Changes in the Diversity of Human Skin Microbiota to Cosmetic Serum Containing Prebiotics: Results from a Randomized Controlled Trial


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cosmetic Serum Formulation
2.2. Subjects and Sampling
2.3. Skin Assessments
2.4. DNA Extraction, Amplification, and Sequencing
2.5. Bioinformatic Analysis Using 16S rRNA Sequences
2.6. Statistical Analysis
3. Results
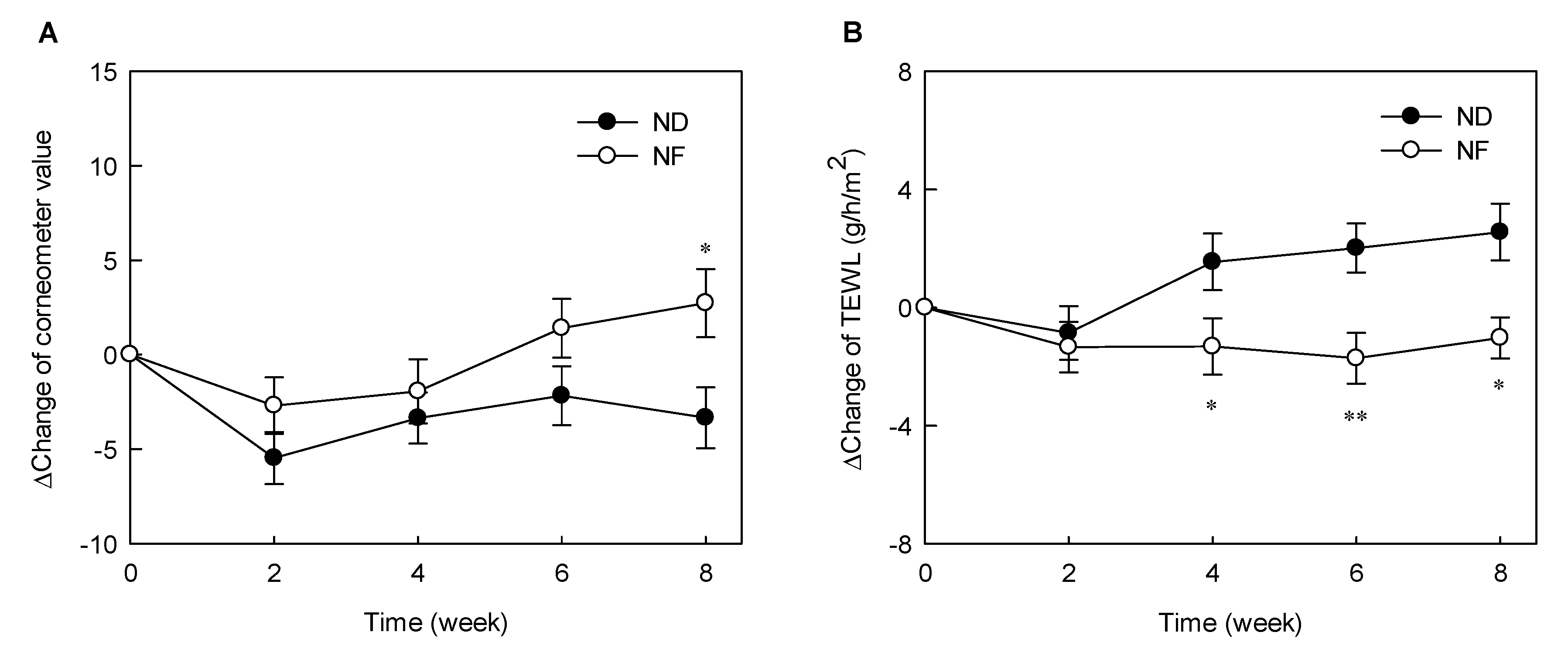
3.1. Changes in the Water Holding Capacity of the Skin Surface and TEWL by Prebiotics-Containing Cosmetic Serum Treatment
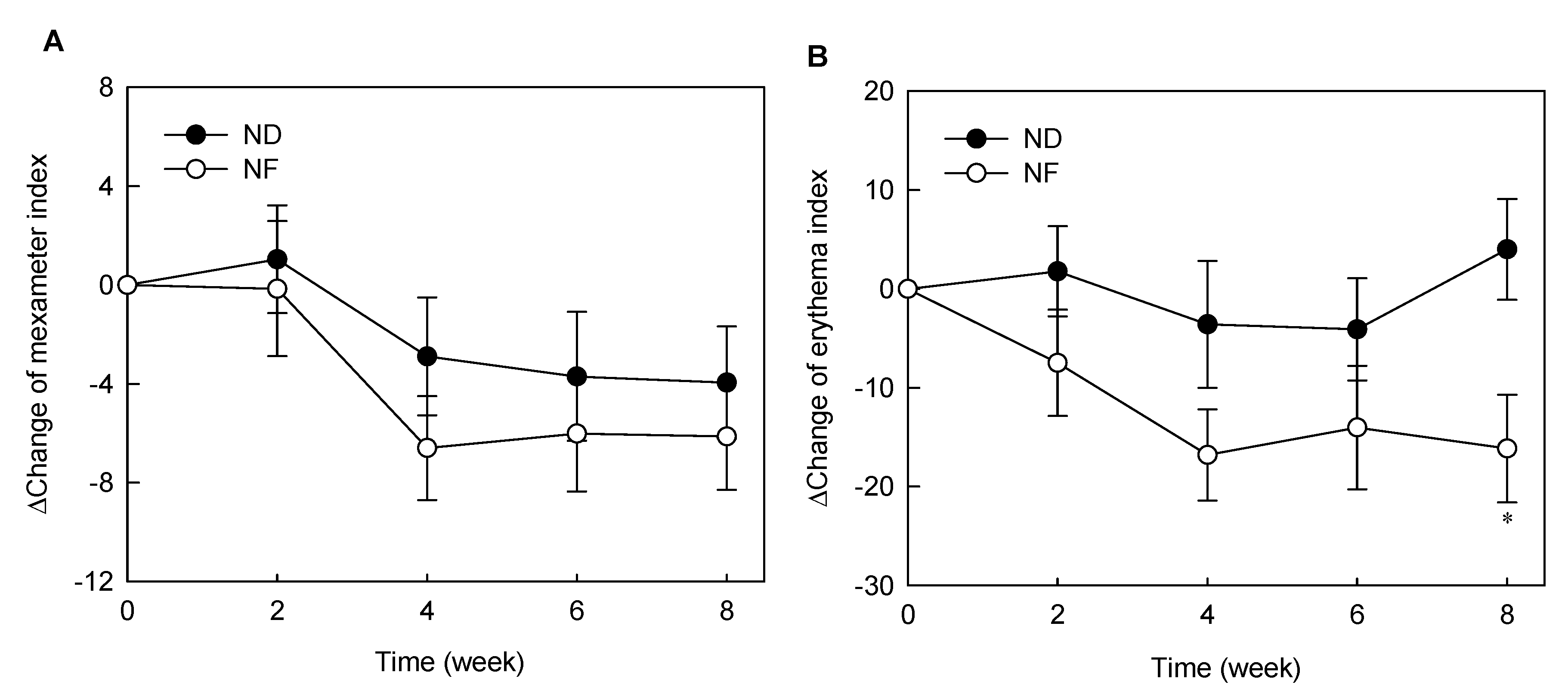
3.2. Changes in Skin Pigments by Prebiotics-Containing Cosmetic Serum Treatment
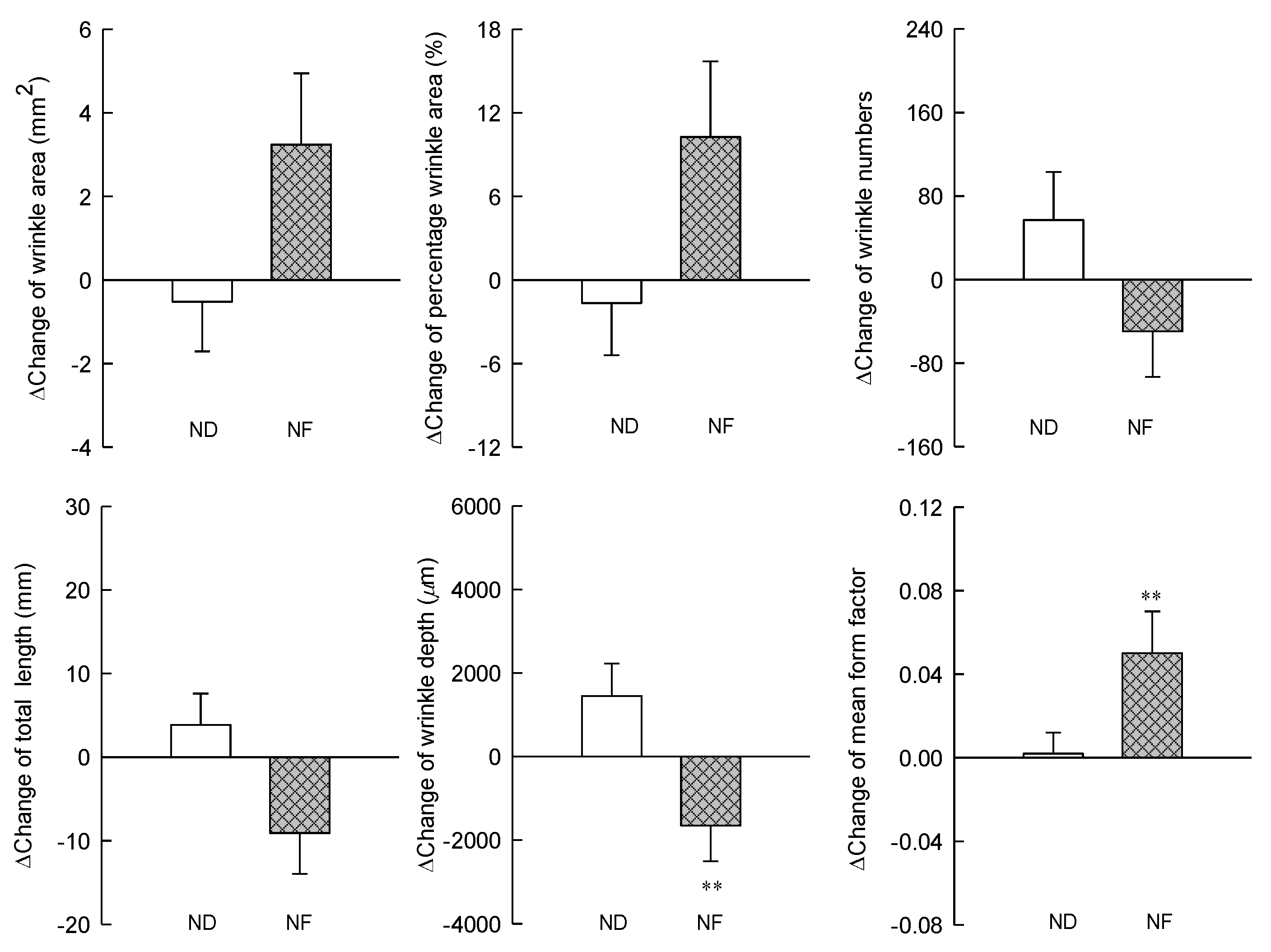
3.3. Changes in the Skin Wrinkles Using Replica by Prebiotics-Containing Cosmetic Serum Treatment
3.4. Changes in the Water Holding Capacity of the Skin Surface and TEWL by Prebiotics-Containing Cosmetic Serum Treatment
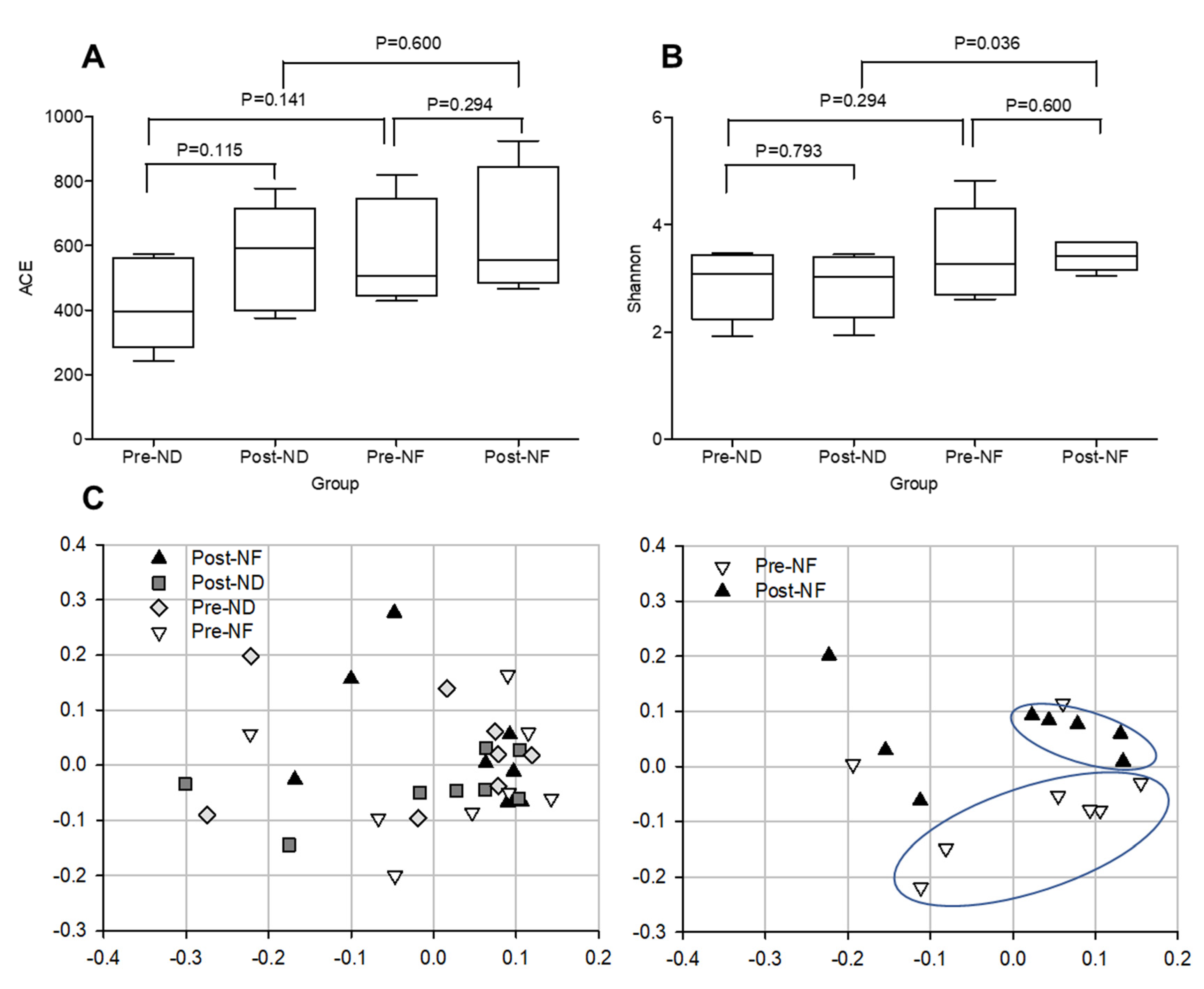
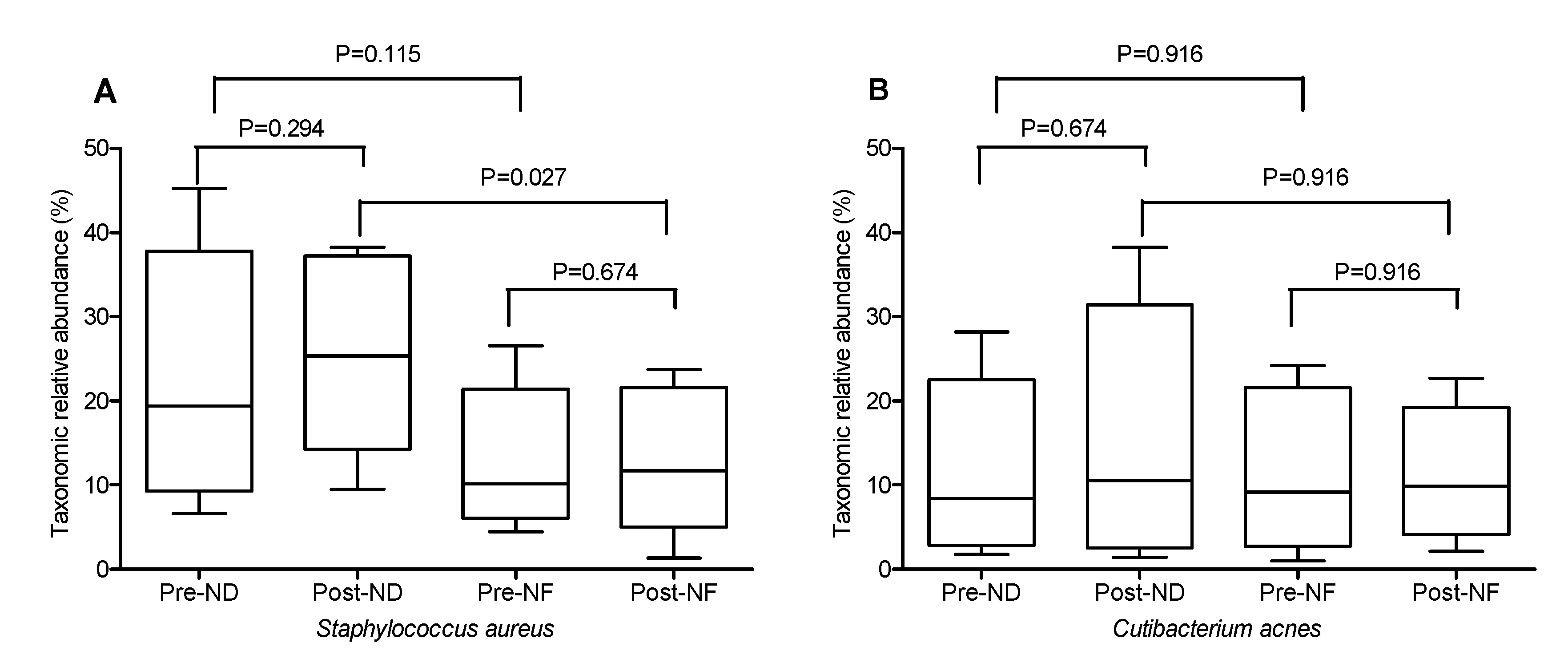
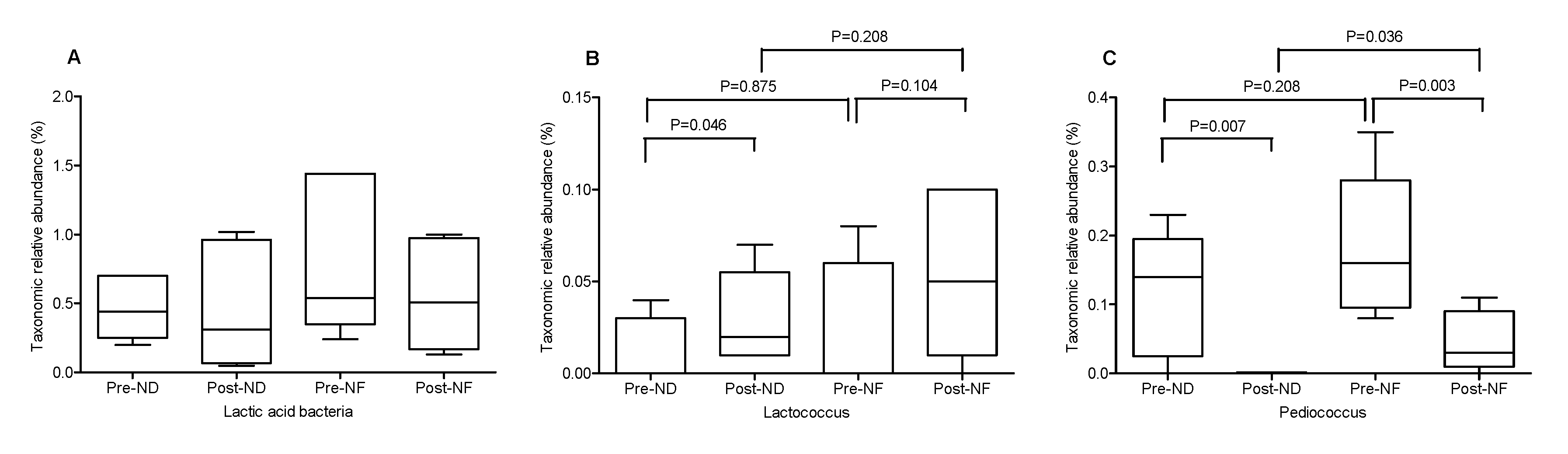
3.5. Microbial Diversity Within and between the Groups
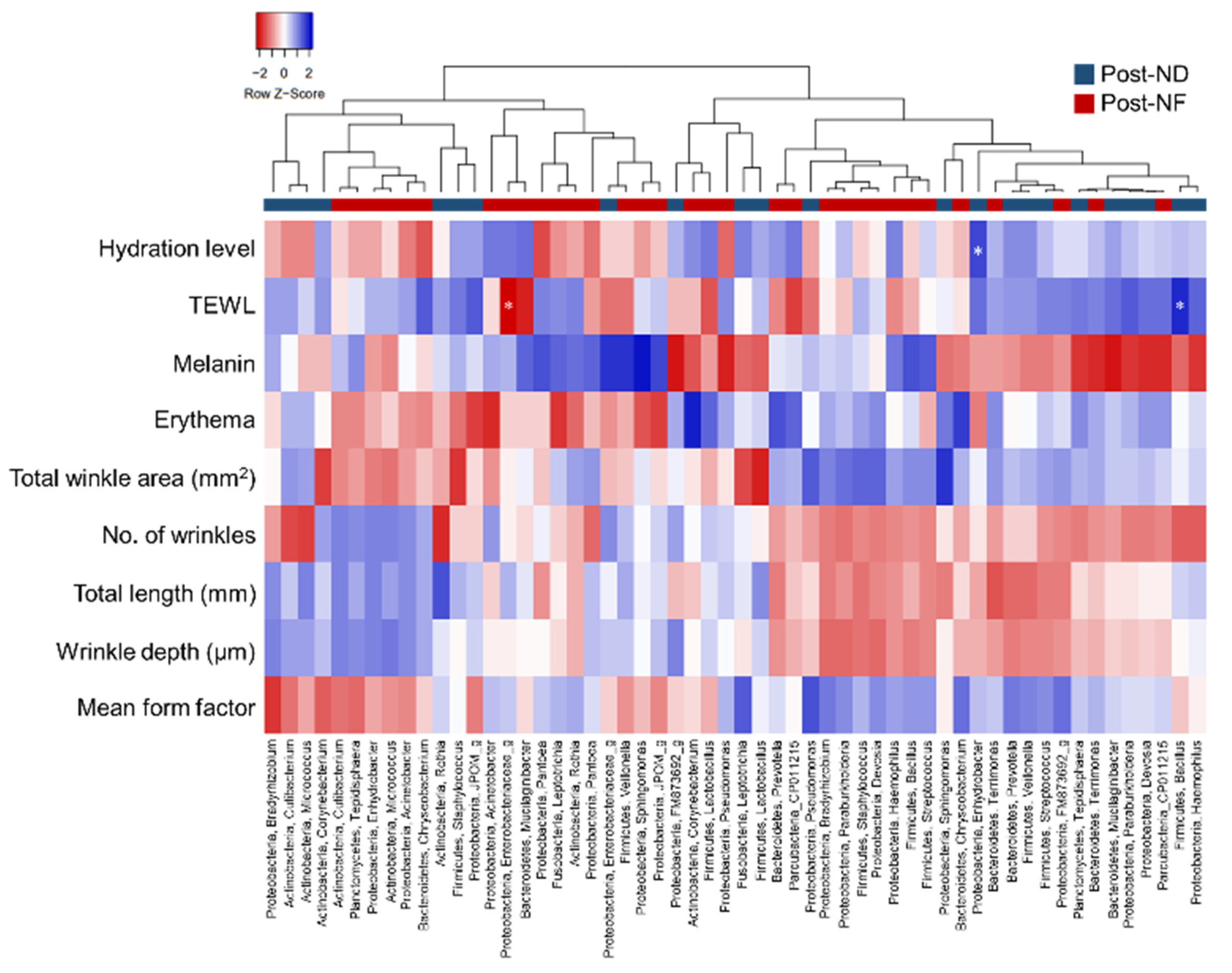
3.6. Correlation Analyses between Skin Microbiota and Skin Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dąbrowska, A.; Spano, F.; Derler, S.; Adlhart, C.; Spencer, N.; Rossi, R. The relationship between skin function, barrier properties, and body-dependent factors. Ski Res. Technol. 2018, 24, 165–174. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb. Ecol. 2018, 76, 113–120. [Google Scholar] [CrossRef]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota–host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, M.; Goldberg, D.; Aiello, A.; Larson, E.; Foxman, B. Skin microbiota: Microbial community structure and its potential association with health and disease. Infect. Genet. Evol. 2011, 11, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.A.; Wang, W.; Roux, C.M.; Beasley, F.C.; Heinrichs, D.E.; Dunman, P.M.; Magarvey, N.A. Staphylococcus aureus nonribosomal peptide secondary metabolites regulate virulence. Science 2010, 329, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Secor, P.R.; Jennings, L.K.; James, G.A.; Kirker, K.R.; Pulcini, E.D.; McInnerney, K.; Gerlach, R.; Livinghouse, T.; Hilmer, J.K.; Bothner, B.; et al. Phevalin (aureusimine B) production by Staphylococcus aureus biofilm and impacts on human keratinocyte gene expression. PLoS ONE 2012, 7, e40973. [Google Scholar] [CrossRef]
- Bockmühl, D.; Jassoy, C.; Nieveler, S.; Scholtyssek, R.; Wadle, A.; Waldmann-Laue, M. Prebiotic cosmetics: An alternative to antibacterial products. Int. J. Cosmet. Sci. 2007, 29, 63–64. [Google Scholar] [CrossRef]
- Ouwehand, A.; Lahtinen, S.; Tiihonen, K. The Potential of Probiotics and Prebiotics for Skin Health. In Textbook of Aging Skin; Springer: Berlin/Heidelberg, Germany, 2010; pp. 799–810. [Google Scholar]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Hong, Y.H.; Chang, U.J.; Kim, Y.S.; Jung, E.Y.; Suh, H.J. Dietary galacto-oligosaccharides improve skin health: A randomized double blind clinical trial. Asia Pac. J. Clin. Nutr. 2017, 26, 613–618. [Google Scholar] [CrossRef]
- Di Mauro, M.D.; Tomasello, B.; Giardina, R.C.; Dattilo, S.; Mazzei, V.; Sinatra, F.; Caruso, M.; D’Antona, N.; Renis, M. Sugar and mineral enriched fraction from olive mill wastewater for promising cosmeceutical application: Characterization, in vitro and in vivo studies. Food Funct. 2017, 8, 4713–4722. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.; Redmond, M.; Röding, J. Anti-wrinkle therapy: Significant new findings in the non-invasive cosmetic treatment of skin wrinkles with beta-glucan. Int. J. Cosmet. Sci. 2005, 27, 292. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B. The Validity and practicality of sun-reactive skin type-I through type-Vi. Arch. Dermatol. 1988, 124, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Quince, C.; Lanzen, A.; Curtis, T.P.; Davenport, R.J.; Hall, N.; Head, I.M.; Read, L.F.; Sloan, W.T. Accurate determination of microbial diversity from 454 pyrosequencing data. Nat. Methods 2009, 6, 639–641. [Google Scholar] [CrossRef]
- Soergel, D.A.W.; Dey, N.; Knight, R.; Brenner, S.E. Selection of primers for optimal taxonomic classification of environmental 16S rRNA gene sequences. ISME J. 2012, 6, 1440–1444. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Mount, D.W. Using the basic local alignment search tool (BLAST). Cold Spring Harb. Protoc. 2007, 2007. [Google Scholar] [CrossRef]
- Houser, T.; Zerweck, C.; Grove, G.; Wickett, R. Shadow analysis via the C+ K Visioline: A technical note. Ski Res. Technol. 2017, 23, 447–451. [Google Scholar] [CrossRef]
- Suh, M.G.; Hong, Y.H.; Jung, E.Y.; Suh, H.J. Inhibitory Effect of Galactooligosaccharide on Skin Pigmentation. Prev. Nutr. Food Sci. 2019, 24, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.Y.; Kwon, J.I.; Hong, Y.H.; Suh, H.J. Evaluation of Anti-Wrinkle Effects of DuOligo, Composed of Lactulose and Galactooligosaccharides. Prev. Nutr. Food Sci. 2017, 22, 381–384. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alberts, B.; Bray, D.; Lewis, J.; Raff, M.; Roberts, K.; Watson, J. Molecular Biology of the Cell; Biochemistry and Molecular Biology Education: Garland, NY, USA, 1994; pp. 907–982. [Google Scholar]
- Verdier-Sévrain, S.; Bonté, F. Skin hydration: A review on its molecular mechanisms. J. Cosmet. Dermatol. 2007, 6, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Izawa, N.; Sone, T. Cosmetic ingredients fermented by lactic acid bacteria. In Microbial Production; Springer: Berlin, Germany, 2014; pp. 233–242. [Google Scholar]
- Hong, K.-B.; Jeong, M.; Han, K.S.; Hwan Kim, J.; Park, Y.; Suh, H.J. Photoprotective effects of galacto-oligosaccharide and/or Bifidobacterium longum supplementation against skin damage induced by ultraviolet irradiation in hairless mice. Int. J. Food Sci. Nutr. 2015, 66, 923–930. [Google Scholar] [CrossRef]
- Dekio, I.; Hayashi, H.; Sakamoto, M.; Kitahara, M.; Nishikawa, T.; Suematsu, M.; Benno, Y. Detection of potentially novel bacterial components of the human skin microbiota using culture-independent molecular profiling. J. Med. Microbiol. 2005, 54, 1231–1238. [Google Scholar] [CrossRef]
- Gao, Z.; Perez-Perez, G.I.; Chen, Y.; Blaser, M.J. Quantitation of major human cutaneous bacterial and fungal populations. J. Clin. Microbiol. 2010, 48, 3575–3581. [Google Scholar] [CrossRef]
- La Fata, G.; Rastall, R.A.; Lacroix, C.; Harmsen, H.J.M.; Mohajeri, M.H.; Weber, P.; Steinert, R.E. Recent Development of Prebiotic Research-Statement from an Expert Workshop. Nutrients 2017, 9, 1376. [Google Scholar] [CrossRef]
- von Eiff, C.; Becker, K.; Machka, K.; Stammer, H.; Peters, G.; Grp, S. Nasal carriage as a source of Staphylococcus aureus bacteremia. N. Engl. J. Med. 2001, 344, 11–16. [Google Scholar] [CrossRef]
- Weidenmaier, C.; Goerke, C.; Wolz, C. Staphylococcus aureus determinants for nasal colonization. Trends Microbiol. 2012, 20, 243–250. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Two, A.M.; Chun, K.A.; Narala, S.; Geha, R.S.; Hata, T.R.; Gallo, R.L. Staphylococcus aureus Exploits Epidermal Barrier Defects in Atopic Dermatitis to Trigger Cytokine Expression. J. Investig. Dermatol. 2016, 136, 2192–2200. [Google Scholar] [CrossRef]
- Hirasawa, Y.; Takai, T.; Nakamura, T.; Mitsuishi, K.; Gunawan, H.; Suto, H.; Ogawa, T.; Wang, X.L.; Ikeda, S.; Okumura, K.; et al. Staphylococcus aureus Extracellular Protease Causes Epidermal Barrier Dysfunction. J. Investig. Dermatol. 2010, 130, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Kemter, A.M.; Nagler, C.R. Influences on allergic mechanisms through gut, lung, and skin microbiome exposures. J. Clin. Investig. 2019, 129, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.L.; Nakatsuji, T. Microbial Symbiosis with the Innate Immune Defense System of the Skin. J. Investig. Dermatol. 2011, 131, 1974–1980. [Google Scholar] [CrossRef]
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Schmidt, R.R.; von Aulock, S.; et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Gotz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin commensals amplify the innate immune response to pathogens by activation of distinct signaling pathways. J. Investig. Dermatol. 2011, 131, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.; Solyman, S.; Yones, M.; Halaby, H.; Abdalla, Y.; Hanora, A. Shotgun Metagenomic analysis of cutaneous microbiome in patients with atopic dermatitis. Rec. Pharm. Biomed. Sci. 2019, 3, 1–3. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ohishi, H.; Kakizoe, K.; Benno, Y. Faecal microbiota and secretory immunogloblin a levels in adult patients with atopic dermatitis. Microb. Ecol. Health Dis. 2004, 16, 13–17. [Google Scholar]
- Yap, G.C.; Loo, E.X.L.; Aw, M.; Lu, Q.; Shek, L.P.-C.; Lee, B.W. Molecular analysis of infant fecal microbiota in an Asian at-risk cohort–correlates with infant and childhood eczema. BMC Res. Notes 2014, 7, 166. [Google Scholar] [CrossRef]
- Suh, M.G.; Bae, G.Y.; Jo, K.; Kim, J.M.; Hong, K.B.; Suh, H.J. Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway. Molecules 2020, 25, 1679. [Google Scholar] [CrossRef]
- Holland, K.T.; Bojar, R.A. Cosmetics. Am. J. Clin. Dermatol. 2002, 3, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic-acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–86. [Google Scholar] [CrossRef]
- Oh, S.; Kim, S.H.; Ko, Y.; Sim, J.H.; Kim, K.S.; Lee, S.H.; Park, S.; Kim, Y.J. Effect of bacteriocin produced by Lactococcus sp HY 449 on skin-inflammatory bacteria. Food Chem. Toxicol. 2006, 44, 1184–1190. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, K.-B.; Hong, Y.H.; Jung, E.Y.; Jo, K.; Suh, H.J. Changes in the Diversity of Human Skin Microbiota to Cosmetic Serum Containing Prebiotics: Results from a Randomized Controlled Trial. J. Pers. Med. 2020, 10, 91. https://doi.org/10.3390/jpm10030091
Hong K-B, Hong YH, Jung EY, Jo K, Suh HJ. Changes in the Diversity of Human Skin Microbiota to Cosmetic Serum Containing Prebiotics: Results from a Randomized Controlled Trial. Journal of Personalized Medicine. 2020; 10(3):91. https://doi.org/10.3390/jpm10030091
Chicago/Turabian StyleHong, Ki-Bae, Yang Hee Hong, Eun Young Jung, Kyungae Jo, and Hyung Joo Suh. 2020. "Changes in the Diversity of Human Skin Microbiota to Cosmetic Serum Containing Prebiotics: Results from a Randomized Controlled Trial" Journal of Personalized Medicine 10, no. 3: 91. https://doi.org/10.3390/jpm10030091
APA StyleHong, K.-B., Hong, Y. H., Jung, E. Y., Jo, K., & Suh, H. J. (2020). Changes in the Diversity of Human Skin Microbiota to Cosmetic Serum Containing Prebiotics: Results from a Randomized Controlled Trial. Journal of Personalized Medicine, 10(3), 91. https://doi.org/10.3390/jpm10030091