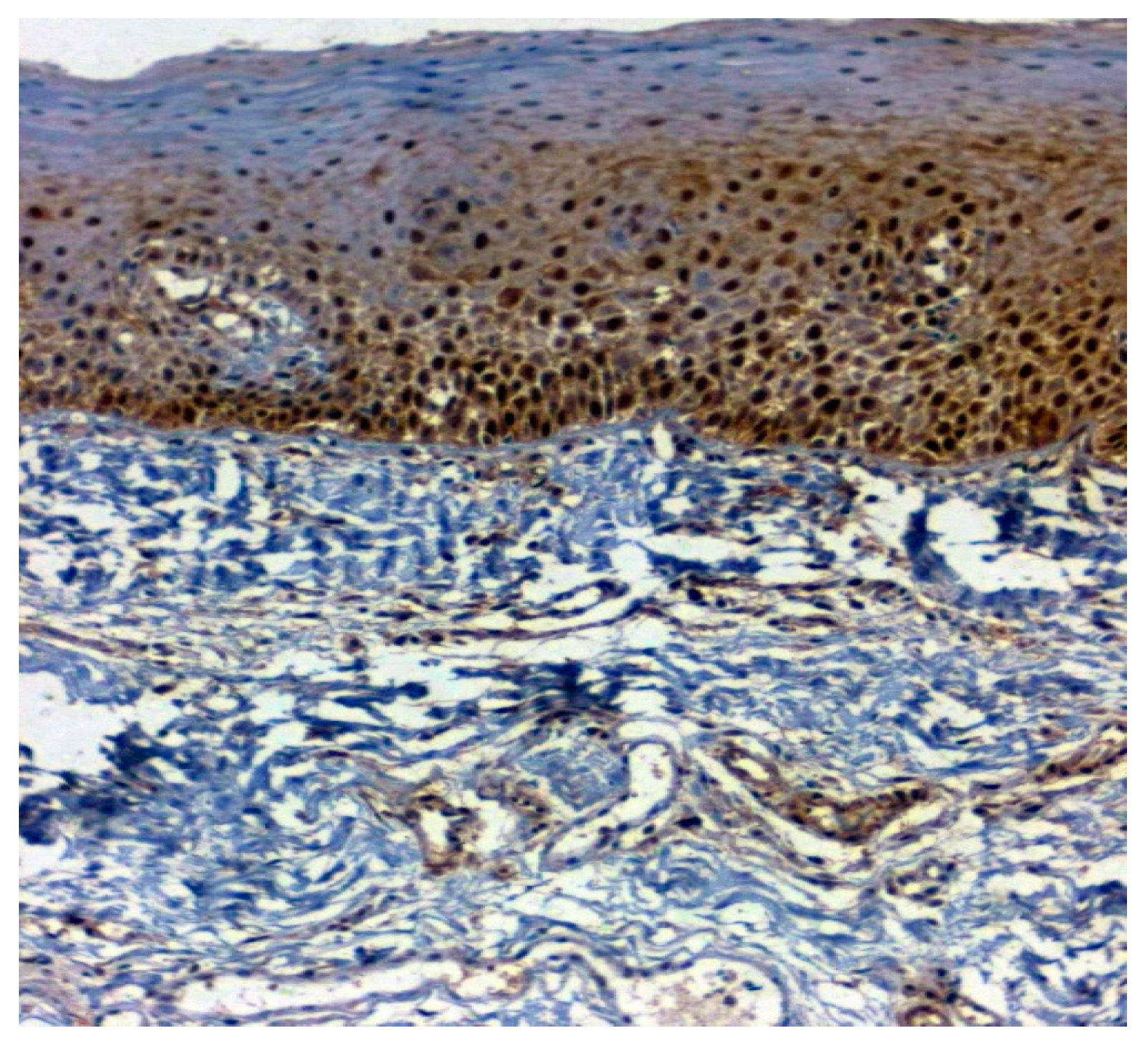
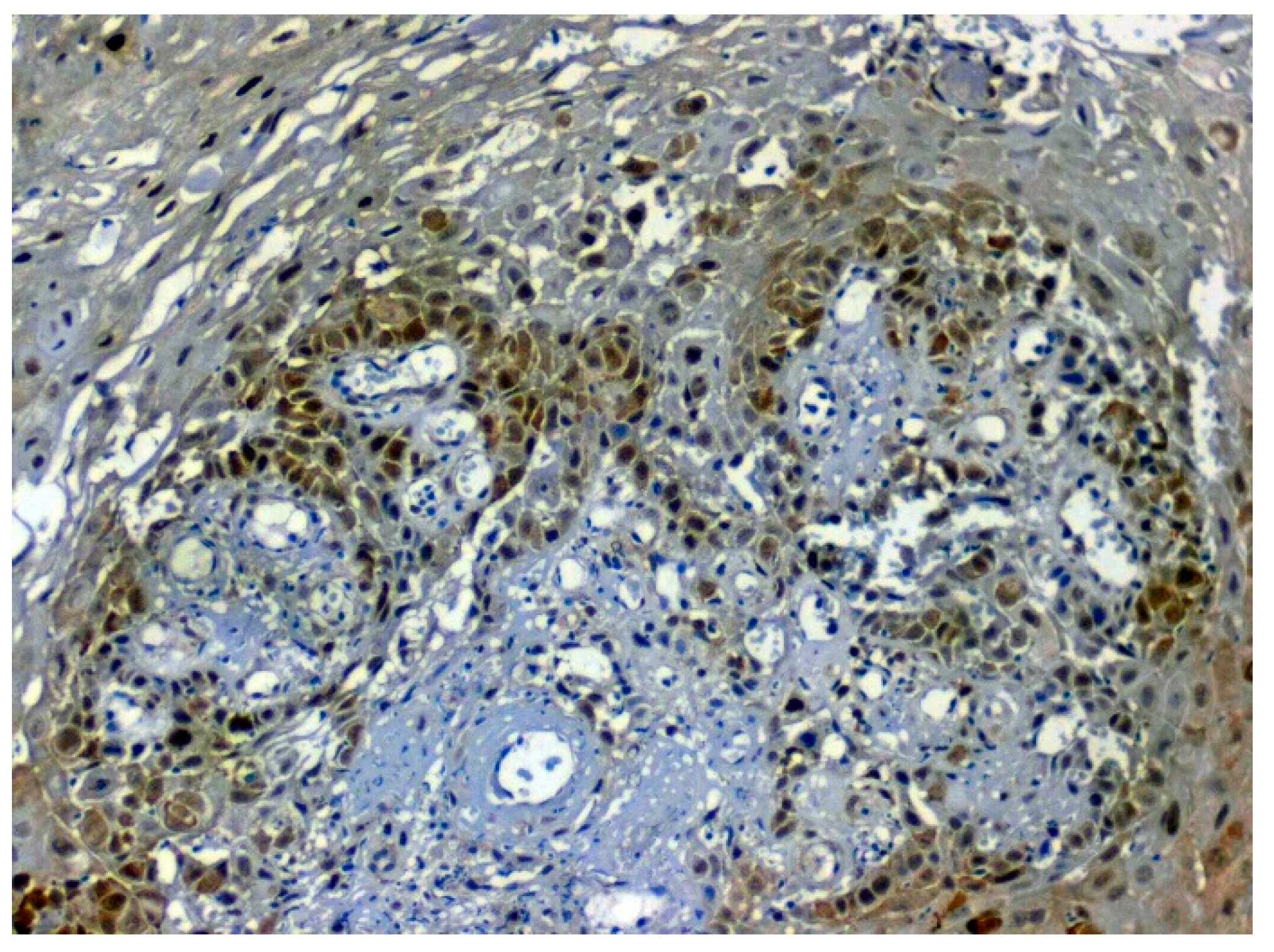
3.3. Validation Studies in Clinical Samples Using Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rajendran, R.; Sugathan, C.K.; Remani, P.; Ankathil, R.; Vijayakumar, T. Cell mediated and humoral immune responses in oral submucous fibrosis. Cancer 1986, 58, 2628–2631. [Google Scholar] [CrossRef]
- Angadi, P.V.; Rekha, K.P. Oral submucous fibrosis: A clinicopathologic review of 205 cases in Indians. Oral Maxillofac. Surg. 2010, 15, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.-M.; Kuo, T.-M.; Yeh, K.-T.; Lee, C.-H.; Ko, Y.-C. Reduction in and Preventive Effects for Oral-Cancer Risk with Antidepressant Treatment. J. Pers. Med. 2021, 11, 591. [Google Scholar] [CrossRef]
- Shih, Y.-H.; Wang, T.-H.; Shieh, T.-M.; Tseng, Y.-H. Oral Submucous Fibrosis: A Review on Etiopathogenesis, Diagnosis, and Therapy. Int. J. Mol. Sci. 2019, 20, 2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, U.; Bartsch, H.; Nair, J. Alert for an epidemic of oral cancer due to use of the betel quid substitutes gutkha and pan masala: A review of agents and causative mechanisms. Mutagenesis 2004, 19, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Yanjia, H.; Xinchun, J. The role of epithelial–mesenchymal transition in oral squamous cell carcinoma and oral submucous fibrosis. Clin. Chim. Acta 2007, 383, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zeng, L.; Li, N.; Wang, F.; Jiang, C.; Guo, F.; Chen, X.; Su, T.; Xu, C.; Zhang, S.; et al. Quantitative proteomic analysis for novel biomarkers of buccal squamous cell carcinoma arising in background of oral submucous fibrosis. BMC Cancer 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Hou, X.; Feng, H.; Liu, R.; Xu, H.; Gong, W.; Deng, J.; Sun, C.; Gao, Y.; Peng, J.; et al. Proteomic identification of cyclophilin A as a potential biomarker and therapeutic target in oral submucous fibrosis. Oncotarget 2016, 7, 60348–60365. [Google Scholar] [CrossRef]
- Das, T.; Prodhan, C.; Patsa, S.; Ray, J.G.; Chaudhuri, K. Identification of over expressed proteins in oral submucous fibrosis by proteomic analysis. J. Cell. Biochem. 2018, 119, 4361–4371. [Google Scholar] [CrossRef]
- Xie, X.; Jiang, Y.; Yuan, Y.; Wang, P.; Li, X.; Chen, F.; Sun, C.; Zhao, H.; Zeng, X.; Jiang, L.; et al. MALDI imaging reveals NCOA7 as a potential biomarker in oral squamous cell carcinoma arising from oral submucous fibrosis. Oncotarget 2016, 7, 59987–60004. [Google Scholar] [CrossRef]
- Chung, L.; Shibli, S.; Moore, K.; Elder, E.; Boyle, F.M.; Marsh, D.J.; Baxter, R.C. Tissue biomarkers of breast cancer and their association with conventional pathologic features. Br. J. Cancer 2013, 108, 351–360. [Google Scholar] [CrossRef]
- Gygi, S.P.; Corthals, G.; Zhang, Y.; Rochon, Y.; Aebersold, R. Evaluation of two-dimensional gel electrophoresis-based proteome analysis technology. Proc. Natl. Acad. Sci. USA 2000, 97, 9390–9395. [Google Scholar] [CrossRef] [Green Version]
- Jun, H.; Kieselbach, T.; Jönsson, L.J. Comparative proteome analysis of Saccharomyces cerevisiae: A global overview of in vivo targets of the yeast activator protein 1. BMC Genom. 2012, 13, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthi, S.; Prajna, N.V.; Lalitha, P.; Valarnila, M.; Dharmalingam, K. Pathogen Induced Changes in the Protein Profile of Human Tears from Fusarium Keratitis Patients. PLoS ONE 2013, 8, e53018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthi, S.; Chitra, T.; Bini, R.; Prajna, N.V.; Lalitha, P.; Dharmalingam, K. Comparative analysis of the tear protein profile in mycotic keratitis patients. Mol. Vis. 2008, 14, 500–507. [Google Scholar] [PubMed]
- Sivagnanam, A.; Shyamsundar, V.; Krishnamurthy, A.; Thangaraj, S.V.; Srinivas, C.V.D.; Kasirajan, H.; Ramani, P.; Ramshankar, V. Evaluation of Vimentin as a Potential Poor Prognostic Indicator and Salivary Biomarker for Oral Cancers and Pre-Cancers by Mass Spectrometry Based Proteomics. Res. Sq. 2021. Available online: https://assets.researchsquare.com/files/rs-656555/v1/f4c58576-2a8b-45e8-b8ac-f6acab40cf44.pdf?c=1631885615 (accessed on 21 December 2021).
- Zhu, H.; Fohlerová, Z.; Pekárek, J.; Basova, E.; Neužil, P. Recent advances in lab-on-a-chip technologies for viral diagnosis. Biosens. Bioelectron. 2020, 153, 112041. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Abdou, A.G.; Maraee, A.H.; Reyad, W. Immunohistochemical expression of heat shock protein 70 in vitiligo. Ann. Diagn. Pathol. 2013, 17, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Selves, J.; Long-Mira, E.; Mathieu, M.-C.; Rochaix, P.; Ilié, M. Immunohistochemistry for Diagnosis of Metastatic Carcinomas of Unknown Primary Site. Cancers 2018, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Hosur, M.B.; Puranik, R.S.; Vanaki, S.S.; Puranik, S.R.; Sudhakara, M.; Das, S. Evaluation of immunohistochemical expression of epithelial–mesenchymal transition markers E-cadherin, Twist and Snail in oral submucous fibrosis and their possible association with malignant transformation. J. Oral Maxillofac. Pathol. 2021, 25, 97–104. [Google Scholar] [CrossRef]
- Tavassol, F.; Starke, O.F.; Kokemüller, H.; Wegener, G.; Müller-Tavassol, C.C.; Gellrich, N.-C.; Eckardt, A. Prognostic significance of heat shock protein 70 (HSP70) in patients with oral cancer. Head Neck Oncol. 2011, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-W.; Kim, J.-Y.; Park, J.-Y.; Cha, I.-H.; Kim, J.; Lee, S. Expression of carbonic anhydrase IX is associated with postoperative recurrence and poor prognosis in surgically treated oral squamous cell carcinoma. Hum. Pathol. 2008, 39, 1317–1322. [Google Scholar] [CrossRef]
- Raungrut, P.; Petjaroen, P.; Geater, S.L.; Keeratichananont, W.; Phukaoloun, M.; Suwiwat, S.; Thongsuksai, P. Methylation of 14-3-3σ gene and prognostic significance of 14-3-3σ expression in non-small cell lung cancer. Oncol. Lett. 2017, 14, 5257–5264. [Google Scholar] [CrossRef] [Green Version]
- How to Cite IBM SPSS Statistics or Earlier Versions of SPSS. Available online: https://www.ibm.com/support/pages/how-cite-ibm-spss-statistics-or-earlier-versions-spss (accessed on 28 December 2021).
- Santosh, A.B.R.; Jones, T.; Harvey, J. A review on oral cancer biomarkers: Understanding the past and learning from the present. J. Cancer Res. Ther. 2016, 12, 486–492. [Google Scholar] [CrossRef]
- Pillai, J.; Chincholkar, T.; Dixit, R.; Pandey, M. A systematic review of proteomic biomarkers in oral squamous cell cancer. World J. Surg. Oncol. 2021, 19, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, P.C.; Liotta, L.A.; Iii, E.F.P. Cancer Proteomics: The State of the Art. Dis. Markers 2001, 17, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Katsani, K.R.; Sakellari, D. Saliva proteomics updates in biomedicine. J. Biol. Res. 2019, 26, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chandramouli, K.; Qian, P.-Y. Proteomics: Challenges, Techniques and Possibilities to Overcome Biological Sample Complexity. Hum. Genom. Proteom. 2009, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, J.O.R.; Oehler, M.K.; Ruszkiewicz, A.; McColl, S.R.; Hoffmann, P. MALDI Imaging Mass Spectrometry (MALDI-IMS)―Application of Spatial Proteomics for Ovarian Cancer Classification and Diagnosis. Int. J. Mol. Sci. 2011, 12, 773–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthi, S.; Lakshmi, C.N.P.; Atmika, P.; Anbarasu, K.; Mahalingam, S. Global Quantitative Proteomics reveal Deregulation of Cytoskeletal and Apoptotic Signalling Proteins in Oral Tongue Squamous Cell Carcinoma. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Van Heusden, G.P.H. 14-3-3 Proteins: Regulators of numerous eukaryotic proteins. IUBMB Life 2005, 57, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Pozuelo-Rubio, M. Proteomic and biochemical analysis of 14-3-3-binding proteins during C2-ceramide-induced apoptosis. FEBS J. 2010, 277, 3321–3342. [Google Scholar] [CrossRef]
- Rubio, M.P.; Geraghty, K.M.; Wong, B.H.C.; Wood, N.; Campbell, D.G.; Morrice, N.; Mackintosh, C. 14-3-3-affinity purification of over 200 human phosphoproteins reveals new links to regulation of cellular metabolism, proliferation and trafficking. Biochem. J. 2004, 379, 395–408. [Google Scholar] [CrossRef]
- Kaplan, A.; Ottmann, C.; Fournier, A.E. 14-3-3 adaptor protein-protein interactions as therapeutic targets for CNS diseases. Pharmacol. Res. 2017, 125, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Skoulakis, E.C.M.; Acevedo, S.F. 14-3-3s are potential biomarkers for HIV-related neurodegeneration. J. NeuroVirology 2012, 18, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef]
- Sluchanko, N.; Gusev, N.B. Moonlighting chaperone-like activity of the universal regulatory 14-3-3 proteins. FEBS J. 2017, 284, 1279–1295. [Google Scholar] [CrossRef] [Green Version]
- Diallo, K.; Oppong, A.K.; Lim, G.E. Can 14-3-3 proteins serve as therapeutic targets for the treatment of metabolic diseases? Pharmacol. Res. 2018, 139, 199–206. [Google Scholar] [CrossRef]
- Shimada, T.; Fournier, A.E.; Yamagata, K. Neuroprotective Function of 14-3-3 Proteins in Neurodegeneration. Biomed. Res. Int. 2013, 2013, 564534. [Google Scholar] [CrossRef]
- Hermeking, H. 14-3-3 proteins and cancer biology. Semin. Cancer Biol. 2006, 16, 161. [Google Scholar] [CrossRef] [PubMed]
- Mboge, M.Y.; Mahon, B.P.; McKenna, R.; Frost, S.C. Carbonic Anhydrases: Role in pH Control and Cancer. Metabolites 2018, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Occhipinti, R.; Boron, W.F. Role of Carbonic Anhydrases and Inhibitors in Acid–Base Physiology: Insights from Mathematical Modeling. Int. J. Mol. Sci. 2019, 20, 3841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alterio, V.; Kellner, M.; Esposito, D.; Liesche-Starnecker, F.; Bua, S.; Supuran, C.T.; Monti, S.M.; Zeidler, R.; De Simone, G. Biochemical and Structural Insights into Carbonic Anhydrase XII/Fab6A10 Complex. J. Mol. Biol. 2019, 431, 4910–4921. [Google Scholar] [CrossRef] [PubMed]
- Frost, S.C. Physiological Functions of the Alpha Class of Carbonic Anhydrases. Subcell. Biochem. 2013, 75, 9–30. [Google Scholar] [CrossRef]
- Hilvo, M.; Tolvanen, M.; Clark, A.; Shen, B.; Shah, G.N.; Waheed, A.; Halmi, P.; Hänninen, M.; Hämäläinen, J.M.; Vihinen, M.; et al. Characterization of CA XV, a new GPI-anchored form of carbonic anhydrase. Biochem. J. 2005, 392, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Li, R.-F.; Man, Q.-W.; Liu, J.-Y.; Zheng, Y.-Y.; Gao, X.; Liu, H.-M. Overexpression of T-type calcium channel Cav3.1 in oral squamous cell carcinoma: Association with proliferation and anti-apoptotic activity. Histochem. J. 2021, 1–10. [Google Scholar] [CrossRef]
- Chatterjee, S.; Burns, T.F. Targeting Heat Shock Proteins in Cancer: A Promising Therapeutic Approach. Int. J. Mol. Sci. 2017, 18, 1978. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [Green Version]
- Priyanka, K.P.; Majumdar, S.; Kotina, S.; Uppala, D.; Balla, H. Expression of Heat Shock Protein 70 in Oral Epithelial Dysplasia and Oral Squamous Cell Carcinoma: An Immunohistochemical Study. Contemp. Clin. Dent. 2020, 10, 185–190. [Google Scholar] [CrossRef]
- Thubashini, M.; Kannan, L.; Malathi, N.; M, T.; N, M.; L, K. Expression of heat shock protein70 in oral submucous fibrosis and oral squamous cell carcinoma: An immunohistochemical study. Indian J. Dent. Res. 2011, 22, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Carreras-Torras, C.; Escoda, C.G. Techniques for early diagnosis of oral squamous cell carcinoma: Systematic review. Med. Oral Patol. Oral Cir. Bucal 2015, 20, e305–e315. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.F.; Tseng, C.-H.; Lu, P.H.; Wang, Y.-P. Contemporary Molecular Analyses of Malignant Tumors for Precision Treatment and the Implication in Oral Squamous Cell Carcinoma. J. Pers. Med. 2022, 12, 12. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot ID | Gene ID | Accession | Description | Score | Coverage | Proteins | Unique Peptides | Peptides | PSMs | AAs | MW [kDa] | calc. pI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | MYL1 | P05976 | Myosin light chain 1/3, skeletal muscle isoform OS = Homo sapiens GN = MYL1 PE = 1 SV = 3-[MYL1_HUMAN] | 21.57 | 60.31 | 3 | 4 | 8 | 12 | 194 | 21.1 | 5.03 |
| 2 | HSPA8 | P11142 | Heat shock cognate 71 kDa protein OS = Homo sapiens GN = HSPA8 PE = 2 SV = 1-[P11142_HUMAN] | 34.78 | 19.94 | 43 | 6 | 11 | 16 | 627 | 68.8 | 5.52 |
| 3 | ENO3 | P13929 | Beta-enolase OS = Homo sapiens GN = ENO3 PE = 1 SV = 5-[ENOB_HUMAN] | 28.92 | 44.47 | 18 | 4 | 16 | 19 | 434 | 47.0 | 7.71 |
| 4 | ENO1 | P06733 | Alpha-enolase OS = Homo sapiens GN = ENO1 PE = 1 SV=2-[ENOA_HUMAN] | 51.42 | 49.08 | 24 | 7 | 17 | 36 | 434 | 47.1 | 7.39 |
| 5 | LUM | P51884 | Lumican OS = Homo sapiens GN = LUM PE = 1 SV = 2-[LUM_HUMAN] | 47.77 | 32.54 | 2 | 7 | 10 | 21 | 338 | 38.4 | 6.61 |
| 6 | CA2 | P00918 | Carbonic anhydrase 2 OS = Homo sapiens GN = CA2 PE = 1 SV = 2-[CAH2_HUMAN] | 121.94 | 55.38 | 3 | 12 | 12 | 33 | 260 | 29.2 | 7.40 |
| 7 | CA1 | P00915 | Carbonic anhydrase 1 OS = Homo sapiens GN = CA1 PE=1 SV =2-[CAH1_HUMAN] | 18.68 | 57.85 | 13 | 6 | 10 | 11 | 261 | 28.85 | 7.12 |
| 8 | GSTP1 | P09211 | Glutathione S-transferase P OS = Homo sapiens GN = GSTP1 PE = 1 SV = 2-[GSTP1_HUMAN] | 207.61 | 56.67 | 5 | 7 | 10 | 55 | 210 | 23.3 | 5.64 |
| 9 | HBB | P68871 | Hemoglobin beta (Fragment) OS = Homo sapiens GN = HBB PE = 2 SV = 1-[P68871_HUMAN] | 273.10 | 73.33 | 15 | 0 | 8 | 171 | 105 | 11.4 | 6.68 |
| 10 | KRT9 | P35527 | Keratin, type I cytoskeletal 9 OS = Homo sapiens GN = KRT9 PE = 1 SV = 3-[K1C9_HUMAN] | 48.25 | 49.92 | 2 | 10 | 21 | 28 | 623 | 62.0 | 5.24 |
| 11 | KRT3 | P12035 | Keratin, type II cytoskeletal 3 OS = Homo sapiens GN = KRT3 PE = 1 SV = 3-[K2C3_HUMAN] | 22.03 | 23.09 | 8 | 0 | 15 | 20 | 628 | 64.4 | 6.48 |
| 12 | KRT6C | P48668 | Keratin, type II cytoskeletal 6C OS = Homo sapiens GN = KRT6C PE = 1 SV = 3-[K2C6C_HUMAN] | 110.84 | 45.92 | 22 | 9 | 28 | 62 | 564 | 60.0 | 8.00 |
| 13 | HBA1 | P69905 | Hemoglobin subunit alpha OS = Homo sapiens GN = HBA1 PE = 1 SV = 2-[HBA_HUMAN] | 22.52 | 64.08 | 15 | 4 | 8 | 20 | 142 | 15.2 | 8.68 |
| 14 | ALB | A0A0C4DGB6 | Serum albumin OS = Homo sapiens PE = 2 SV = 1-A0A0C4DGB6_HUMAN] | 159.38 | 50.90 | 13 | 14 | 31 | 90 | 609 | 69.0 | 6.20 |
| 15 | ANXA2 | H0YMM1 | Annexin (Fragment) OS = Homo sapiens GN = ANXA2 PE = 2 SV = 1-[H0YMM1_HUMAN] | 22.31 | 26.85 | 24 | 3 | 3 | 7 | 149 | 16.4 | 5.91 |
| 16 | VIM | P08670 | Vimentin OS = Homo sapiens GN = VIM PE = 3 SV = 1-[P08670_HUMAN] | 19.27 | 20.88 | 32 | 5 | 10 | 19 | 431 | 49.6 | 5.25 |
| 17 | YWHAE | P62258 | 14-3-3 protein epsilon OS = Homo sapiens GN = YWHAE PE = 1 SV = 1-[1433E_HUMAN] | 55.66 | 37.25 | 14 | 5 | 6 | 16 | 255 | 29.2 | 4.74 |
| 18 | SERPINB4 | Q5K634 | SCCA2/SCCA1 fusion protein isoform 1 OS = Homo sapiens PE = 2 SV = 1-[Q5K634_HUMAN] | 270.44 | 78.72 | 3 | 0 | 42 | 91 | 390 | 44.6 | 6.39 |
| 19 | MYL6 | P60660 | Myosin light polypeptide 6 OS = Homo sapiens GN = MYL6 PE = 2 SV = 1-[P60660_HUMAN] | 84.22 | 56.55 | 20 | 7 | 7 | 26 | 145 | 16.3 | 4.65 |
| 20 | PFN1 | P07737 | Profilin-1 OS = Homo sapiens GN = PFN1 PE = 1 SV = 2-[PROF1_HUMAN] | 28.97 | 57.86 | 3 | 4 | 9 | 23 | 140 | 15.0 | 8.27 |
| 21 | HSP90AA1 | Q2VPJ6 | HSP90AA1 protein (Fragment) OS = Homo sapiens GN = HSP90AA1 PE = 2 SV = 1-[Q2VPJ6_HUMAN] | 49.70 | 34.53 | 30 | 4 | 17 | 22 | 585 | 68.3 | 5.19 |
| 22 | KRT77 | Q0IIN1 | Keratin 77 OS = Homo sapiens GN = KRT77 PE = 2 SV = 1-[Q0IIN1_HUMAN] | 23.03 | 11.42 | 4 | 1 | 6 | 8 | 578 | 61.8 | 5.85 |
| 23 | DCD | P81605 | Dermcidin OS = Homo sapiens GN = DCD PE = 1 SV = 2-[DCD_HUMAN] | 9.25 | 10.00 | 1 | 1 | 1 | 4 | 110 | 11.3 | 6.54 |
| Spot ID | Protein | Regulation in OSMF Sample |
|---|---|---|
| 1 | Myosin light chain 1 | UP |
| 2 | Heat shock 70 kDa protein | UP |
| 3 | Beta-enolase | UP |
| 4 | Alpha-enolase | UP |
| 5 | Lumican | UP |
| 6 | Carbonic anhydrase 2 | UP |
| 7 | Carbonic anhydrase 1 | UP |
| 8 | Glutathione S-transferase P | UP |
| 9 | Hemoglobin subunit beta | UP |
| 10 | Keratin, type I cytoskeletal 9 | UP |
| 11 | Keratin, type II cytoskeletal 3 | UP |
| 12 | Keratin, type II cytoskeletal 6C | UP |
| 13 | Hemoglobin subunit alpha | UP |
| 14 | Serum Albumin | UP |
| 15 | Annexin A2 | UP |
| 16 | Vimentin | UP |
| 17 | 14-3-3 protein epsilon | UP |
| 18 | SCCA2/SCCA1 fusion protein isoform 1 | UP |
| 19 | Myosin light polypeptide 6 | UP |
| 20 | Profilin-1 | UP |
| 21 | HSP90AA1 protein (Fragment) | UP |
| 22 | Keratin 77 | UP |
| 23 | Dermcidin | UP |
| Criteria | Total (n = 125) | CA 1-Negative (n = 37) | CA 1-Positive (n = 88) |
|---|---|---|---|
| Age | |||
| <42 yrs | 60 | 21 (35%) | 39 (65%) |
| >42 yrs | 65 | 16 (24.6%) | 49 (75.4%) |
| Gender | |||
| Male | 93 | 29 (31.2%) | 64 (68.8%) |
| Female | 32 | 8 (25%) | 24 (75%) |
| OSMF Clinical Stage (n = 72) | |||
| Stage I | 16 | 4 (25%) | 12 (75%) |
| Stage II | 28 | 10 (35.7%) | 18 (64.3%) |
| Stage III | 26 | 2 (7.7%) | 24 (92.3%) |
| Stage IV | 2 | 0 (0%) | 2 (100%) |
| Habits (OSMF n = 72) | |||
| Pan | 33 | 9 (27.3%) | 24 (72.7%) |
| Betel Nut | 20 | 5 (25%) | 15 (75%) |
| Maava | 13 | 2 (15.4%) | 11 (84.6%) |
| Gutka | 6 | 0 (0%) | 6 (100%) |
| Vascularity (OSMF n = 72) | |||
| Normal | 10 | 2 (20%) | 8 (80%) |
| Reduced | 40 | 11 (27.5%) | 29 (72.5%) |
| Increased | 20 | 3 (15%) | 17 (85%) |
| Enlarged and Increased | 2 | 0 (0%) | 2 (100%) |
| OSCC Histological Stage (n = 40) | |||
| Well-differentiated OSCC | 28 | 8 (28.6%) | 20 (71.4%) |
| Moderately differentiated OSCC | 10 | 2 (20%) | 8 (80%) |
| Poorly differentiated OSCC | 2 | 0 (0%) | 2 (100%) |
| OSCC Clinical Stage (n = 40) | |||
| Stage I | 8 | 2 (25%) | 6 (75%) |
| Stage II | 12 | 3 (25%) | 9 (75%) |
| Stage III | 6 | 1 (16.7%) | 5 (83.3%) |
| Stage IV | 14 | 4 (28.6%) | 10 (71.4%) |
| Inflammation | |||
| No | 6 | 3 (50%) | 3 (50%) |
| Mild | 32 | 11 (34.4%) | 21 (65.6%) |
| Moderate | 55 | 14 (25.5%) | 41 (74.5%) |
| Severe | 32 | 9 (28.1%) | 23 (71.9%) |
| Fibrosis (OSMF n = 72) | |||
| Mild | 18 | 5 (27.8%) | 13 (72.2%) |
| Moderate | 28 | 8 (28.6%) | 20 (71.4%) |
| Severe | 26 | 3 (11.5%) | 23 (88.5%) |
| Significant Factors | |||
| Diagnosis | |||
| Normal | 13 | 11 (84.6%) | 2 (15.4%) |
| OSMF | 72 | 16 (22.2%) | 56 (77.8%) |
| OSCC | 40 | 10 (25%) | 30 (75%) |
| p value = 0.000; χ2 = 21.169 | |||
| Epithelial Nature | |||
| Normal | 30 | 16 (53.3%) | 14 (46.7%) |
| Atrophic | 49 | 10 (20.4%) | 39 (79.6%) |
| Atrophic+mild dysplasia | 1 | 0 (0%) | 1 (100%) |
| Atrophic+moderate dysplasia | 5 | 1 (20%) | 4 (80%) |
| OSCC | 40 | 10 (25%) | 30 (75%) |
| p value = 0.025; χ2 = 11.144 | |||
| Criteria | Total (n = 130) | 14-3-3ε-Negative (n = 24) | 14-3-3ε-Positive (n = 106) |
|---|---|---|---|
| Gender | |||
| Female | 32 | 4 (12.5) | 28 (87.5) |
| Male | 98 | 20 (20.4) | 78 (79.6) |
| OSMF Clinical Stage (n = 77) | |||
| Stage I | 19 | 5 (26.3%) | 14 (73.7%) |
| Stage II | 29 | 4 (13.8%) | 25 (86.2%) |
| Stage III | 26 | 1 (3.8%) | 25 (96.2%) |
| Stage IV | 3 | 1 (33.3%) | 2 (66.7%) |
| Habits (OSMF n = 77) | |||
| Pan | 36 | 7 (19.4%) | 29 (80.6%) |
| Maava | 13 | 3 (23.1%) | 10 (76.9%) |
| Gutka | 6 | 0 (0%) | 6 (100%) |
| Betel nut | 22 | 1 (4.5% | 21 (95.5%) |
| Fibrosis (OSMF n = 77) | |||
| Mild | 20 | 3 (25%) | 15 (75%) |
| Moderate | 29 | 3 (10.3%) | 26 (89.7%) |
| Severe | 28 | 3 (10.7%) | 25 (89.3%) |
| Vascularity (OSMF n = 77) | |||
| Normal | 12 | 4 (33.3%) | 8 (66.6%) |
| Reduced | 43 | 6 (14%) | 37 (86%) |
| Increased | 20 | 1 (5%) | 19 (95%) |
| Enlarged and increased | 02 | 0 (0%) | 2 (100%) |
| OSCC Histological Stage (n = 40) | |||
| Well-differentiated OSCC | 28 | 2 (7.1%) | 26 (92.9%) |
| Moderately differentiated OSCC | 10 | 1 (10%) | 9 (90%) |
| Poorly differentiated OSCC | 2 | 0 (0%) | 2 (100%) |
| OSCC Clinical Stage (n = 40) | |||
| Stage I | 8 | 0 (0%) | 8 (100%) |
| Stage II | 12 | 0 (0%) | 12 (100%) |
| Stage III | 6 | 0 (0%) | 6 (100%) |
| Stage IV | 14 | 3 (21.4%) | 11 (78.6%) |
| Significant | |||
| Age | |||
| <43 yrs | 64 | 17 (26.6%) | 47 (73.4%) |
| >43 yrs | 66 | 7 (10.6%) | 59 (89.4%) |
| p value = 0.01; χ2 = 5.496 | |||
| Diagnosis | |||
| Normal | 13 | 10 (76.9%) | 3 (23.1%) |
| OSMF | 77 | 11 (14.3%) | 66 (85.7%) |
| OSCC | 40 | 3 (7.5%) | 37 (92.5%) |
| p value = 0.000; χ2 = 33.600 | |||
| Epithelial Nature | |||
| Normal | 30 | 12 (40%) | 18 (60%) |
| Atrophic + mild dysplasia | 1 | 0 (0%) | 1 (100%) |
| Atrophic+ moderate dysplasia | 5 | 1 (20%) | 4 (80%) |
| Atrophic | 54 | 8 (14.8%) | 46 (85.2%) |
| Malignant | 40 | 3 (7.5%) | 37 (92.5%) |
| p value = 0.010; χ2 = 13.149 | |||
| Inflammation | |||
| No | 6 | 1 (16.7%) | 5 (83.3%) |
| Mild | 32 | 11 (34.4%) | 21 (65.6%) |
| Moderate | 60 | 6 (10%) | 54 (90%) |
| Severe | 32 | 6 (18.8%) | 26 (81.2%) |
| p value = 0.041; χ2 = 8.252 | |||
| Criteria | Total (n = 117) | HSP 70-Negative (n = 36) | HSP 70-Positive (n = 81) |
|---|---|---|---|
| Gender | |||
| Male | 85 | 27 (31.8%) | 58 (68.2%) |
| Female | 32 | 9 (28.1%) | 23 (71.9%) |
| OSMF Clinical Stage (n = 47) | |||
| Stage I | 10 | 3 (30%) | 7 (70%) |
| Stage II | 18 | 9 (50%) | 9 (50%) |
| Stage III | 16 | 2 (12.5%) | 14 (87.5%) |
| Stage IV | 3 | 1 (33.3%) | 2 (66.7%) |
| Habits (OSMF n = 47) | |||
| Pan | 28 | 10 (35.7%) | 18 (64.3%) |
| Betel Nut | 7 | 2 (28.6%) | 5 (71.4%) |
| Maava | 10 | 2 (20%) | 8 (80%) |
| Gutka | 2 | 1 (50%) | 1 (50%) |
| Inflammation | |||
| Mild | 44 | 19 (43.2%) | 25 (56.8%) |
| Moderate | 49 | 12 (24.5%) | 37 (75.5%) |
| Severe | 24 | 5 (20.8%) | 19 (79.2%) |
| Fibrosis (OSMF n = 47) | |||
| Mild | 10 | 3 (30%) | 7 (70%) |
| Moderate | 18 | 5 (27.8%) | 13 (72.2%) |
| Severe | 19 | 7 (36.8%) | 12 (63.2%) |
| Vascularity (OSMF n = 47) | |||
| Normal | 7 | 3 (42.9%) | 4 (57.1%) |
| Reduced | 23 | 8 (34.8%) | 15 (65.2%) |
| Increased | 17 | 4 (23.5%) | 13 (76.5%) |
| OSCC Histological Stage (n = 53) | |||
| Well-differentiated OSCC | 25 | 6 (24%) | 19 (76%) |
| Moderately differentiated OSCC | 24 | 5 (20.8%) | 19 (79.2%) |
| Poorly differentiated OSCC | 4 | 0 (0%) | 4 (100%) |
| OSCC Clinical Stage (n = 53) | |||
| Stage I | 9 | 1 (11.1%) | 8 (88.9%) |
| Stage II | 17 | 1 (5.9%) | 16 (94.1%) |
| Stage III | 13 | 5 (38.5%) | 8 (61.5%) |
| Stage IV | 14 | 4 (28.6%) | 10 (71.4%) |
| Significant | |||
| Age | |||
| <47 yrs | 57 | 24 (42.1%) | 33 (57.9%) |
| >47 yrs | 60 | 12 (20%) | 48 (80%) |
| p value = 0.016; χ2 = 6.705 | |||
| Diagnosis | |||
| Normal | 17 | 10 (58.8%) | 7 (41.2%) |
| OSMF | 47 | 15 (31.9%) | 32 (68.1%) |
| OSCC | 53 | 11 (20.8%) | 42 (79.2%) |
| p value = 0.012; χ2 = 8.805 | |||
| Epithelial Nature | |||
| Normal | 9 | 5 (55.6%) | 4 (44.4%) |
| Hypertrophic | 16 | 8 (50%) | 8 (50%) |
| Atrophic | 36 | 12 (33.3%) | 24 (66.7%) |
| Dysplasia | 03 | 0 (0%) | 3 (100%) |
| OSCC | 53 | 11 (20.8%) | 42 (79.2%) |
| p value = 0.05; χ2 = 9.313 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venugopal, D.C.; Ravindran, S.; Shyamsundar, V.; Sankarapandian, S.; Krishnamurthy, A.; Sivagnanam, A.; Madhavan, Y.; Ramshankar, V. Integrated Proteomics Based on 2D Gel Electrophoresis and Mass Spectrometry with Validations: Identification of a Biomarker Compendium for Oral Submucous Fibrosis—An Indian Study. J. Pers. Med. 2022, 12, 208. https://doi.org/10.3390/jpm12020208
Venugopal DC, Ravindran S, Shyamsundar V, Sankarapandian S, Krishnamurthy A, Sivagnanam A, Madhavan Y, Ramshankar V. Integrated Proteomics Based on 2D Gel Electrophoresis and Mass Spectrometry with Validations: Identification of a Biomarker Compendium for Oral Submucous Fibrosis—An Indian Study. Journal of Personalized Medicine. 2022; 12(2):208. https://doi.org/10.3390/jpm12020208
Chicago/Turabian StyleVenugopal, Divyambika Catakapatri, Soundharya Ravindran, Vidyarani Shyamsundar, Sathasivasubramanian Sankarapandian, Arvind Krishnamurthy, Ananthi Sivagnanam, Yasasve Madhavan, and Vijayalakshmi Ramshankar. 2022. "Integrated Proteomics Based on 2D Gel Electrophoresis and Mass Spectrometry with Validations: Identification of a Biomarker Compendium for Oral Submucous Fibrosis—An Indian Study" Journal of Personalized Medicine 12, no. 2: 208. https://doi.org/10.3390/jpm12020208
APA StyleVenugopal, D. C., Ravindran, S., Shyamsundar, V., Sankarapandian, S., Krishnamurthy, A., Sivagnanam, A., Madhavan, Y., & Ramshankar, V. (2022). Integrated Proteomics Based on 2D Gel Electrophoresis and Mass Spectrometry with Validations: Identification of a Biomarker Compendium for Oral Submucous Fibrosis—An Indian Study. Journal of Personalized Medicine, 12(2), 208. https://doi.org/10.3390/jpm12020208