
Combinations of Low-Frequency Genetic Variants Might Predispose to Familial Pancreatic Cancer



 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
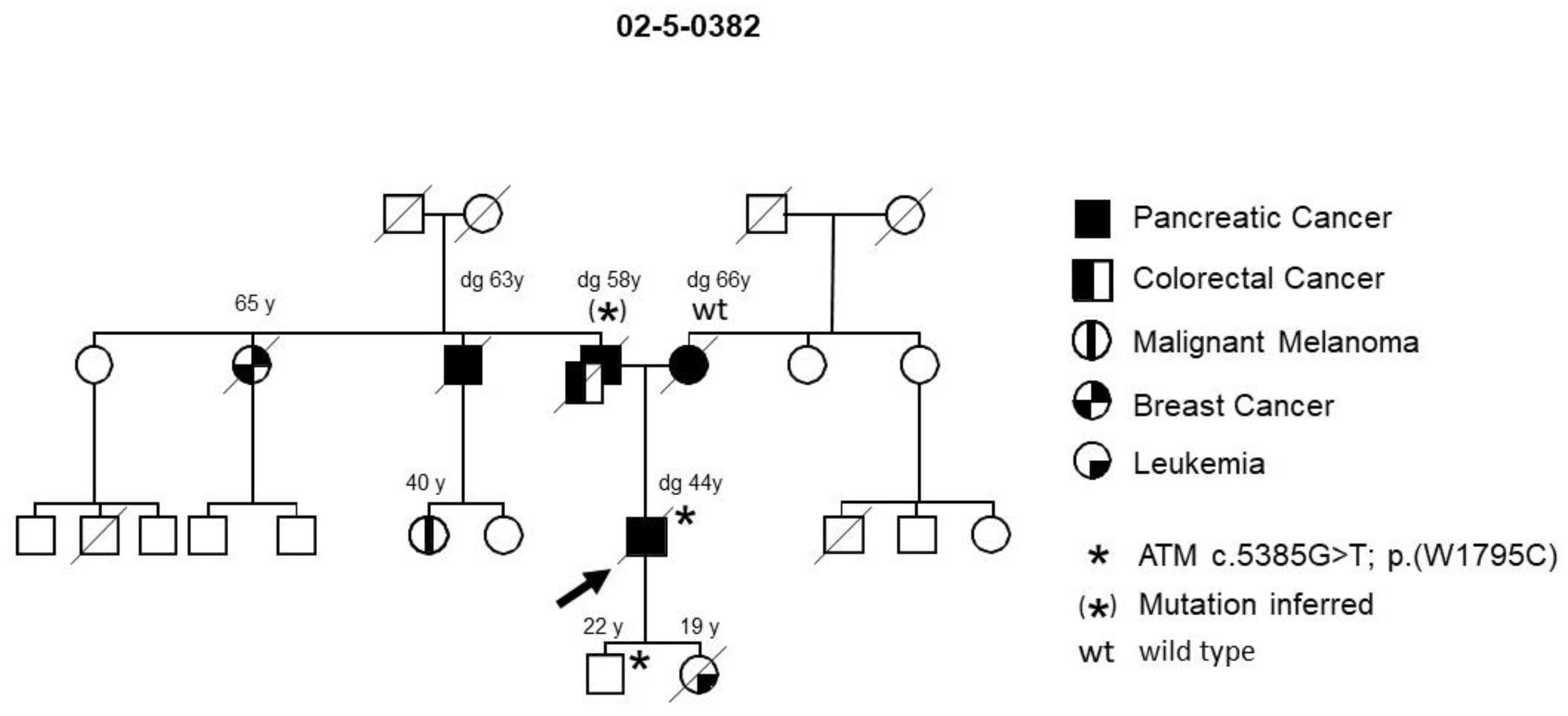
2.1. Recruitment of Cases and High Risk Individuals
2.2. DNA Extraction
2.3. Next Generation Sequencing of PDAC Cases, High Risk Individuals and Non-PDAC Cases
2.4. Immunohistochemical Staining
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quante, A.S.; Ming, C.; Rottmann, M.; Engel, J.; Boeck, S.; Heinemann, V.; Westphalen, C.B.; Strauch, K. Projections of cancer incidence and cancer-related deaths in Germany by 2020 and 2030. Cancer Med. 2016, 5, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, D.K.; Kress, R.; Sina-Frey, M.; Grützmann, R.; Gerdes, B.; Pilarsky, C.; Heise, J.W.; Schulte, K.-M.; Colombo-Benkmann, M.; Schleicher, C.; et al. Prevalence of familial pancreatic cancer in Germany. Int. J. Cancer 2004, 110, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, D.K.; Gress, T.; Langer, P. Familial pancreatic cancer—current knowledge. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Canto, M.I.; Harinck, F.; Hruban, R.H.; Offerhaus, G.J.; Poley, J.-W.; Kamel, I.; Nio, Y.; Schulick, R.S.; Bassi, C.; Kluijt, I.; et al. International Cancer of the Pancreas Screening (CAPS) Consortium summit on the management of patients with increased risk for familial pancreatic cancer. Gut 2012, 62, 339–347. [Google Scholar] [CrossRef]
- Bartsch, D.K.; Matthäi, E.; Mintziras, I.; Bauer, C.; Figiel, J.; Sina-Boemers, M.; Gress, T.M.; Langer, P.; Slater, E.P. The German National Case Collection for Familial Pancreatic Carcinoma (FaPaCa)—knowledge gained in 20 years. Dtsch. Aerzteblatt 2021, 18, 163–168. [Google Scholar]
- Fendrich, V.; Langer, P.; Bartsch, D.K. Familial pancreatic cancer—Status quo. Int. J. Colorectal Dis. 2014, 29, 139–145. [Google Scholar] [CrossRef]
- Abe, T.; Blackford, A.L.; Tamura, K.; Ford, M.; McCormick, P.; Chuidian, M.; Almario, J.A.; Borges, M.; Lennon, A.M.; Shin, E.J.; et al. Deleterious Germline Mutations Are a Risk Factor for Neoplastic Progression Among High-Risk Individuals Undergoing Pancreatic Surveillance. J. Clin. Oncol. 2019, 37, 1070–1080. [Google Scholar] [CrossRef]
- Vasen, H.F.A.; Ibrahim, I.S.; Ponce, C.G.; Slater, E.P.; Matthäi, E.; Carrato Mena, A.; Earl, J.; Robbers, K.; van Mil, A.M.; Potjer, T.P.; et al. The benefit of surveillance for pancreatic cancer in high-risk individuals: Outcome of long-term prospective follow-up studies from three European expert centres. J. Clin. Oncol. 2016, 34, 2010–2019. [Google Scholar] [CrossRef]
- Canto, M.I.; Almario, J.A.; Schulick, R.D.; Yeo, C.J.; Klein, A.; Blackford, A.; Shin, E.J.; Sanyal, A.; Yenokyan, G.; Lennon, A.M.; et al. Risk of Neoplastic Progression in Individuals at High Risk for Pancreatic Cancer Undergoing Long-term Surveillance. Gastroenterology 2018, 155, 740–751.e2. [Google Scholar] [CrossRef]
- Roberts, N.; Norris, A.L.; Petersen, G.M.; Bondy, M.L.; Brand, R.; Gallinger, S.; Kurtz, R.C.; Olson, S.H.; Rustgi, A.K.; Schwartz, A.G.; et al. Whole Genome Sequencing Defines the Genetic Heterogeneity of Familial Pancreatic Cancer. Cancer Discov. 2016, 6, 166–175. [Google Scholar] [CrossRef]
- Salo-Mullen, E.; O’Reilly, E.M.; Kelsen, D.P.; Ba, A.M.A.; Lowery, M.; Yu, K.; Reidy, D.L.; Epstein, A.; Lincoln, A.; Saldia, A.; et al. Identification of germline genetic mutations in patients with pancreatic cancer. Cancer 2015, 121, 4382–4388. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Yachida, S.; Shimizu, K.; Furuse, J.; Kubo, E.; Ohmoto, A.; Suzuki, M.; Hruban, R.H.; Okusaka, T.; Morizane, C.; et al. Germline mutations in Japanese familial pancreatic cancer patients. Oncotarget 2016, 7, 74227–74235. [Google Scholar] [CrossRef]
- Bartsch, D.K.; Sina-Frey, M.; Ziegler, A.; Hahn, S.A.; Przypadlo, E.; Kress, R.; Gerdes, B.; Rieder, H. Update of familial pan-creatic cancer in Germany. Pancreatology 2001, 1, 510–516. [Google Scholar] [CrossRef]
- Bartsch, D.K.; Sina-Frey, M.; Lang, S.; Wild, A.; Gerdes, B.; Barth, P.; Kress, R.; Grützmann, R.; Colombo-Benkmann, M.; Ziegler, A.; et al. CDKN2A Germline Mutations in Familial Pancreatic Cancer. Ann. Surg. 2002, 236, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.A.; Greenhalf, B.; Ellis, I.; Sina-Frey, M.; Rieder, H.; Korte, B.; Gerdes, B.; Kress, R.; Ziegler, A.; Raeburn, J.A.; et al. BRCA2 Germline Mutations in Familial Pancreatic Carcinoma. J. Natl. Cancer Inst. 2003, 95, 214–221. [Google Scholar] [CrossRef]
- Slater, E.P.; Langer, P.; Niemczyk, E.; Strauch, K.; Butler, J.; Habbe, N.; Neoptolemos, J.; Greenhalf, W.; Bartsch, D.K. PALB2 mutations in European familial pancreatic cancer families. Clin. Genet. 2010, 78, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Slater, E.P.; Langer, P.; Fendrich, V.; Habbe, N.; Chaloupka, B.; Matthäi, E.; Sina, M.; Hahn, S.A.; Bartsch, D.K. Prevalence of BRCA2 and CDKN2a mutations in German familial pancreatic cancer families. Fam. Cancer 2010, 9, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Jun, G.; Wing, M.K.; Abecasis, G.R.; Kang, H.M. An efficient and scalable analysis framework for variant extraction and refinement from population-scale DNA sequence data. Genome Res. 2015, 25, 918–925. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jian, X.; Boerwinkle, E. dbNSFP: A lightweight database of human nonsynonymous SNPs and their functional predic-tions. Hum. Mutat. 2011, 32, 894–899. [Google Scholar] [CrossRef]
- Liu, X.; Wu, C.; Li, C.; Boerwinkle, E. dbNSFP v3.0: A One-Stop Database of Functional Predictions and Annotations for Human Nonsynonymous and Splice-Site SNVs. Hum. Mutat. 2016, 37, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [PubMed]
- Khanna, K.K. Cancer Risk and the ATM Gene: A Continuing Debate. J. Natl. Cancer Inst. 2000, 92, 795–802. [Google Scholar] [CrossRef]
- Rahman, N. Realizing the promise of cancer predisposition genes. Nature 2014, 505, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Liu, W.; Mahdessian, H.; Bryant, P.; Ringdahl, J.; Timofeeva, M.; Farrington, S.M.; Dunlop, M.; Lindblom, A. Recurrent, low-frequency coding variants contributing to colorectal cancer in the Swedish population. PLoS ONE 2018, 13, e0193547. [Google Scholar] [CrossRef] [PubMed]
- Permuth, J.B.; Pirie, A.; Chen, Y.; Lin, H.-Y.; Reid, B.M.; Chen, Z.; Monteiro, A.; Dennis, J.; Mendoza-Fandino, G.; Anton-Culver, H.; et al. Exome genotyping arrays to identify rare and low frequency variants associated with epithelial ovarian cancer risk. Hum. Mol. Genet. 2016, 25, 3600–3612. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, D.K.; Matthäi, E.; Mintziras, I.; Böhm, L.B.; Gercke, N.; Bauer, C.; Figiel, J.; Slater, E.P. Characteristics of Pure Familial Pancreatic Cancer Families and Those with Additional Breast Cancer. Open Access J. Oncol. Med. 2020, 4. [Google Scholar] [CrossRef]
- Couch, F.J.; Johnson, M.R.; Rabe, K.G.; Brune, K.; De Andrade, M.; Goggins, M.; Rothenmund, H.; Gallinger, S.; Klein, A.; Petersen, G.M.; et al. The Prevalence of BRCA2 Mutations in Familial Pancreatic Cancer. Cancer Epidemiol. Biomark. Prev. 2007, 16, 342–346. [Google Scholar] [CrossRef]
- Goldstein, A.; Fraser, M.; Struewing, J.; Hussussian, C.; Ranade, K.; Zametkin, D.; Fontaine, L.; Organic, S.; Dracopoli, N.; Clark, W.J.; et al. Increased risk of pancreatic cancer in melanoma-prone kindreds with p16INK4 mutations. N. Engl. J. Med. 1995, 333, 970–974. [Google Scholar] [CrossRef]
- Roberts, N.; Jiao, Y.; Yu, J.; Kopelovich, L.; Petersen, G.M.; Bondy, M.L.; Gallinger, S.; Schwartz, A.G.; Syngal, S.; Cote, M.L.; et al. ATM Mutations in Patients with Hereditary Pancreatic Cancer. Cancer Discov. 2011, 2, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Nakamura, H.; Chiku, S.; Kubo, E.; Ohmoto, A.; Totoki, Y.; Shibata, T.; Higuchi, R.; Yamamoto, M.; Furuse, J.; et al. Whole-exome Sequencing Reveals New Potential Susceptibility Genes for Japanese Familial Pancreatic Cancer. Ann. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.C.; Al-Sukhni, W.; Borgida, A.E.; Holter, S.; Kanji, Z.S.; McPherson, T.; Whelan, E.; Serra, S.; Trinh, Q.M.; Peltekova, V.; et al. Exome sequencing identifies nonsegregating nonsense ATM and PALB2 variants in familial pancreatic cancer. Hum. Genom. 2013, 7, 11. [Google Scholar] [CrossRef]
- Lavin, M.F. Ataxia-telangiectasia: From a rare disorder to a paradigm for cell signalling and cancer. Nat. Rev. Mol. Cell Biol. 2008, 9, 759–769. [Google Scholar] [CrossRef]
- Roger, E.; Gout, J.; Arnold, F.; Beutel, A.K.; Müller, M.; Abaei, A.; Barth, T.F.E.; Rasche, V.; Seufferlein, T.; Perkhofer, L.; et al. Maintenance Therapy for ATM-Deficient Pancreatic Cancer by Multiple DNA Damage Response Interferences after Platinum-Based Chemotherapy. Cells 2020, 9, 2110. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, Y.; Sun, B.; McMahon, A.P.; Wang, Y. Hedgehog Signaling: From Basic Biology to Cancer Therapy. Cell Chem. Biol. 2017, 24, 252–280. [Google Scholar] [CrossRef] [PubMed]
- Rhim, A.D.; Oberstein, P.E.; Thomas, D.H.; Mirek, E.T.; Palermo, C.F.; Sastra, S.A.; Dekleva, E.N.; Saunders, T.; Becerra, C.P.; Tattersall, I.; et al. Stromal Elements Act to Restrain, Rather Than Support, Pancreatic Ductal Adenocarcinoma. Cancer Cell 2014, 25, 735–747. [Google Scholar] [CrossRef]
- Pastorino, L.; Ghiorzo, P.; Nasti, S.; Battistuzzi, L.; Cusano, R.; Marzocchi, C.; Garrè, M.L.; Clementi, M.; Scarrà, G.B. Iden-tification of a SUFU germline mutation in a family with Gorlin syndrome. Am. J. Med. Genet. Part A 2009, 149, 1539–1543. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, D.K.; Slater, E.P.; Carrato, A.; Ibrahim, I.S.; Ponce, C.G.; Vasen, H.; Matthäi, E.; Earl, J.; Jendryschek, F.S.; Figiel, J.; et al. Refinement of screening for familial pancreatic cancer. Gut 2016, 65, 1314–1321. [Google Scholar] [CrossRef]
- Skaro, M.; Nanda, N.; Gauthier, C.; Felsenstein, M.; Jiang, Z.; Qiu, M.; Shindo, K.; Yu, J.; Hutchings, D.; Javed, A.A.; et al. Prevalence of Germline Mutations Associated with Cancer Risk in Patients with Intraductal Papillary Mucinous Neoplasms. Gastroenterology 2019, 156, 1905–1913. [Google Scholar] [CrossRef]
- Waddell, N.; Initiative, A.P.C.G.; Pajic, M.; Patch, A.-M.; Chang, D.; Kassahn, K.; Bailey, P.; Johns, A.L.; Miller, D.; Nones, K.; et al. Whole genomes redefine the mutational landscape of pancreatic cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef]
- Downs, B.; Sherman, S.; Cui, J.; Kim, Y.C.; Snyder, C.; Christensen, M.; Luo, J.; Lynch, H.; Wang, S.M. Common genetic variants contribute to incomplete penetrance: Evidence from cancer-free BRCA1 mutation carriers. Eur. J. Cancer 2019, 107, 68–78. [Google Scholar] [CrossRef]
- Nguyen-Dumont, T.; Myszka, A.; Karpinski, P.; Sasiadek, M.M.; Akopyan, H.; Hammet, F.; Tsimiklis, H.; Park, D.J.; Pope, B.J.; Slezak, R.; et al. FANCM and RECQL genetic variants and breast cancer susceptibility: Relevance to South Poland and West Ukraine. BMC Med Genet. 2018, 19, 12. [Google Scholar] [CrossRef]
- Sanada, K.; Gupta, A.; Tsai, L.H. Disabled-1-regulated adhesion of migrating neurons to radial glial fiber contributes to neu-ronal positioning during early corticogenesis. Neuron 2004, 42, 197–211. [Google Scholar] [CrossRef]
- Sato, N.; Fukushima, N.; Chang, R.; Matsubayashi, H.; Goggins, M. Differential and Epigenetic Gene Expression Profiling Identifies Frequent Disruption of the RELN Pathway in Pancreatic Cancers. Gastroenterology 2006, 130, 548–565. [Google Scholar] [CrossRef] [PubMed]
- Brandalize, A.P.C.; Schuler-Faccini, L.; Hoffmann, J.-S.; Caleffi, M.; Cazaux, C.; Ashton-Prolla, P. A DNA repair variant in POLQ (c.-1060A > G) is associated to hereditary breast cancer patients: A case-control study. BMC Cancer 2014, 14, 850. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Szabo, C.; Qian, C.; Amadio, P.G.; Thibodeau, S.N.; Cerhan, J.R.; Petersen, G.M.; Liu, W.; Couch, F.J. Mutational Analysis of Thirty-two Double-Strand DNA Break Repair Genes in Breast and Pancreatic Cancers. Cancer Res. 2008, 68, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Felicio, P.S.; Grasel, R.S.; Campacci, N.; de Paula, A.E.; Galvão, H.C.R.; Torrezan, G.T.; Sabato, C.S.; Fernandes, G.C.; Souza, C.P.; Michelli, R.D.; et al. Whole-exome sequencing of non-BRCA1/BRCA2 mutation carrier cases at high-risk for hereditary breast/ovarian cancer. Hum. Mutat. 2021, 42, 290–299. [Google Scholar] [CrossRef]
- Family, L.; Bensen, J.T.; Troester, M.A.; Wu, M.C.; Anders, C.K.; Olshan, A.F. Single-nucleotide polymorphisms in DNA bypass polymerase genes and association with breast cancer and breast cancer subtypes among African Americans and Whites. Breast Cancer Res. Treat. 2014, 149, 181–190. [Google Scholar] [CrossRef][Green Version]
- Kang, X.; Lin, Z.; Xu, M.; Pan, J.; Wang, Z. Deciphering role of FGFR signalling pathway in pancreatic cancer. Cell Prolif. 2019, 52, e12605. [Google Scholar] [CrossRef]
- Tassi, E.; Wellstein, A. The Angiogenic Switch Molecule, Secreted FGF-Binding Protein, an Indicator of Early Stages of Pancreatic and Colorectal Adenocarcinoma. In Seminars in Oncology; WB Saunders: Philadelphia, PA, USA, 2006; pp. 50–56. [Google Scholar]
- Jia, W.; Dong, Y.; Tao, L.; Pang, L.; Ren, Y.; Liang, W.; Jiang, J.; Cheng, G.; Zhang, W.J.; Yuan, X.; et al. MAP3K3 overexpression is associated with poor survival in ovarian carcinoma. Hum. Pathol. 2016, 50, 162–169. [Google Scholar] [CrossRef]
- Fan, Y.; Ge, N.; Wang, X.; Sun, W.; Mao, R.; Bu, W.; Creighton, C.J.; Zheng, P.; Vasudevan, S.; An, L.; et al. Amplification and over-expression ofMAP3K3gene in human breast cancer promotes formation and survival of breast cancer cells. J. Pathol. 2014, 232, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Sharma, R.; Saraya, A.; Chattopadhyay, T.K.; Dattagupta, S.; Walfish, P.G.; Chauhan, S.S.; Ralhan, R. Mitogen activated protein kinase kinase kinase 3 (MAP3K3/MEKK3) overexpression is an early event in esophageal tumorigenesis and is a predictor of poor disease prognosis. BMC Cancer 2014, 14, 2. [Google Scholar] [CrossRef]
- Slattery, M.L.; Lundgreen, A.; Wolff, R.K. MAP kinase genes and colon and rectal cancer. Carcinogenesis 2012, 33, 2398–2408. [Google Scholar] [CrossRef]
- Haack, T.B.; Danhauser, K.; Haberberger, B.; Hoser, J.; Strecker, V.; Boehm, D.; Uziel, G.; Lamantea, E.; Invernizzi, F.; Poulton, J.; et al. Exome sequencing identifies ACAD9 mutations as a cause of complex I deficiency. Nat. Genet. 2010, 42, 1131–1134. [Google Scholar] [CrossRef]
- Collet, M.; Assouline, Z.; Bonnet, D.; Rio, M.; Iserin, F.; Sidi, D.; Goldenberg, A.; Lardennois, C.; Metodiev, M.D.; Haberberger, B.; et al. High incidence and variable clinical outcome of cardiac hypertrophy due to ACAD9 mutations in childhood. Eur. J. Hum. Genet. 2015, 24, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Gato, D.; Thysell, E.; Tyanova, S.; Crnalic, S.; Santos, A.; Lima, T.S.; Geiger, T.; Cox, J.; Widmark, A.; Bergh, A.; et al. The Proteome of Prostate Cancer Bone Metastasis Reveals Heterogeneity with Prognostic Implications. Clin. Cancer Res. 2018, 24, 5433–5444. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chai, T.; Zhang, Y.; Chu, A.; Liang, B.; Guo, X.; Guo, Z.; Song, R.; Hou, G.; Yuan, J.; et al. Identification of abnormal nuclear and mitochondrial genes in esophageal cancer cells. Mol. Med. Rep. 2017, 17, 2817–2826. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Family ID | Family Type | Variant | Mutation Type | Biological Significance (Pathogenic, VUS *) | Segregation with Disease | Families with the Variant | MAF | Somatic Gene Variant Described in PDAC Tumor Tissue |
|---|---|---|---|---|---|---|---|---|---|
| ATM | 02-5-0382 | FPC | c.5385G > T | missense | deleterious | yes | 1/36 | n.a. | no |
| p.(W1795C) | |||||||||
| SUFU | 25-5-67 | FPC-breast | Position | promoter variant | pathogenic | yes | 1/15 | 0.000024 1 | no |
| −8C > T | |||||||||
| DAB1 # | 25-9-44 | PCMS | c.786 + 1G > A | splice | pathogenic | yes | 1/49 | 0.00001 1 | yes [7] |
| site | |||||||||
| POLQ # | 25-9-44 | PCMS | c.4141C > A p.(P1381T) | missense | VUS | yes | 1/29 | 0.015/ | no |
| 0.025 2 | |||||||||
| FGFBP3 # | 25-9-44 | PCMS | c.724G > A p.(A242T) | missense | deleterious | yes | 1/29 | 0.000004 1 | no |
| MAP3K3 § | 25-4-46 | FPC | c.1511T > C p.(I504T) | missense | deleterious | yes | 1/29 | 0.00001 1 | no |
| ACAD9 § | 25-4-46; 25-1-91; 25-9-113; 25-2-209 | FPC FPC FPC FPC | c.976G > A p.(A326T) | missense | VUS | yes | 4/46 | 0.0152/ 0.0219 2 | no |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slater, E.P.; Wilke, L.M.; Böhm, L.B.; Strauch, K.; Lutz, M.; Gercke, N.; Matthäi, E.; Hemminki, K.; Försti, A.; Schlesner, M.; et al. Combinations of Low-Frequency Genetic Variants Might Predispose to Familial Pancreatic Cancer. J. Pers. Med. 2021, 11, 631. https://doi.org/10.3390/jpm11070631
Slater EP, Wilke LM, Böhm LB, Strauch K, Lutz M, Gercke N, Matthäi E, Hemminki K, Försti A, Schlesner M, et al. Combinations of Low-Frequency Genetic Variants Might Predispose to Familial Pancreatic Cancer. Journal of Personalized Medicine. 2021; 11(7):631. https://doi.org/10.3390/jpm11070631
Chicago/Turabian StyleSlater, Emily P., Lisa M. Wilke, Lutz Benedikt Böhm, Konstantin Strauch, Manuel Lutz, Norman Gercke, Elvira Matthäi, Kari Hemminki, Asta Försti, Matthias Schlesner, and et al. 2021. "Combinations of Low-Frequency Genetic Variants Might Predispose to Familial Pancreatic Cancer" Journal of Personalized Medicine 11, no. 7: 631. https://doi.org/10.3390/jpm11070631
APA StyleSlater, E. P., Wilke, L. M., Böhm, L. B., Strauch, K., Lutz, M., Gercke, N., Matthäi, E., Hemminki, K., Försti, A., Schlesner, M., Paramasivam, N., & Bartsch, D. K. (2021). Combinations of Low-Frequency Genetic Variants Might Predispose to Familial Pancreatic Cancer. Journal of Personalized Medicine, 11(7), 631. https://doi.org/10.3390/jpm11070631