
Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Production and Microinjection of AAV Vector
2.4. Quantitative Real-Time PCR
2.5. Western Blotting
2.6. Electrophysiological Recordings
2.7. In Vivo Microdialysis
2.8. Optogenetic Stimulation
2.9. Behavioral Analyses
- Locomotor activity test
- PPI test
- Novel object recognition test
- Contextual learning test
- Y-maze test (spontaneous alternation behaviors)
- Mild social defeat stress exposure
- Social interaction test
- Forced swimming test
2.10. Statistical Analyses
3. Results
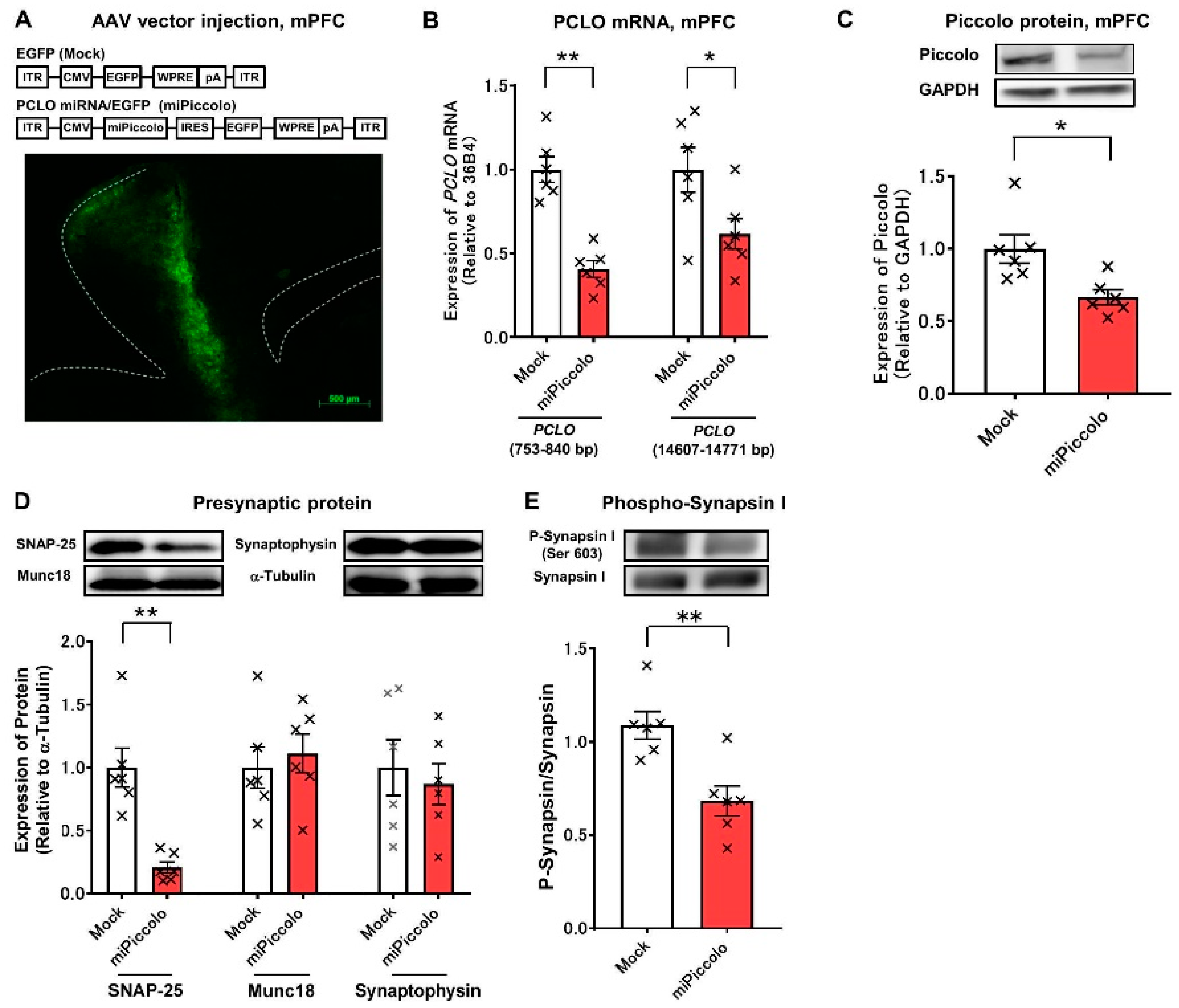
3.1. Mice with a Suppressed Piccolo Expression in the mPFC
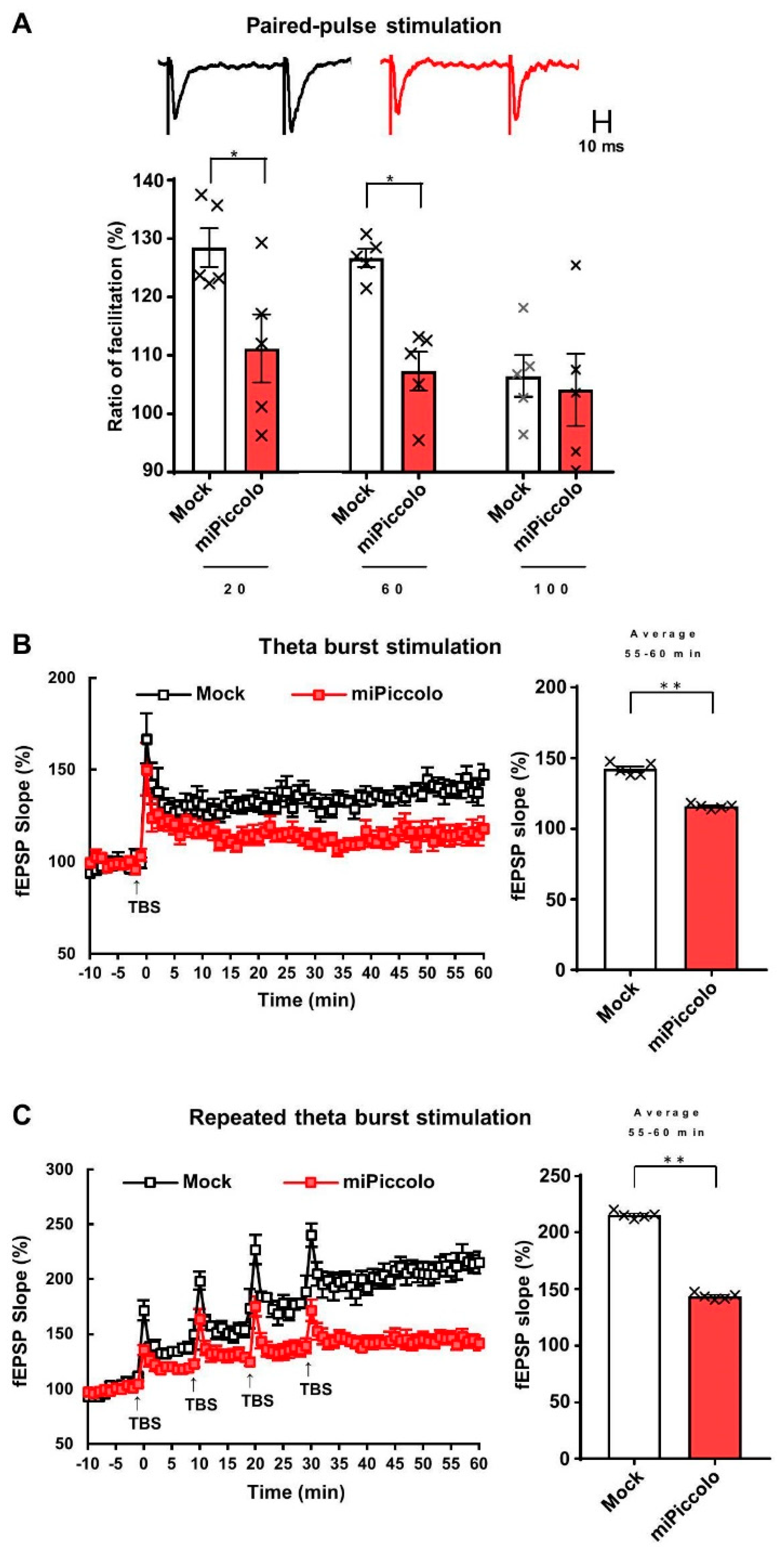
3.2. Diminished Synaptic Properties in the mPFC by Piccolo Suppression
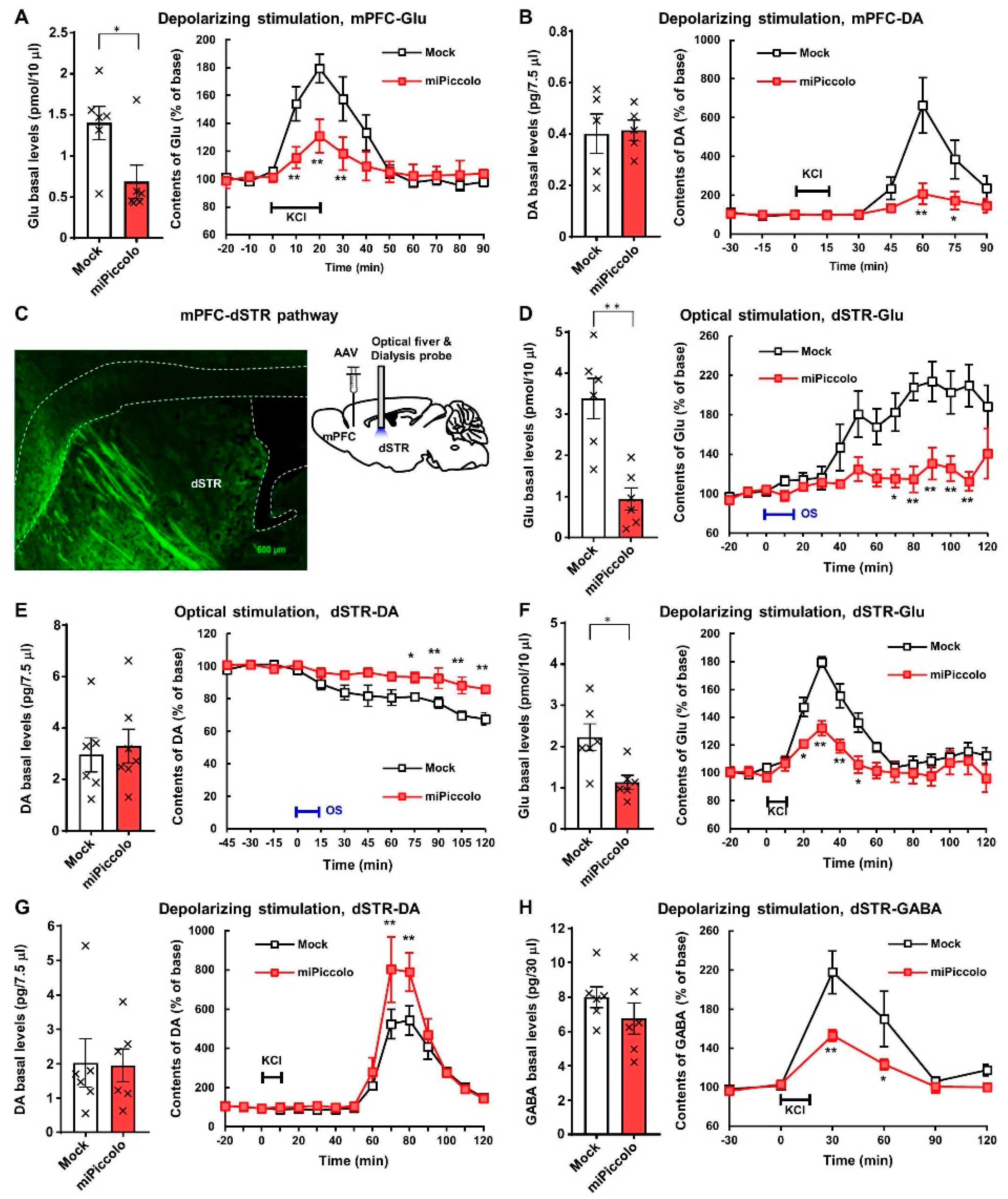
3.3. Reduction in Depolarization-Evoked Glutamate and Dopamine Elevation in the mPFC by Piccolo Suppression
3.4. Diminished Neuronal Responses in the mPFC-dSTR Pathway by Piccolo Suppression
3.5. Enhanced Depolarization-Evoked Dopamine Elevation Via Disinhibition of GABAergic Regulation in the dSTR by Piccolo Suppression
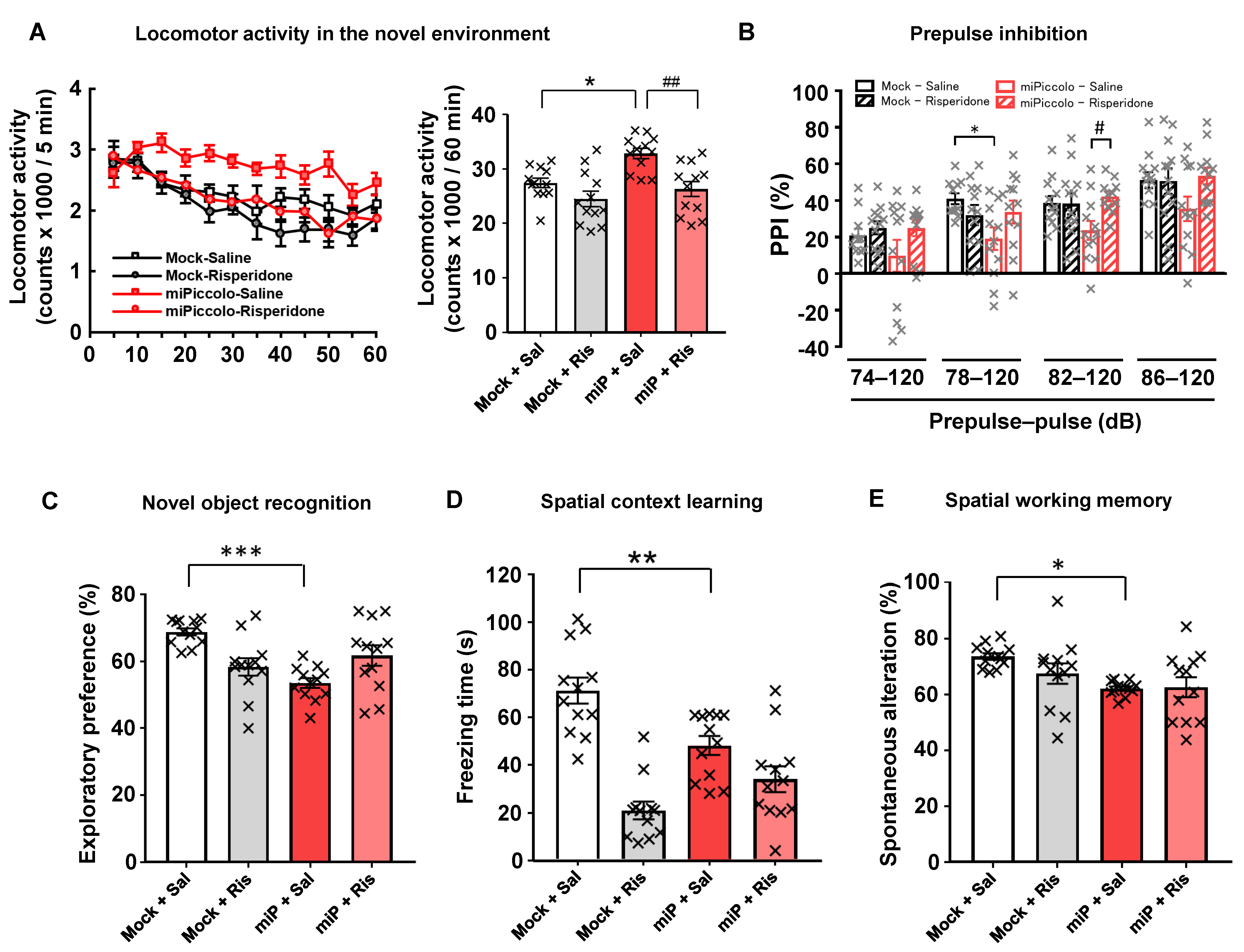
3.6. Disruption of Locomotor Activity, Sensorimotor Gating, and the Cognitive Function by Piccolo Suppression
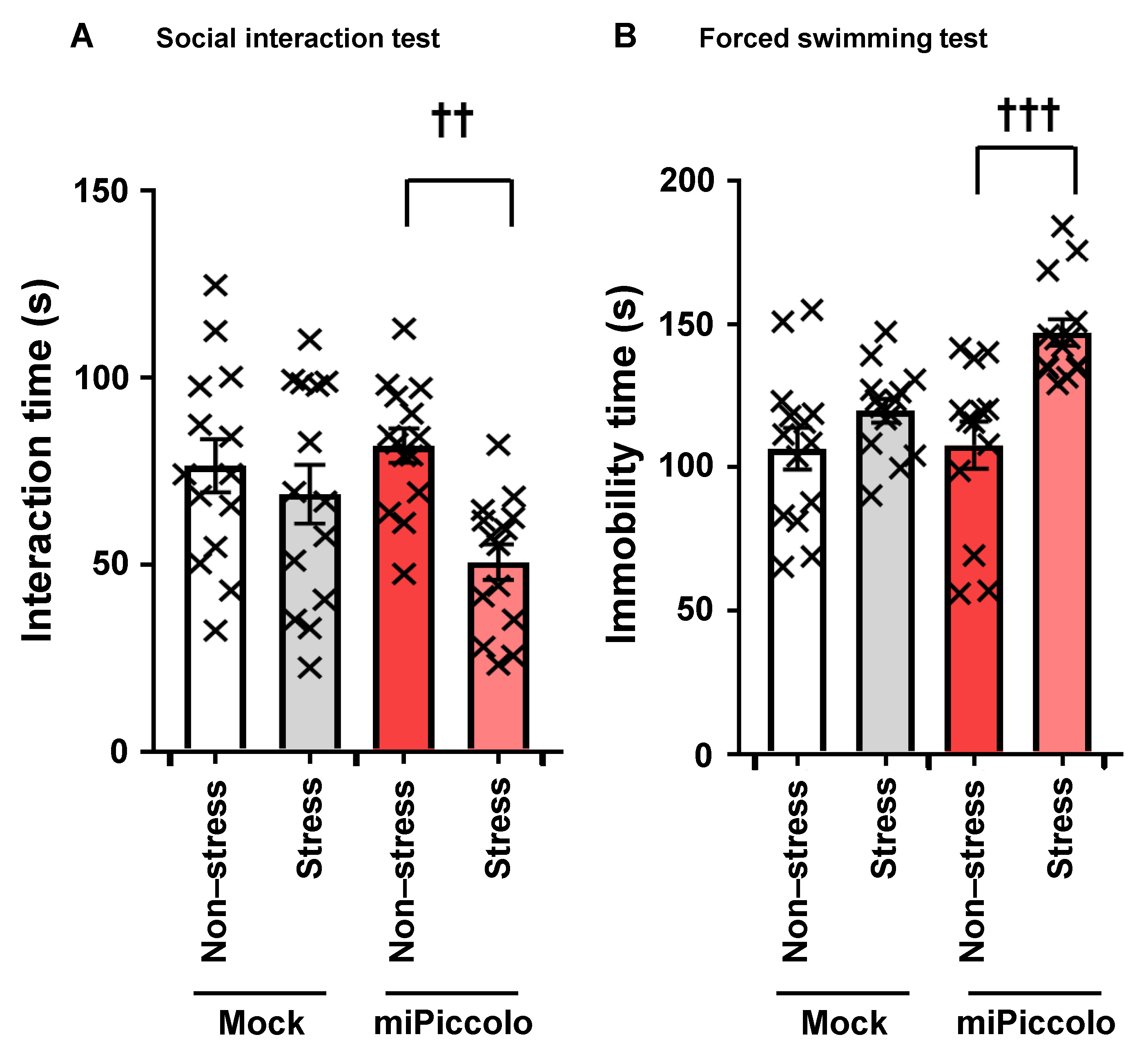
3.7. Stress Vulnerability Induced by Piccolo Suppression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearlson, G.D. Neurobiology of schizophrenia. Ann. Neurol. 2000, 48, 556–566. [Google Scholar] [CrossRef]
- Nuechterlein, K.H.; Barch, D.M.; Gold, J.M.; Goldberg, T.E.; Green, M.F.; Heaton, R.K. Identification of separable cognitive factors in schizophrenia. Schizophr. Res. 2004, 72, 29–39. [Google Scholar] [CrossRef]
- Rössler, W.; Salize, H.J.; van Os, J.; Riecher-Rössler, A. Size of burden of schizophrenia and psychotic disorders. Eur. Neuropsychopharmacol. 2005, 15, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Kani, A.S.; Shinn, A.K.; Lewandowski, K.E.; Öngür, D. Converging effects of diverse treatment modalities on frontal cortex in schizophrenia: A review of longitudinal functional magnetic resonance imaging studies. J. Psychiatr. Res. 2017, 84, 256–276. [Google Scholar] [CrossRef][Green Version]
- van Rossum, J.M. The significance of dopamine receptor blockade for the mechanism of action of neuroleptic drugs. Arch. Int. Pharmacodyn. Ther. 1966, 160, 492–494. [Google Scholar] [PubMed]
- Weinberger, D.; Laruelle, M. Neurochemical and neuropharmacological imaging in schizophrenia. In Neuropsychopharmacology—The Fifth Generation of Progress; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 833–855. [Google Scholar]
- Benes, F.M.; McSparren, J.; Bird, E.D.; SanGiovanni, J.P.; Vincent, S.L. Deficits in small interneurons in prefrontal and cingulate cortices of schizophrenic and schizoaffective patients. Arch. Gen. Psychiatry 1991, 48, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, D.R.; Egan, M.F.; Bertolino, A.; Callicott, J.H.; Mattay, V.S.; Lipska, B.K.; Berman, K.F.; Goldberg, T.E. Prefrontal neurons and the genetics of schizophrenia. Biol. Psychiatry 2001, 50, 825–844. [Google Scholar] [CrossRef]
- Walton, E.; Hibar, D.P.; Van Erp, T.G.M.; Potkin, S.G.; Roiz-Santiañez, R.; Crespo-Facorro, B.; Suarez-Pinilla, P.; Van Haren, N.E.M.; De Zwarte, S.M.C.; Kahn, R.S.; et al. Positive symptoms associate with cortical thinning in the superior temporal gyrus via the ENIGMA schizophrenia consortium. Acta Psychiatr. Scand. 2017, 135, 439–447. [Google Scholar] [CrossRef]
- Dieck, S.T.; Sanmartí-Vila, L.; Langnaese, K.; Richter, K.; Kindler, S.; Soyke, A.; Wex, H.; Smalla, K.-H.; Kämpf, U.; Fränzer, J.-T.; et al. Bassoon, a novel zinc-finger CAG/glutamine-repeat protein selectively localized at the active zone of presynaptic nerve terminals. J. Cell Biol. 1998, 142, 499–509. [Google Scholar] [CrossRef]
- Fenster, S.; Chung, W.J.; Zhai, R.; Cases-Langhoff, C.; Voss, B.; Garner, A.M.; Kaempf, U.; Kindler, S.; Gundelfinger, E.D.; Garner, C. Piccolo, a presynaptic zinc finger protein structurally related to Bassoon. Neuron 2000, 25, 203–214. [Google Scholar] [CrossRef]
- Gerber, S.H.; García, J.; Rizo, J.; Südhof, T.C. An unusual C2-domain in the active-zone protein piccolo: Implications for Ca2+ regulation of neurotransmitter release. EMBO J. 2001, 20, 1605–1619. [Google Scholar] [CrossRef]
- García, J.; Gerber, S.H.; Sugita, S.; Südhof, T.C.; Rizo, J. A conformational switch in the Piccolo C2A domain regulated by alternative splicing. Nat. Struct. Mol. Biol. 2003, 11, 45–53. [Google Scholar] [CrossRef]
- Shibasaki, T.; Sunaga, Y.; Fujimoto, K.; Kashima, Y.; Seino, S. Interaction of ATP sensor, cAMP sensor, Ca2+ sensor, and voltage-dependent Ca2+ channel in insulin granule exocytosis. J. Biol. Chem. 2004, 279, 7956–7961. [Google Scholar] [CrossRef]
- Wang, X.; Kibschull, M.; Laue, M.M.; Lichte, B.; Petrasch-Parwez, E.; Kilimann, M.W. Aczonin, a 550-Kd putative scaffolding protein of presynaptic active zones, shares homology regions with Rim and Bassoon and binds profilin. J. Cell Biol. 1999, 147, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef]
- Wang, X.; Hu, B.; Zieba, A.; Neumann, N.G.; Kasper-Sonnenberg, M.; Honsbein, A.; Hultqvist, G.; Conze, T.; Witt, W.; Limbach, C.; et al. A protein interaction node at the neurotransmitter release site: Domains of Aczonin/Piccolo, Bassoon, CAST, and Rim converge on the N-terminal domain of Munc13-1. J. Neurosci. 2009, 29, 12584–12596. [Google Scholar] [CrossRef] [PubMed]
- Ibi, D.; Nitta, A.; Ishige, K.; Cen, X.; Ohtakara, T.; Nabeshima, T.; Ito, Y. Piccolo knockdown-induced impairments of spatial learning and long-term potentiation in the hippocampal CA1 region. Neurochem. Int. 2010, 56, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Cen, X.; Nitta, A.; Ibi, D.; Zhao, Y.; Niwa, M.; Taguchi, K.; Hamada, M.; Ito, Y.; Wang, L.; Nabeshima, T. Identification of Piccolo as a regulator of behavioral plasticity and dopamine transporter internalization. Mol. Psychiatry 2008, 13, 451–463. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Seo, S.; Takayama, K.; Uno, K.; Ohi, K.; Hashimoto, R.; Nishizawa, D.; Ikeda, K.; Ozaki, N.; Nabeshima, T.; Miyamoto, Y.; et al. Functional analysis of deep intronic SNP rs13438494 in intron 24 of PCLO gene. PLoS ONE 2013, 8, e76960. [Google Scholar] [CrossRef]
- Uno, K.; Nishizawa, D.; Seo, S.; Takayama, K.; Matsumura, S.; Sakai, N.; Ohi, K.; Nabeshima, T.; Hashimoto, R.; Ozaki, N.; et al. The Piccolo intronic single nucleotide polymorphism rs13438494 Regulates dopamine and serotonin uptake and shows associations with dependence-like behavior in genomic association study. Curr. Mol. Med. 2015, 15, 265–274. [Google Scholar] [CrossRef]
- Choi, K.H.; Higgs, B.W.; Wendland, J.R.; Song, J.; McMahon, F.J.; Webster, M.J. Gene expression and genetic variation data implicate PCLO in bipolar disorder. Biol. Psychiatry 2011, 69, 353–359. [Google Scholar] [CrossRef]
- Giniatullina, A.; Maroteaux, G.; Geerts, C.; Koopmans, B.; Loos, M.; Klaassen, R.; Chen, N.; Van Der Schors, R.; Van Nierop, P.; Li, K.W.; et al. Functional characterization of the PCLO p. Ser4814Ala variant associated with major depressive disorder reveals cellular but not behavioral differences. Neuroscience 2015, 300, 518–538. [Google Scholar] [CrossRef]
- Sullivan, P.F.; De Geus, E.J.C.; Willemsen, G.; James, M.R.; Smit, J.H.; Zandbelt, T.; Arolt, V.; Baune, B.T.; Blackwood, D.; Cichon, S.; et al. Genome-wide association for major depressive disorder: A possible role for the presynaptic protein Piccolo. Mol. Psychiatry 2008, 14, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Igata, R.; Katsuki, A.; Kakeda, S.; Watanabe, K.; Igata, N.; Hori, H.; Konishi, Y.; Atake, K.; Kawasaki, Y.; Korogi, Y.; et al. PCLO rs2522833-mediated gray matter volume reduction in patients with drug-naive, first-episode major depressive disorder. Transl. Psychiatry 2017, 7, e1140. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.; Artero, S.; Carrière, I.; Maller, J.J.; Meslin, C.; Ritchie, K.; Ancelin, M.-L. GWAS-identified risk variants for major depressive disorder: Preliminary support for an association with late-life depressive symptoms and brain structural alterations. Eur. Neuropsychopharmacol. 2016, 26, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Schott, B.H.; Assmann, A.; Schmierer, P.; Soch, J.; Erk, S.; Garbusow, M.; Mohnke, S.; Pöhland, L.; Romanczuk-Seiferth, N.; Barman, A.; et al. Epistatic interaction of genetic depression risk variants in the human subgenual cingulate cortex during memory encoding. Transl. Psychiatry 2014, 4, e372. [Google Scholar] [CrossRef] [PubMed]
- Woudstra, S.; Bochdanovits, Z.; Van Tol, M.-J.; Veltman, D.J.; Zitman, F.G.; Van Buchem, M.A.; Van Der Wee, N.J.; Opmeer, E.M.; Demenescu, L.R.; Aleman, A.; et al. Piccolo genotype modulates neural correlates of emotion processing but not executive functioning. Transl. Psychiatry 2012, 2, e99. [Google Scholar] [CrossRef] [PubMed]
- Krzyżosiak, A.; Szyszka-Niagolov, M.; Wietrzych, M.; Gobaille, S.; Muramatsu, S.-I.; Krezel, W. Retinoid X receptor gamma control of affective behaviors involves dopaminergic signaling in mice. Neuron 2010, 66, 908–920. [Google Scholar] [CrossRef] [PubMed]
- Iida, A.; Takino, N.; Miyauchi, H.; Shimazaki, K.; Muramatsu, S.-I. Systemic delivery of tyrosine-mutant AAV vectors results in robust transduction of neurons in adult mice. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K. The Mouse Brain in Stereotaxic Coordinates: Compact, 3rd ed.; Elsevier: Amsterdam, The Netherland, 2008; ISBN 9780123742445. [Google Scholar]
- Fu, K.; Miyamoto, Y.; Sumi, K.; Saika, E.; Muramatsu, S.-I.; Uno, K.; Nitta, A. Overexpression of transmembrane protein 168 in the mouse nucleus accumbens induces anxiety and sensorimotor gating deficit. PLoS ONE 2017, 12, e0189006. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Iegaki, N.; Fu, K.; Ishikawa, Y.; Sumi, K.; Azuma, S.; Uno, K.; Muramatsu, S.-I.; Nitta, A. Striatal N-acetylaspartate synthetase Shati/Nat8l regulates depression-like behaviors via mGluR3-mediated serotonergic suppression in mice. Int. J. Neuropsychopharmacol. 2017, 20, 1027–1035. [Google Scholar] [CrossRef]
- Niwa, M.; Nitta, A.; Mizoguchi, H.; Ito, Y.; Noda, Y.; Nagai, T.; Nabeshima, T. A novel molecule “Shati” is involved in methamphetamine-induced hyperlocomotion, sensitization, and conditioned place preference. J. Neurosci. 2007, 27, 7604–7615. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Ishikawa, Y.; Iegaki, N.; Sumi, K.; Fu, K.; Sato, K.; Furukawa-Hibi, Y.; Muramatsu, S.-I.; Nabeshima, T.; Uno, K.; et al. Overexpression of Shati/Nat8l, an N-acetyltransferase, in the nucleus accumbens attenuates the response to methamphetamine via activation of group II mGluRs in mice. Int. J. Neuropsychopharmacol. 2014, 17, 1283–1294. [Google Scholar] [CrossRef]
- Lin, J.Y.; Lin, M.Z.; Steinbach, P.; Tsien, R.Y. Characterization of engineered channelrhodopsin variants with improved properties and kinetics. Biophys. J. 2009, 96, 1803–1814. [Google Scholar] [CrossRef]
- Alabi, A.A.; Tsien, R.W. Synaptic vesicle pools and dynamics. Cold Spring Harb. Perspect. Biol. 2012, 4, a013680. [Google Scholar] [CrossRef]
- Wang, H.-X.; Gao, W.-J. Cell type-specific development of NMDA receptors in the interneurons of rat prefrontal cortex. Neuropsychopharmacology 2009, 34, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, M.; Shi, J.; Zhou, Y.; Wan, Z.; Wang, Y.; Wan, Y.; Li, J.; Wang, Z.; Fei, J. Brain-specific SNAP-25 deletion leads to elevated extracellular glutamate level and schizophrenia-like behavior in mice. Neural Plast. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.; Waters, N.; Holm-Waters, S.; Tedroff, J.; Nilsson, M.; Carlsson, M.L. Interactions between monoamines, glutamate and GABA in schizophrenia: New evidence. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 237–260. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Miletich, R.S.; Kohn, P.D.; Esposito, G.; Carson, R.E.; Quarantelli, M.; Weinberger, D.R.; Berman, K.F. Reduced prefrontal activity predicts exaggerated striatal dopaminergic function in schizophrenia. Nat. Neurosci. 2002, 5, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Bartha, R.; Williamson, P.C.; Drost, D.J.; Malla, A.; Carr, T.J.; Cortese, L.; Canaran, G.; Rylett, R.J.; Neufeld, R.W.J. Measurement of glutamate and glutamine in the medial prefrontal cortex of never-treated schizophrenic patients and healthy controls by proton magnetic resonance spectroscopy. Arch. Gen. Psychiatry 1997, 54, 959–965. [Google Scholar] [CrossRef]
- Pilowsky, L.S.; Bressan, R.; Stone, J.M.; Erlandsson, K.; Mulligan, R.S.; Krystal, J.H.; Ell, P.J. First in vivo evidence of an NMDA receptor deficit in medication-free schizophrenic patients. Mol. Psychiatry 2005, 11, 118–119. [Google Scholar] [CrossRef]
- Paus, T.; Castro-Alamancos, M.A.; Petrides, M. Cortico-cortical connectivity of the human mid-dorsolateral frontal cortex and its modulation by repetitive transcranial magnetic stimulation. Eur. J. Neurosci. 2001, 14, 1405–1411. [Google Scholar] [CrossRef]
- Strafella, A.P.; Paus, T.; Barrett, J.; Dagher, A. Repetitive transcranial magnetic stimulation of the human prefrontal cortex induces dopamine release in the caudate nucleus. J. Neurosci. 2001, 21, RC157. [Google Scholar] [CrossRef]
- Uylings, H.B.; Groenewegen, H.J.; Kolb, B. Do rats have a prefrontal cortex? Behav. Brain Res. 2003, 146, 3–17. [Google Scholar] [CrossRef]
- Durstewitz, D.; Seamans, J.K. The dual-state theory of prefrontal cortex dopamine function with relevance to catechol-o-methyltransferase genotypes and schizophrenia. Biol. Psychiatry 2008, 64, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Suhara, T.; Suzuki, K.; Kobayashi, K.; Inoue, O.; Terasaki, O.; Someya, Y.; Sassa, T.; Sudo, Y.; Matsushima, E.; et al. Decreased prefrontal dopamine D1 receptors in schizophrenia revealed by PET. Nat. Cell Biol. 1997, 385, 634–636. [Google Scholar] [CrossRef]
- Davis, J.; Eyre, H.; Jacka, F.N.; Dodd, S.; Dean, O.; McEwen, S.; Debnath, M.; McGrath, J.; Maes, M.; Amminger, P.; et al. A review of vulnerability and risks for schizophrenia: Beyond the two hit hypothesis. Neurosci. Biobehav. Rev. 2016, 65, 185–194. [Google Scholar] [CrossRef]
- Thompson, P.M.; Sower, A.C.; Perrone-Bizzozero, N.I. Altered levels of the synaptosomal associated protein SNAP-25 in schizophrenia. Biol. Psychiatry 1998, 43, 239–243. [Google Scholar] [CrossRef]
- Karson, C.N.; Mrak, R.E.; Schluterman, K.O.; Sturner, W.Q.; Sheng, J.G.; Griffin, W.S.T. Alterations in synaptic proteins and their encoding mRNAs in prefrontal cortex in schizophrenia: A possible neurochemical basis for “hypofrontality”. Mol. Psychiatry 1999, 4, 39–45. [Google Scholar] [CrossRef][Green Version]
- Hashimoto, T.; Volk, D.W.; Eggan, S.M.; Mirnics, K.; Pierri, J.N.; Sun, Z.X.; Sampson, A.R.; Lewis, D.A. Gene expression defi-cits in a subclass of GABA neurons in the prefrontal cortex of subjects with schizophrenia. J. Neurosci. 2003, 23, 6315–6326. [Google Scholar] [CrossRef] [PubMed]
- Weidenhofer, J.; Bowden, N.A.; Scott, R.J.; Tooney, P.A. Altered gene expression in the amygdala in schizophrenia: Up-regulation of genes located in the cytomatrix active zone. Mol. Cell. Neurosci. 2006, 31, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Weidenhofer, J.; Scott, R.J.; Tooney, P.A. Investigation of the expression of genes affecting cytomatrix active zone function in the amygdala in schizophrenia: Effects of antipsychotic drugs. J. Psychiatr. Res. 2009, 43, 282–290. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nitta, A.; Izuo, N.; Hamatani, K.; Inagaki, R.; Kusui, Y.; Fu, K.; Asano, T.; Torii, Y.; Habuchi, C.; Sekiguchi, H.; et al. Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex. J. Pers. Med. 2021, 11, 607. https://doi.org/10.3390/jpm11070607
Nitta A, Izuo N, Hamatani K, Inagaki R, Kusui Y, Fu K, Asano T, Torii Y, Habuchi C, Sekiguchi H, et al. Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex. Journal of Personalized Medicine. 2021; 11(7):607. https://doi.org/10.3390/jpm11070607
Chicago/Turabian StyleNitta, Atsumi, Naotaka Izuo, Kohei Hamatani, Ryo Inagaki, Yuka Kusui, Kequan Fu, Takashi Asano, Youta Torii, Chikako Habuchi, Hirotaka Sekiguchi, and et al. 2021. "Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex" Journal of Personalized Medicine 11, no. 7: 607. https://doi.org/10.3390/jpm11070607
APA StyleNitta, A., Izuo, N., Hamatani, K., Inagaki, R., Kusui, Y., Fu, K., Asano, T., Torii, Y., Habuchi, C., Sekiguchi, H., Iritani, S., Muramatsu, S.-i., Ozaki, N., & Miyamoto, Y. (2021). Schizophrenia-Like Behavioral Impairments in Mice with Suppressed Expression of Piccolo in the Medial Prefrontal Cortex. Journal of Personalized Medicine, 11(7), 607. https://doi.org/10.3390/jpm11070607