
Circulating Cell-Free DNA as an Epigenetic Biomarker for Early Diabetic Retinopathy: A Narrative Review

 , , ,
, , ,  ,
,
Abstract
1. Introduction
1.1. Background on Diabetic Retinopathy
1.2. Importance of Early Detection
1.3. Emerging Role of Epigenetics
1.4. Circulating cfDNA as a Biomarker for DR
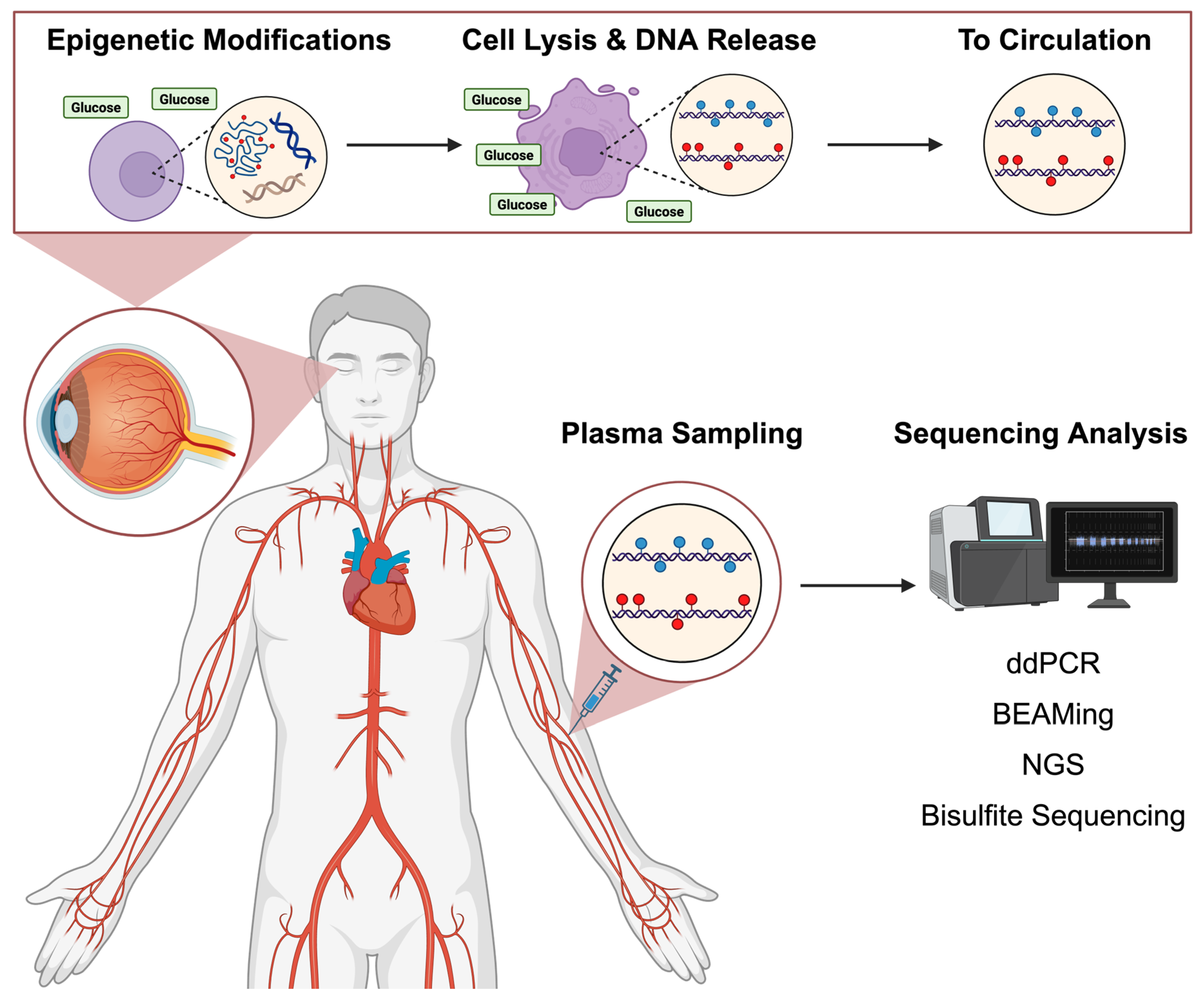
2. Pathophysiology of DR and cfDNA Origin
2.1. Vascular and Neural Apoptosis Contributing to the Accumulation of cfDNA
2.2. DR Epigenetic Dysregulation
3. Epigenetic Signatures in DR
3.1. DNA Methylation Signatures
3.2. Histone Modification Signatures and Changes to Gene Expression

{kind=link}
| Author and Year | Origin | Sample | Marker | T2D Groups (n) | Control Group (n) | Main Results |
|---|---|---|---|---|---|---|
| Han et al., 2021 [32] | China | Plasma | MESP1, LY6G6D, and LINC01556 | DR (35) | Age-, gender-, and diabetic duration-matched T2DM (35) | Three-gene signature active expression associated with DR. |
| Maghbooli et al., 2015 [60] | Iran | Peripheral blood leukocytes | Global DNA methylation (5-methylcytosine content) | PDR, NPDR (74), NDR (94) | None | Increased global DNA methylation associated with PDR, NPDR, and NDR. |
| Nunes et al., 2017 [65] | Brazil | Peripheral blood leukocytes | Methylation of MTHFR-promoter and polymorphism of 1298AA of MTHFR | DR (16), DN (29) | T2DM with no complications (60) | MTHFR-promoter hypermethylation associated with DR. |
| Bezerra et al., 2019 [66] | Brazil | Peripheral blood leukocytes | Methylation of MTHFR-promoter and polymorphism of C677T and A1298C of MTHFR | DR (22), NDR (25) | T2DM with no complications (60) | MTHFR-promoter hypermethylation with 1298AA polymorphism is associated with higher glycemia, LDL cholesterol, and total cholesterol. |
| Nunes et al., 2018 [68] | Brazil | Peripheral blood leukocytes | Methylation of miR-9-3-, miR-34a-, and miR-137-promoters | DR (19), DN (29) | T2DM with no complications (60) | miR-9-3-promoter hypermethylation associated with increased risk of DR. miR-137-promoter hypermethylation associated with protective effects, reducing microvascular diabetes complications. |
| Duraisamy et al., 2019 [69] | USA | Blood | MHL1 and SOD2 hypermethylation | PDR (23) | Non-DR-T2DM (23); healthy control (15). | Higher 5mC levels in the SOD2-promoter resulted in a 50% decrease in SOD2 mRNA in PDR and a 20% decrease in non-DR-T2DM. |
| Yang et al., 2022 [73] | China | Blood | ZDHHC23 and SLC25A21 hypermethylation | T2DM with DR (43) | T2DM without DR (92) | Hypermethylation of cg12869254 and cg04026387, containing the ZDHHC23 and SLC25A21 genes |
| Han et al., 2021 [32] | China | Blood | H3K4me1 | DR (35) | Age-, gender-, and diabetic duration-matched T2DM (35) | H3K4me1 active expression associated with DR. |
4. CfDNA in Detection Technologies
4.1. Origin of cfDNA in Circulation
4.2. Tissue-Specific cfDNA from Retinal Cells
4.3. Detection and Quantification Techniques
4.3.1. DdPCR
4.3.2. BEAMing
4.3.3. NGS
4.3.4. Bisulfite Sequencing
| Analysis | Type | Sensitivity | Target | Advantages | Limitations | First Author and Year |
|---|---|---|---|---|---|---|
| DdPCR | Targeted | ~0.01% | Known epigenetic markers | High sensitivity; low DNA input; fast results | Limited to known targets, limited multiplex ability | Medina et al., 2023; Hindson et al., 2013 [27,84] |
| BEAMing | Targeted (Digital PCR) | ~0.01% | Rare epigenetic variants | High accuracy; detects ultra-low variant levels | Requires specialized equipment | Diehl et al., 2008; Pircalabioru et al., 2024 [85,86] |
| NGS (Targeted) | Targeted | Moderate | Specific CpG sites | Broad analysis; scalable | Requires assay design; longer processing time | Medina et al., 2023; Shinjo et al., 2020; Koval et al., 2021 [27,80,88] |
| NGS (Genome-wide) | Unbiased | High | Entire methylome | Discovery of novel markers | High DNA input; resource-intensive | Medina et al., 2023; Shen et al., 2019 [27,87] |
| Bisulfite Sequencing | Targeted or genome-wide | High | Single-CpG resolution | Quantitative and specific | DNA degradation; cannot distinguish 5mC from 5hmC | Medina et al., 2023; Koval et al., 2021; Zhang et al., 2015 [27,88,90] |
5. Current Limitations and Future Directions
5.1. Limitations in cfDNA Sequencing
5.2. Epigenetic Variability and the Future of cfDNA in Precision Medicine
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DR | Diabetic retinopathy |
| T2DM | Type 2 diabetes mellitus |
| cfDNA | Circulating cell-free DNA |
| ERG | Electroretinography |
| mtDNA | Mitochondrial DNA |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| AGE | Advanced glycation end-product |
| MTHFR | Methylenetetrahydrofolate reductase |
| 5hmC | 5-hydroxymethylcytosine |
| CAD | Caspase-activated DNase |
| ddPCR | Digital droplet PCR |
| BEAMing | Magnetic bead-based enrichment |
| NGS | Next-generation sequencing |
| TAm-Seq | Tagged-amplicon deep sequencing |
| PDR | Proliferative diabetic retinopathy |
| NPDR | Non-proliferative diabetic retinopathy |
| NDR | Not diabetic retinopathy |
| DN | Diabetic nephropathy |
References
- Yau, J.W.Y.; Rogers, S.L.; Kawasaki, R.; Lamoureux, E.L.; Kowalski, J.W.; Bek, T.; Chen, S.-J.; Dekker, J.M.; Fletcher, A.; Grauslund, J.; et al. Global Prevalence and Major Risk Factors of Diabetic Retinopathy. Diabetes Care 2012, 35, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.; Klein, B.E.; Moss, S.E.; Davis, M.D.; DeMets, D.L. The Wisconsin Epidemiologic Study of Diabetic Retinopathy. III. Prevalence and Risk of Diabetic Retinopathy When Age at Diagnosis Is 30 or More Years. Arch. Ophthalmol. 1984, 102, 527–532. [Google Scholar] [CrossRef]
- Cai, J.; Boulton, M. The Pathogenesis of Diabetic Retinopathy: Old Concepts and New Questions. Eye 2002, 16, 242–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Wong, T.Y.; Sabanayagam, C. Epidemiology of Diabetic Retinopathy, Diabetic Macular Edema and Related Vision Loss. Eye Vis. 2015, 2, 17. [Google Scholar] [CrossRef]
- Zeng, Y.; Cao, D.; Yu, H.; Yang, D.; Zhuang, X.; Hu, Y.; Li, J.; Yang, J.; Wu, Q.; Liu, B.; et al. Early Retinal Neurovascular Impairment in Patients with Diabetes without Clinically Detectable Retinopathy. Br. J. Ophthalmol. 2019, 103, 1747–1752. [Google Scholar] [CrossRef]
- Oh, K.; Kang, H.M.; Leem, D.; Lee, H.; Seo, K.Y.; Yoon, S. Early Detection of Diabetic Retinopathy Based on Deep Learning and Ultra-Wide-Field Fundus Images. Sci. Rep. 2021, 11, 1897. [Google Scholar] [CrossRef]
- Sachdeva, M. Diabetic Retinopathy. Available online: https://www.hopkinsmedicine.org/health/conditions-and-diseases/diabetes/diabetic-retinopathy (accessed on 22 March 2025).
- Mackay, D.D.; Garza, P.S.; Bruce, B.B.; Newman, N.J.; Biousse, V. The Demise of Direct Ophthalmoscopy. Neurol. Clin. Pract. 2015, 5, 150–157. [Google Scholar] [CrossRef]
- Tabl, M.A. Early Detection of Neurodegeneration in Type 2 Diabetic Patients without Diabetic Retinopathy Using Electroretinogram and Spectral-Domain Optical Coherence Tomography. J. Egypt. Ophthalmol. Soc. 2020, 113, 26. [Google Scholar] [CrossRef]
- Zeng, Y.; Cao, D.; Yang, D.; Zhuang, X.; Yu, H.; Hu, Y.; Zhang, Y.; Yang, C.; He, M.; Zhang, L. Screening for Diabetic Retinopathy in Diabetic Patients with a Mydriasis-Free, Full-Field Flicker Electroretinogram Recording Device. Doc. Ophthalmol. 2020, 140, 211–220. [Google Scholar] [CrossRef]
- McAnany, J.J.; Liu, K.; Park, J.C. Electrophysiological Measures of Dysfunction in Early-Stage Diabetic Retinopathy: No Correlation between Cone Phototransduction and Oscillatory Potential Abnormalities. Doc. Ophthalmol. 2020, 140, 31–42. [Google Scholar] [CrossRef]
- Kim, M.; Kim, R.-Y.; Park, W.; Park, Y.-G.; Kim, I.-B.; Park, Y.-H. Electroretinography and Retinal Microvascular Changes in Type 2 Diabetes. Acta Ophthalmol. 2020, 98, e807–e813. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.; Gordon-Reid, A.; Shawkat, F.; Self, J.E. Comparison of the Handheld RETeval ERG System with a Routine ERG System in Healthy Adults and in Paediatric Patients. Eye 2021, 35, 2180–2189. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.P. Epigenetics: Principles and Practice. Dig. Dis. 2011, 29, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Drag, M.H.; Kilpeläinen, T.O. Cell-Free DNA and RNA—Measurement and Applications in Clinical Diagnostics with Focus on Metabolic Disorders. Physiol. Genom. 2021, 53, 33–46. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, H.; Wang, R.; Chen, Y.; Ouyang, X.; Li, W.; Sun, Y.; Peng, A. Cancer Epigenetics: From Laboratory Studies and Clinical Trials to Precision Medicine. Cell Death Discov. 2024, 10, 1–12. [Google Scholar] [CrossRef]
- Feehley, T.; O’Donnell, C.W.; Mendlein, J.; Karande, M.; McCauley, T. Drugging the Epigenome in the Age of Precision Medicine. Clin. Epigenetics 2023, 15, 6. [Google Scholar] [CrossRef]
- Cheng, Z.; Zheng, L.; Almeida, F.A. Epigenetic Reprogramming in Metabolic Disorders: Nutritional Factors and Beyond. J. Nutr. Biochem. 2018, 54, 1–10. [Google Scholar] [CrossRef]
- Wu, Y.-L.; Lin, Z.-J.; Li, C.-C.; Lin, X.; Shan, S.-K.; Guo, B.; Zheng, M.-H.; Li, F.; Yuan, L.-Q.; Li, Z. Epigenetic Regulation in Metabolic Diseases: Mechanisms and Advances in Clinical Study. Signal Transduct. Target. Ther. 2023, 8, 1–27. [Google Scholar] [CrossRef]
- Arguelles, A.O.; Meruvu, S.; Bowman, J.D.; Choudhury, M. Are Epigenetic Drugs for Diabetes and Obesity at Our Door Step? Drug Discov. Today 2016, 21, 499–509. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Kowluru, A.; Mishra, M.; Kumar, B. Oxidative Stress and Epigenetic Modifications in the Pathogenesis of Diabetic Retinopathy. Prog. Retin. Eye Res. 2015, 48, 40–61. [Google Scholar] [CrossRef]
- Reddy, M.A.; Zhang, E.; Natarajan, R. Epigenetic Mechanisms in Diabetic Complications and Metabolic Memory. Diabetologia 2015, 58, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Santos, J.M.; Mishra, M. Epigenetic Modifications and Diabetic Retinopathy. BioMed Res. Int. 2013, 2013, 635284. [Google Scholar] [CrossRef] [PubMed]
- Wisman, G.B.A.; Wojdacz, T.K.; Altucci, L.; Rots, M.G.; DeMeo, D.L.; Snieder, H. Clinical Promise and Applications of Epigenetic Biomarkers. Clin. Epigenetics 2024, 16, 192. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lo, A.C.Y. Diabetic Retinopathy: Pathophysiology and Treatments. Int. J. Mol. Sci. 2018, 19, 1816. [Google Scholar] [CrossRef]
- Davidson, B.A.; Miranda, A.X.; Reed, S.C.; Bergman, R.E.; Kemp, J.D.J.; Reddy, A.P.; Pantone, M.V.; Fox, E.K.; Dorand, R.D.; Hurley, P.J.; et al. An in Vitro CRISPR Screen of Cell-Free DNA Identifies Apoptosis as the Primary Mediator of Cell-Free DNA Release. Commun. Biol. 2024, 7, 1–15. [Google Scholar] [CrossRef]
- Medina, J.E.; Dracopoli, N.C.; Bach, P.B.; Lau, A.; Scharpf, R.B.; Meijer, G.A.; Andersen, C.L.; Velculescu, V.E. Cell-Free DNA Approaches for Cancer Early Detection and Interception. J. Immunother. Cancer 2023, 11, e006013. [Google Scholar] [CrossRef]
- Yu, D.; Tong, Y.; Guo, X.; Feng, L.; Jiang, Z.; Ying, S.; Jia, J.; Fang, Y.; Yu, M.; Xia, H.; et al. Diagnostic Value of Concentration of Circulating Cell-Free DNA in Breast Cancer: A Meta-Analysis. Front. Oncol. 2019, 9, 95. [Google Scholar] [CrossRef]
- Gianni, C.; Palleschi, M.; Merloni, F.; Di Menna, G.; Sirico, M.; Sarti, S.; Virga, A.; Ulivi, P.; Cecconetto, L.; Mariotti, M.; et al. Cell-Free DNA Fragmentomics: A Promising Biomarker for Diagnosis, Prognosis and Prediction of Response in Breast Cancer. Int. J. Mol. Sci. 2022, 23, 14197. [Google Scholar] [CrossRef]
- Uehiro, N.; Sato, F.; Pu, F.; Tanaka, S.; Kawashima, M.; Kawaguchi, K.; Sugimoto, M.; Saji, S.; Toi, M. Circulating Cell-Free DNA-Based Epigenetic Assay Can Detect Early Breast Cancer. Breast Cancer Res. 2016, 18, 129. [Google Scholar] [CrossRef]
- Lianidou, E. Detection and Relevance of Epigenetic Markers on ctDNA: Recent Advances and Future Outlook. Mol. Oncol. 2021, 15, 1683–1700. [Google Scholar] [CrossRef]
- Han, L.; Chen, C.; Lu, X.; Song, Y.; Zhang, Z.; Zeng, C.; Chiu, R.; Li, L.; Xu, M.; He, C.; et al. Alterations of 5-Hydroxymethylcytosines in Circulating Cell-Free DNA Reflect Retinopathy in Type 2 Diabetes. Genomics 2021, 113, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The Pathobiology of Diabetic Complications: A Unifying Mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Oxidative Stress and Diabetic Complications | Circulation Research. Available online: https://www.ahajournals.org/doi/10.1161/CIRCRESAHA.110.223545 (accessed on 20 March 2025).
- Kang, Q.; Dai, H.; Jiang, S.; Yu, L. Advanced Glycation End Products in Diabetic Retinopathy and Phytochemical Therapy. Front. Nutr. 2022, 9, 1037186. [Google Scholar] [CrossRef]
- Goh, S.-Y.; Cooper, M.E. The Role of Advanced Glycation End Products in Progression and Complications of Diabetes. J. Clin. Endocrinol. Metab. 2008, 93, 1143–1152. [Google Scholar] [CrossRef]
- Naruse, K.; Nakamura, J.; Hamada, Y.; Nakayama, M.; Chaya, S.; Komori, T.; Kato, K.; Kasuya, Y.; Miwa, K.; Naruse, K.; et al. Aldose Reductase Inhibition Prevents Glucose-Induced Apoptosis in Cultured Bovine Retinal Microvascular Pericytes. Exp. Eye Res. 2000, 71, 309–315. [Google Scholar] [CrossRef]
- Romeo, G.; Liu, W.-H.; Asnaghi, V.; Kern, T.S.; Lorenzi, M. Activation of Nuclear Factor-κB Induced by Diabetes and High Glucose Regulates a Proapoptotic Program in Retinal Pericytes. Diabetes 2002, 51, 2241–2248. [Google Scholar] [CrossRef]
- Ejaz, S.; Chekarova, I.; Ejaz, A.; Sohail, A.; Lim, C.W. Importance of Pericytes and Mechanisms of Pericyte Loss during Diabetic Retinopathy. Diabetes Obes. Metab. 2008, 10, 53–63. [Google Scholar] [CrossRef]
- Yu, D.-Y.; Cringle, S.J.; Balaratnasingam, C.; Morgan, W.H.; Yu, P.K.; Su, E.-N. Retinal Ganglion Cells: Energetics, Compartmentation, Axonal Transport, Cytoskeletons and Vulnerability. Prog. Retin. Eye Res. 2013, 36, 217–246. [Google Scholar] [CrossRef]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural Apoptosis in the Retina during Experimental and Human Diabetes. Early Onset and Effect of Insulin. J. Clin. Invest. 1998, 102, 783–791. [Google Scholar] [CrossRef]
- Podestà, F.; Romeo, G.; Liu, W.-H.; Krajewski, S.; Reed, J.C.; Gerhardinger, C.; Lorenzi, M. Bax Is Increased in the Retina of Diabetic Subjects and Is Associated with Pericyte Apoptosis in Vivo and in Vitro. Am. J. Pathol. 2000, 156, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- El-Asrar, A.M.A.; Dralands, L.; Missotten, L.; Al-Jadaan, I.A.; Geboes, K. Expression of Apoptosis Markers in the Retinas of Human Subjects with Diabetes. Invest. Ophthalmol. Vis. Sci. 2004, 45, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; and Koppolu, P. Diabetes-Induced Activation of Caspase-3 in Retina: Effect of Antioxidant Therapy. Free Radic. Res. 2002, 36, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Tamkovich, S.; Cherepanova, A.; Kolesnikova, E.; Rykova, E.; Pyshnyi, D.; Vlassov, V.; Laktionov, P. Circulating DNA and DNase Activity in Human Blood. N. Y. Acad. Sci. 2006, 1075, 191–196. [Google Scholar] [CrossRef]
- Sender, R.; Noor, E.; Milo, R.; Dor, Y. What Fraction of Cellular DNA Turnover Becomes cfDNA? eLife 2024, 12, RP89321. [Google Scholar] [CrossRef]
- Yao, W.; Mei, C.; Nan, X.; Hui, L. Evaluation and Comparison of in Vitro Degradation Kinetics of DNA in Serum, Urine and Saliva: A Qualitative Study. Gene 2016, 590, 142–148. [Google Scholar] [CrossRef]
- Diehl, F.; Schmidt, K.; Choti, M.A.; Romans, K.; Goodman, S.; Li, M.; Thornton, K.; Agrawal, N.; Sokoll, L.; Szabo, S.A.; et al. Circulating Mutant DNA to Assess Tumor Dynamics. Nat. Med. 2008, 14, 985–990. [Google Scholar] [CrossRef]
- Barber, A.J.; Gardner, T.W.; Abcouwer, S.F. The Significance of Vascular and Neural Apoptosis to the Pathology of Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 2011, 52, 1156–1163. [Google Scholar] [CrossRef]
- Li, X.; Hu, R.; Luo, T.; Peng, C.; Gong, L.; Hu, J.; Yang, S.; Li, Q. Serum Cell-Free DNA and Progression of Diabetic Kidney Disease: A Prospective Study. BMJ Open Diabetes Res. Care 2020, 8, e001078. [Google Scholar] [CrossRef]
- Tarhouny, S.A.E.; Hadhoud, K.M.; Ebrahem, M.M.; Azizi, N.M.A. Assessment of Cell-Free DNA with Microvascular Complication of Type II Diabetes Mellitus, Using PCR and Elisa. Nucleosides Nucleotides Nucleic Acids 2010, 29, 228–236. [Google Scholar] [CrossRef]
- Lyssenko, V.; Jonsson, A.; Almgren, P.; Pulizzi, N.; Isomaa, B.; Tuomi, T.; Berglund, G.; Altshuler, D.; Nilsson, P.; Groop, L. Clinical Risk Factors, DNA Variants, and the Development of Type 2 Diabetes. N. Engl. J. Med. 2008, 359, 2220–2232. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.J.; Pinho, M.G.M.; Abreu, T.C.; den Braver, N.R.; Lam, T.M.; Huss, A.; Vlaanderen, J.; Sonnenschein, T.; Siddiqui, N.Z.; Yuan, Z.; et al. Environmental Risk Factors of Type 2 Diabetes—An Exposome Approach. Diabetologia 2022, 65, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Florez, J.C.; Loos, R.J.F.; Froguel, P. Dissection of Type 2 Diabetes: A Genetic Perspective. Lancet Diabetes Endocrinol. 2025, 13, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-D.; Zhang, C.-Y.; Zhang, J.-T.; Gu, L.-M.; Xu, G.-T.; Zhang, J.-F. Epigenetic Modifications and Metabolic Memory in Diabetic Retinopathy: Beyond the Surface. Neural Regen. Res. 2022, 18, 1441–1449. [Google Scholar] [CrossRef]
- Singal, R.; Ginder, G.D. DNA Methylation. Blood 1999, 93, 4059–4070. [Google Scholar] [CrossRef]
- González, P.; Lozano, P.; Ros, G.; Solano, F. Hyperglycemia and Oxidative Stress: An Integral, Updated and Critical Overview of Their Metabolic Interconnections. Int. J. Mol. Sci. 2023, 24, 9352. [Google Scholar] [CrossRef]
- Ahmad, S.; Akhter, F.; Ahmad, K.; Khan, S. Editorial: Impact of Hyperglycemia Induced Oxidative Stress in Genetics and Epigenetics of Metabolic Diseases. Front. Genet. 2023, 14, 1123665. [Google Scholar] [CrossRef]
- Maghbooli, Z.; Hossein-nezhad, A.; Larijani, B.; Amini, M.; Keshtkar, A. Global DNA Methylation as a Possible Biomarker for Diabetic Retinopathy. Diabetes Metab. Res. Rev. 2015, 31, 183–189. [Google Scholar] [CrossRef]
- He, S.; Yiu, G.; Zhou, P.; Chen, D.F. Chapter 33—Epigenetic Mechanisms of Retinal Disease. In Retina, 5th ed.; Ryan, S.J., Sadda, S.R., Hinton, D.R., Schachat, A.P., Sadda, S.R., Wilkinson, C.P., Wiedemann, P., Schachat, A.P., Eds.; W.B. Saunders: London, UK, 2013; pp. 642–651. ISBN 978-1-4557-0737-9. [Google Scholar]
- Milluzzo, A.; Maugeri, A.; Barchitta, M.; Sciacca, L.; Agodi, A. Epigenetic Mechanisms in Type 2 Diabetes Retinopathy: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 10502. [Google Scholar] [CrossRef]
- Cai, C.; Meng, C.; He, S.; Gu, C.; Lhamo, T.; Draga, D.; Luo, D.; Qiu, Q. DNA Methylation in Diabetic Retinopathy: Pathogenetic Role and Potential Therapeutic Targets. Cell Biosci. 2022, 12, 186. [Google Scholar] [CrossRef]
- Tewari, S.; Zhong, Q.; Santos, J.M.; Kowluru, R.A. Mitochondria DNA Replication and DNA Methylation in the Metabolic Memory Associated with Continued Progression of Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 2012, 53, 4881–4888. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Nunes, M.K.; Silva, A.S.; de Queiroga Evangelista, I.W.; Filho, J.M.; Gomes, C.N.A.P.; do Nascimento, R.A.F.; Luna, R.C.P.; de Carvalho Costa, M.J.; de Oliveira, N.F.P.; Persuhn, D.C. Hypermethylation in the Promoter of the MTHFR Gene Is Associated with Diabetic Complications and Biochemical Indicators. Diabetol. Metab. Syndr. 2017, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Santana Bezerra, H.; Severo de Assis, C.; dos Santos Nunes, M.K.; Wanderley de Queiroga Evangelista, I.; Modesto Filho, J.; Alves Pegado Gomes, C.N.; Ferreira do Nascimento, R.A.; Pordeus Luna, R.C.; de Carvalho Costa, M.J.; de Oliveira, N.F.P.; et al. The MTHFR Promoter Hypermethylation Pattern Associated with the A1298C Polymorphism Influences Lipid Parameters and Glycemic Control in Diabetic Patients. Diabetol. Metab. Syndr. 2019, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, X.; Lu, X.; You, L.; Song, Y.; Luo, Z.; Zhang, J.; Nie, J.; Zheng, W.; Xu, D.; et al. 5-Hydroxymethylcytosine Signatures in Circulating Cell-Free DNA as Diagnostic Biomarkers for Human Cancers. Cell Res. 2017, 27, 1243–1257. [Google Scholar] [CrossRef]
- Dos Santos Nunes, M.K.; Silva, A.S.; Wanderley de Queiroga Evangelista, I.; Modesto Filho, J.; Alves Pegado Gomes, C.N.; Ferreira do Nascimento, R.A.; Pordeus Luna, R.C.; de Carvalho Costa, M.J.; Paulo de Oliveira, N.F.; Camati Persuhn, D. Analysis of the DNA Methylation Profiles of miR-9-3, miR-34a, and miR-137 Promoters in Patients with Diabetic Retinopathy and Nephropathy. J. Diabetes Complicat. 2018, 32, 593–601. [Google Scholar] [CrossRef]
- Duraisamy, A.J.; Radhakrishnan, R.; Seyoum, B.; Abrams, G.W.; Kowluru, R.A. Epigenetic Modifications in Peripheral Blood as Potential Noninvasive Biomarker of Diabetic Retinopathy. Transl. Vis. Sci. Technol. 2019, 8, 43. [Google Scholar] [CrossRef]
- Branco, M.R.; Ficz, G.; Reik, W. Uncovering the Role of 5-Hydroxymethylcytosine in the Epigenome. Nat. Rev. Genet. 2012, 13, 7–13. [Google Scholar] [CrossRef]
- Zhong, Q.; Kowluru, R.A. Role of Histone Acetylation in the Development of Diabetic Retinopathy and the Metabolic Memory Phenomenon. J. Cell. Biochem. 2010, 110, 1306–1313. [Google Scholar] [CrossRef]
- Zhong, Q.; Kowluru, R.A. Epigenetic Modification of Sod2 in the Development of Diabetic Retinopathy and in the Metabolic Memory: Role of Histone Methylation. Invest. Ophthalmol. Vis. Sci. 2013, 54, 244–250. [Google Scholar] [CrossRef]
- Yang, S.; Guo, X.; Cheng, W.; Seth, I.; Bulloch, G.; Chen, Y.; Shang, X.; Zhu, Z.; Huang, W.; Wang, W. Genome-Wide DNA Methylation Analysis of Extreme Phenotypes in the Identification of Novel Epigenetic Modifications in Diabetic Retinopathy. Clin. Epigenetics 2022, 14, 137. [Google Scholar] [CrossRef]
- Li, W.; Yanoff, M.; Liu, X.; Ye, X. Retinal Capillary Pericyte Apoptosis in Early Human Diabetic Retinopathy. Chin. Med. J. 1997, 110, 659–663. [Google Scholar] [PubMed]
- DNA Packaging: Nucleosomes and Chromatin | Learn Science at Scitable. Available online: http://www.nature.com/scitable/topicpage/dna-packaging-nucleosomes-and-chromatin-310 (accessed on 20 March 2025).
- Jahr, S.; Hentze, H.; Englisch, S.; Hardt, D.; Fackelmayer, F.O.; Hesch, R.-D.; Knippers, R. DNA Fragments in the Blood Plasma of Cancer Patients: Quantitations and Evidence for Their Origin from Apoptotic and Necrotic Cells. Cancer Res. 2001, 61, 1659–1665. [Google Scholar] [PubMed]
- Snyder, M.W.; Kircher, M.; Hill, A.J.; Daza, R.M.; Shendure, J. Cell-Free DNA Comprises an In Vivo Nucleosome Footprint That Informs Its Tissues-Of-Origin. Cell 2016, 164, 57–68. [Google Scholar] [CrossRef]
- Bachman, M.; Uribe-Lewis, S.; Yang, X.; Williams, M.; Murrell, A.; Balasubramanian, S. 5-Hydroxymethylcytosine Is a Predominantly Stable DNA Modification. Nat. Chem. 2014, 6, 1049–1055. [Google Scholar] [CrossRef]
- Song, D.; Zhang, Z.; Zheng, J.; Zhang, W.; Cai, J. 5-Hydroxymethylcytosine Modifications in Circulating Cell-Free DNA: Frontiers of Cancer Detection, Monitoring, and Prognostic Evaluation. Biomark. Res. 2025, 13, 39. [Google Scholar] [CrossRef]
- Shinjo, K.; Hara, K.; Nagae, G.; Umeda, T.; Katsushima, K.; Suzuki, M.; Murofushi, Y.; Umezu, Y.; Takeuchi, I.; Takahashi, S.; et al. A Novel Sensitive Detection Method for DNA Methylation in Circulating Free DNA of Pancreatic Cancer. PLoS ONE 2020, 15, e0233782. [Google Scholar] [CrossRef]
- Merbs, S.L.; Khan, M.A.; Hackler, L.; Oliver, V.F.; Wan, J.; Qian, J.; Zack, D.J. Cell-Specific DNA Methylation Patterns of Retina-Specific Genes. PLoS ONE 2012, 7, e32602. [Google Scholar] [CrossRef]
- Wu, F.; Bard, J.E.; Kann, J.; Yergeau, D.; Sapkota, D.; Ge, Y.; Hu, Z.; Wang, J.; Liu, T.; Mu, X. Single Cell Transcriptomics Reveals Lineage Trajectory of Retinal Ganglion Cells in Wild-Type and Atoh7-Null Retinas. Nat. Commun. 2021, 12, 1465. [Google Scholar] [CrossRef]
- Ulz, P.; Perakis, S.; Zhou, Q.; Moser, T.; Belic, J.; Lazzeri, I.; Wölfler, A.; Zebisch, A.; Gerger, A.; Pristauz, G.; et al. Inference of Transcription Factor Binding from Cell-Free DNA Enables Tumor Subtype Prediction and Early Detection. Nat. Commun. 2019, 10, 4666. [Google Scholar] [CrossRef]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-Throughput Droplet Digital PCR System for Absolute Quantitation of DNA Copy Number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef]
- Diehl, F.; Schmidt, K.; Durkee, K.H.; Moore, K.J.; Goodman, S.N.; Shuber, A.P.; Kinzler, K.W.; Vogelstein, B. Analysis of Mutations in DNA Isolated from Plasma and Stool of Colorectal Cancer Patients. Gastroenterology 2008, 135, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Gradisteanu Pircalabioru, G.; Musat, M.; Elian, V.; Iliescu, C. Liquid Biopsy: A Game Changer for Type 2 Diabetes. Int. J. Mol. Sci. 2024, 25, 2661. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.Y.; Burgener, J.M.; Bratman, S.V.; De Carvalho, D.D. Preparation of cfMeDIP-Seq Libraries for Methylome Profiling of Plasma Cell-Free DNA. Nat. Protoc. 2019, 14, 2749–2780. [Google Scholar] [CrossRef] [PubMed]
- Koval, A.P.; Blagodatskikh, K.A.; Kushlinskii, N.E.; Shcherbo, D.S. The Detection of Cancer Epigenetic Traces in Cell-Free DNA. Front. Oncol. 2021, 11, 662094. [Google Scholar] [CrossRef]
- Forshew, T.; Murtaza, M.; Parkinson, C.; Gale, D.; Tsui, D.W.Y.; Kaper, F.; Dawson, S.-J.; Piskorz, A.M.; Jimenez-Linan, M.; Bentley, D.; et al. Noninvasive Identification and Monitoring of Cancer Mutations by Targeted Deep Sequencing of Plasma DNA. Sci. Transl. Med. 2012, 4, 136ra68. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.-Z.; Xiao, X.-F.; Chen, J.; Zhou, X.-Q.; Zhu, W.-Y.; Dai, Z.; Zou, X.-Y. Development of Techniques for DNA-Methylation Analysis. TrAC Trends Anal. Chem. 2015, 72, 114–122. [Google Scholar] [CrossRef]
- Matesva, M. Urinary Vegf and Cell-Free Dna as Non-Invasive Biomarkers for Diabetic Retinopathy Screening. Ph.D. Thesis, Yale Medicine Thesis Digital Library, New Haven, CT, USA, 2024. [Google Scholar]
- Gezer, U.; Bronkhorst, A.J.; Holdenrieder, S. The Utility of Repetitive Cell-Free DNA in Cancer Liquid Biopsies. Diagnostics 2022, 12, 1363. [Google Scholar] [CrossRef]
- Spindler, K.-L.G.; Appelt, A.L.; Pallisgaard, N.; Andersen, R.F.; Brandslund, I.; Jakobsen, A. Cell-Free DNA in Healthy Individuals, Noncancerous Disease and Strong Prognostic Value in Colorectal Cancer. Int. J. Cancer 2014, 135, 2984–2991. [Google Scholar] [CrossRef]
- Einbinder, Y.; Shnaider, A.; Ghanayem, K.; Basok, A.; Rogachev, B.; Lior, Y.; Haviv, Y.S.; Cohen-Hagai, K.; Nacasch, N.; Rozenberg, I.; et al. Elevated Circulating Cell-Free DNA in Hemodialysis-Treated Patients Is Associated with Increased Mortality. Am. J. Nephrol. 2021, 51, 852–860. [Google Scholar] [CrossRef]
- Rolfo, C.; Cardona, A.F.; Cristofanilli, M.; Paz-Ares, L.; Diaz Mochon, J.J.; Duran, I.; Raez, L.E.; Russo, A.; Lorente, J.A.; Malapelle, U.; et al. Challenges and Opportunities of cfDNA Analysis Implementation in Clinical Practice: Perspective of the International Society of Liquid Biopsy (ISLB). Crit. Rev. Oncol. Hematol. 2020, 151, 102978. [Google Scholar] [CrossRef]
- Chang, A.; Mzava, O.; Lenz, J.S.; Cheng, A.P.; Burnham, P.; Motley, S.T.; Bennett, C.; Connelly, J.T.; Dadhania, D.M.; Suthanthiran, M.; et al. Measurement Biases Distort Cell-Free DNA Fragmentation Profiles and Define the Sensitivity of Metagenomic Cell-Free DNA Sequencing Assays. Clin. Chem. 2022, 68, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Forestier, J.; Dusserre, E.; Wozny, A.-S.; Geiguer, F.; Merle, P.; Tissot, C.; Ferraro-Peyret, C.; Jones, F.S.; Edelstein, D.L.; et al. Cross-Platform Comparison for the Detection of RAS Mutations in cfDNA (ddPCR Biorad Detection Assay, BEAMing Assay, and NGS Strategy). Oncotarget 2018, 9, 21122–21131. [Google Scholar] [CrossRef] [PubMed]
- García-Foncillas, J.; Alba, E.; Aranda, E.; Díaz-Rubio, E.; López-López, R.; Tabernero, J.; Vivancos, A. Incorporating BEAMing Technology as a Liquid Biopsy into Clinical Practice for the Management of Colorectal Cancer Patients: An Expert Taskforce Review. Ann. Oncol. 2017, 28, 2943–2949. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, B.; Hrebien, S.; Beaney, M.; Fribbens, C.; Garcia-Murillas, I.; Jiang, J.; Li, Y.; Huang Bartlett, C.; André, F.; Loibl, S.; et al. Comparison of BEAMing and Droplet Digital PCR for Circulating Tumor DNA Analysis. Clin. Chem. 2019, 65, 1405–1413. [Google Scholar] [CrossRef]
- Lau, E.; McCoy, P.; Reeves, F.; Chow, K.; Clarkson, M.; Kwan, E.M.; Packwood, K.; Northen, H.; He, M.; Kingsbury, Z.; et al. Detection of ctDNA in Plasma of Patients with Clinically Localised Prostate Cancer Is Associated with Rapid Disease Progression. Genome Med. 2020, 12, 72. [Google Scholar] [CrossRef]
- Gale, D.; Plagnol, V.; Lawson, A.; Pugh, M.; Smalley, S.; Howarth, K.; Madi, M.; Durham, B.; Kumanduri, V.; Lo, K.; et al. Abstract 3639: Analytical Performance and Validation of an Enhanced TAm-Seq Circulating Tumor DNA Sequencing Assay. Cancer Res. 2016, 76, 3639. [Google Scholar] [CrossRef]
- Lawson, A.R.; Plagnol, V.; Fahem, A.; Forshew, T.; Brenton, J.D.; Gale, D.; Rosenfeld, N. Abstract 2412: Assessment of Clinical Applications of Circulating Tumor DNA Using an Enhanced TAm-Seq Platform. Cancer Res. 2015, 75, 2412. [Google Scholar] [CrossRef]
- Henriksen, T.V.; Reinert, T.; Rasmussen, M.H.; Demuth, C.; Løve, U.S.; Madsen, A.H.; Gotschalck, K.A.; Iversen, L.H.; Andersen, C.L. Comparing Single-Target and Multitarget Approaches for Postoperative Circulating Tumour DNA Detection in Stage II–III Colorectal Cancer Patients. Mol. Oncol. 2022, 16, 3654–3665. [Google Scholar] [CrossRef]
- Song, P.; Wu, L.R.; Yan, Y.H.; Zhang, J.X.; Chu, T.; Kwong, L.N.; Patel, A.A.; Zhang, D.Y. Limitations and Opportunities of Technologies for the Analysis of Cell-Free DNA in Cancer Diagnostics. Nat. Biomed. Eng. 2022, 6, 232–245. [Google Scholar] [CrossRef]
- Ivanov, M.; Baranova, A.; Butler, T.; Spellman, P.; Mileyko, V. Non-Random Fragmentation Patterns in Circulating Cell-Free DNA Reflect Epigenetic Regulation. BMC Genom. 2015, 16, S1. [Google Scholar] [CrossRef]
- Stewart, C.M.; Kothari, P.D.; Mouliere, F.; Mair, R.; Somnay, S.; Benayed, R.; Zehir, A.; Weigelt, B.; Dawson, S.-J.; Arcila, M.E.; et al. The Value of Cell-Free DNA for Molecular Pathology. J. Pathol. 2018, 244, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Yuwono, N.L.; Warton, K.; Ford, C.E. The Influence of Biological and Lifestyle Factors on Circulating Cell-Free DNA in Blood Plasma. eLife 2021, 10, e69679. [Google Scholar] [CrossRef] [PubMed]
- Adkins, R.M.; Krushkal, J.; Tylavsky, F.A.; Thomas, F. Racial Differences in Gene-Specific DNA Methylation Levels Are Present at Birth. Birt. Defects Res. A. Clin. Mol. Teratol. 2011, 91, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.Y.; Hahn, S.; Kiefer, V.; Holzgreve, W. Is the Quantity of Circulatory Cell-Free DNA in Human Plasma and Serum Samples Associated with Gender, Age and Frequency of Blood Donations? Ann. Hematol. 2007, 86, 139–143. [Google Scholar] [CrossRef]
- Ørntoft, M.-B.W.; Jensen, S.Ø.; Øgaard, N.; Henriksen, T.V.; Ferm, L.; Christensen, I.J.; Reinert, T.; Larsen, O.H.; Nielsen, H.J.; Andersen, C.L. Age-Stratified Reference Intervals Unlock the Clinical Potential of Circulating Cell-Free DNA as a Biomarker of Poor Outcome for Healthy Individuals and Patients with Colorectal Cancer. Int. J. Cancer 2021, 148, 1665–1675. [Google Scholar] [CrossRef]
- Kolesar, J.; Peh, S.; Thomas, L.; Baburaj, G.; Mukherjee, N.; Kantamneni, R.; Lewis, S.; Pai, A.; Udupa, K.S.; Kumar AN, N.; et al. Integration of Liquid Biopsy and Pharmacogenomics for Precision Therapy of EGFR Mutant and Resistant Lung Cancers. Mol. Cancer 2022, 21, 61. [Google Scholar] [CrossRef]
- Cai, K.; Liu, Y.-P.; Wang, D. Prevalence of Diabetic Retinopathy in Patients with Newly Diagnosed Type 2 Diabetes: A Systematic Review and Meta-Analysis. Diabetes Metab. Res. Rev. 2023, 39, e3586. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Yim, M.M.; Jin, Y.X.; Tao, B.K.; Xie, J.S.; Balas, M.; Khan, H.; Lam, W.-C.; Yan, P.; Navajas, E.V. Circulating Cell-Free DNA as an Epigenetic Biomarker for Early Diabetic Retinopathy: A Narrative Review. Diagnostics 2025, 15, 1161. https://doi.org/10.3390/diagnostics15091161
Li B, Yim MM, Jin YX, Tao BK, Xie JS, Balas M, Khan H, Lam W-C, Yan P, Navajas EV. Circulating Cell-Free DNA as an Epigenetic Biomarker for Early Diabetic Retinopathy: A Narrative Review. Diagnostics. 2025; 15(9):1161. https://doi.org/10.3390/diagnostics15091161
Chicago/Turabian StyleLi, Boaz, Megan M. Yim, Yu Xuan Jin, Brendan K. Tao, Jim S. Xie, Michael Balas, Haaris Khan, Wai-Ching Lam, Peng Yan, and Eduardo V. Navajas. 2025. "Circulating Cell-Free DNA as an Epigenetic Biomarker for Early Diabetic Retinopathy: A Narrative Review" Diagnostics 15, no. 9: 1161. https://doi.org/10.3390/diagnostics15091161
APA StyleLi, B., Yim, M. M., Jin, Y. X., Tao, B. K., Xie, J. S., Balas, M., Khan, H., Lam, W.-C., Yan, P., & Navajas, E. V. (2025). Circulating Cell-Free DNA as an Epigenetic Biomarker for Early Diabetic Retinopathy: A Narrative Review. Diagnostics, 15(9), 1161. https://doi.org/10.3390/diagnostics15091161