
Recent Advances and Potential Future Applications of MALDI-TOF Mass Spectrometry for Identification of Helminths


Abstract
1. Introduction
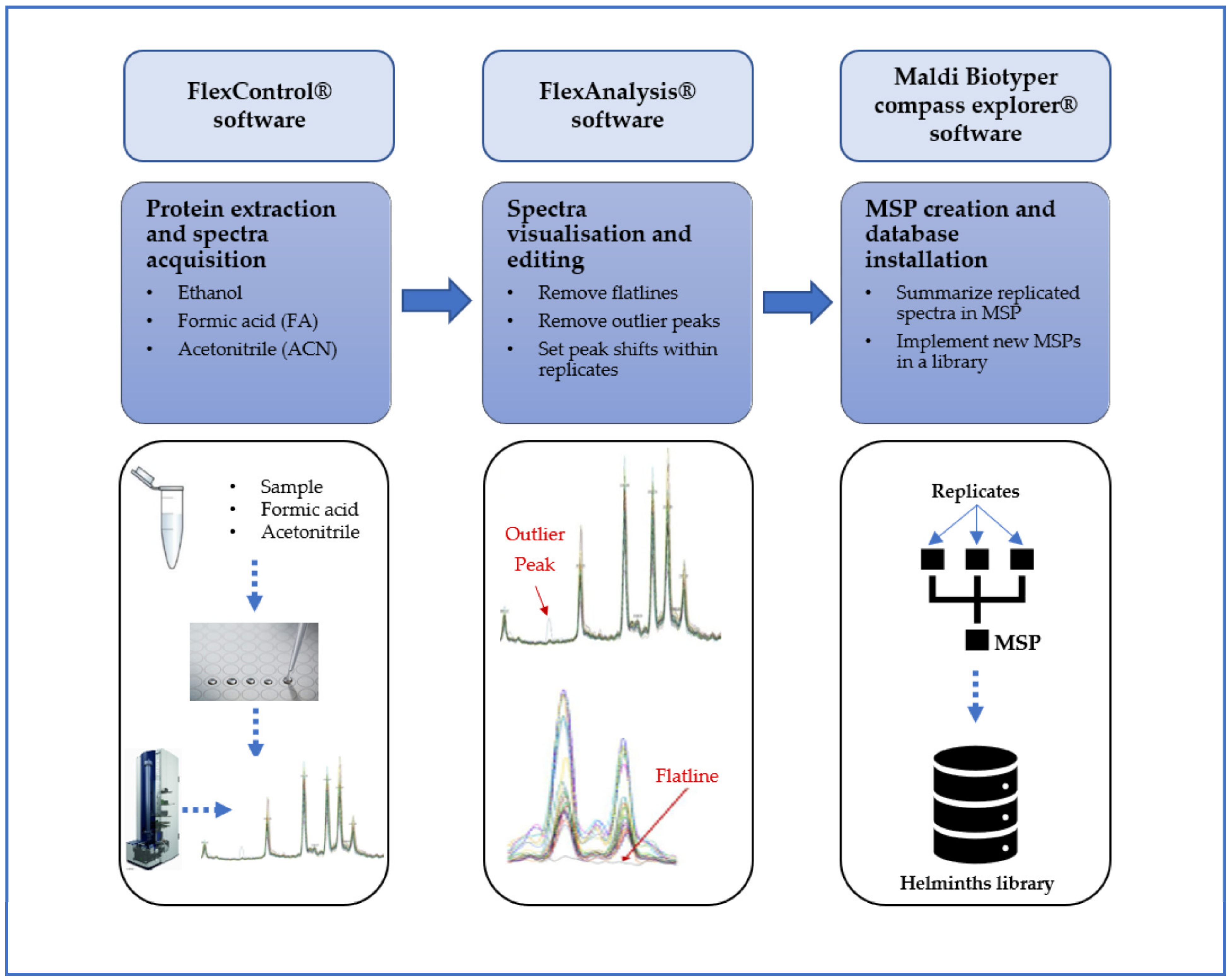
2. Materials and Methods
3. Specific Application of MALDI-TOF MS for Different Helminths
3.1. Cestodes
3.2. Nematodes
3.3. Trematodes
4. Effect of Storage Media and Duration
5. Challenges and Future Research Needs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castro, G.A. Helminths: Structure, Classification, Growth, and Development. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 978-0-9631172-1-2. [Google Scholar]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth Infections: The Great Neglected Tropical Diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Research Priorities for Helminth Infections: Technical Report of the TDR Disease Reference Group on Helminth Infections; World Health Organization: Geneva, Switzerland, 2012; ISBN 978-92-4-120972-4.
- Irisarri-Gutiérrez, M.J.; Muñoz-Antolí, C.; Acosta, L.; Parker, L.A.; Toledo, R.; Bornay-Llinares, F.J.; Esteban, J.G. Hookworm-like Eggs in Children’s Faecal Samples from a Rural Area of Rwanda. Afr. Health Sci. 2016, 16, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Ide, L. To Differentiate Taenia Eggs. J. Clin. Microbiol. 2012, 50, 2836; author reply 2837. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Utzinger, J.; Becker, S.L.; van Lieshout, L.; van Dam, G.J.; Knopp, S. New Diagnostic Tools in Schistosomiasis. Clin. Microbiol. Infect. 2015, 21, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Knopp, S.; Becker, S.L.; Ingram, K.J.; Keiser, J.; Utzinger, J. Diagnosis and Treatment of Schistosomiasis in Children in the Era of Intensified Control. Expert Rev. Anti-Infect. Ther. 2013, 11, 1237–1258. [Google Scholar] [CrossRef]
- Gordon, C.A.; Gray, D.J.; Gobert, G.N.; McManus, D.P. DNA Amplification Approaches for the Diagnosis of Key Parasitic Helminth Infections of Humans. Mol. Cell. Probes 2011, 25, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF Mass Spectrometry in Clinical Diagnostic Microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef]
- Welker, M.; Van Belkum, A.; Girard, V.; Charrier, J.-P.; Pincus, D. An Update on the Routine Application of MALDI-TOF MS in Clinical Microbiology. Expert Rev. Proteom. 2019, 16, 695–710. [Google Scholar] [CrossRef]
- Oliva, E.; Arosio, M.; Mazzola, E.; Mascheroni, M.; Cerro, A.; Cuntró, M.; Vailati, F.; Cavallini, M.; Farina, C. Rapid Identification of Non-Tuberculous Mycobacteria with MALDI-TOF Mass Spectrometry. Infez. Med. 2021, 29, 79–84. [Google Scholar]
- Fernández-Esgueva, M.; Fernández-Simon, R.; Monforte-Cirac, M.L.; López-Calleja, A.I.; Fortuño, B.; Viñuelas-Bayon, J. Use of MALDI-TOF MS (Bruker Daltonics) for Identification of Mycobacterium Species Isolated Directly from Liquid Medium. Enferm. Infecc. Microbiol. Clín. (Engl. Ed.) 2021, 39, 241–243. [Google Scholar] [CrossRef]
- Robert, M.-G.; Cornet, M.; Hennebique, A.; Rasamoelina, T.; Caspar, Y.; Pondérand, L.; Bidart, M.; Durand, H.; Jacquet, M.; Garnaud, C.; et al. MALDI-TOF MS in a Medical Mycology Laboratory: On Stage and Backstage. Microorganisms 2021, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.-S.; Kim, Y.H. Rapid and Robust MALDI-TOF MS Techniques for Microbial Identification: A Brief Overview of Their Diverse Applications. J. Microbiol. 2018, 56, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Solntceva, V.; Kostrzewa, M.; Larrouy-Maumus, G. Detection of Species-Specific Lipids by Routine MALDI TOF Mass Spectrometry to Unlock the Challenges of Microbial Identification and Antimicrobial Susceptibility Testing. Front. Cell. Infect. Microbiol. 2020, 10, 621452. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, X.; Broda, A.; Drobniewski, F.; Larrouy-Maumus, G. Performance of Lipid Fingerprint-Based MALDI-ToF for the Diagnosis of Mycobacterial Infections. Clin. Microbiol. Infect. 2021, 27, 912.e1–912.e5. [Google Scholar] [CrossRef]
- Larrouy-Maumus, G.; Puzo, G. Mycobacterial Envelope Lipids Fingerprint from Direct MALDI-TOF MS Analysis of Intact Bacilli. Tuberculosis 2015, 95, 75–85. [Google Scholar] [CrossRef]
- Feucherolles, M.; Nennig, M.; Becker, S.L.; Martiny, D.; Losch, S.; Penny, C.; Cauchie, H.-M.; Ragimbeau, C. Combination of MALDI-TOF Mass Spectrometry and Machine Learning for Rapid Antimicrobial Resistance Screening: The Case of Campylobacter spp. Front. Microbiol. 2021, 12, 804484. [Google Scholar] [CrossRef]
- Dingle, T.C.; Butler-Wu, S.M. Maldi-Tof Mass Spectrometry for Microorganism Identification. Clin. Lab. Med. 2013, 33, 589–609. [Google Scholar] [CrossRef]
- Feucherolles, M.; Poppert, S.; Utzinger, J.; Becker, S.L. MALDI-TOF Mass Spectrometry as a Diagnostic Tool in Human and Veterinary Helminthology: A Systematic Review. Parasites Vectors 2019, 12, 245. [Google Scholar] [CrossRef]
- Díaz-Zaragoza, M.; Jiménez, L.; Hernández, M.; Hernández-Ávila, R.; Navarro, L.; Ochoa-Sánchez, A.; Encarnación-Guevara, S.; Ostoa-Saloma, P.; Landa, A. Protein Expression Profile of Taenia crassiceps Cysticerci Related to Th1- and Th2-Type Responses in the Mouse Cysticercosis Model. Acta Trop. 2020, 212, 105696. [Google Scholar] [CrossRef]
- Wendel, T.P.; Feucherolles, M.; Rehner, J.; Poppert, S.; Utzinger, J.; Becker, S.L.; Sy, I. Evaluating Different Storage Media for Identification of Taenia saginata Proglottids Using MALDI-TOF Mass Spectrometry. Microorganisms 2021, 9, 2006. [Google Scholar] [CrossRef]
- Bhat, K.A.; Mir, R.A.; Farooq, A.; Manzoor, M.; Hami, A.; Allie, K.A.; Wani, S.M.; Khan, M.N.; Sayyed, R.Z.; Poczai, P.; et al. Advances in Nematode Identification: A Journey from Fundamentals to Evolutionary Aspects. Diversity 2022, 14, 536. [Google Scholar] [CrossRef]
- Bogale, M.; Baniya, A.; DiGennaro, P. Nematode Identification Techniques and Recent Advances. Plants 2020, 9, 1260. [Google Scholar] [CrossRef] [PubMed]
- Bredtmann, C.M.; Krücken, J.; Murugaiyan, J.; Balard, A.; Hofer, H.; Kuzmina, T.A.; von Samson-Himmelstjerna, G. Concurrent Proteomic Fingerprinting and Molecular Analysis of Cyathostomins. Proteomics 2019, 19, e1800290. [Google Scholar] [CrossRef] [PubMed]
- Nagorny, S.; Aleshukina, A.; Aleshukina, I.; Denisenko, V.; Ermakova, L.; Pshenichnaya, N. Application of Mathematical Models for MALDI-TOF MS on the Example of Dirofilariasis. Int. J. Infect. Dis. 2022, 116, S95. [Google Scholar] [CrossRef]
- Rivero, J.; Zurita, A.; Cutillas, C.; Callejón, R. The Use of MALDI-TOF MS as a Diagnostic Tool for Adult Trichuris Species. Front. Vet. Sci. 2022, 9, 867919. [Google Scholar] [CrossRef]
- Karadjian, G.; Bilska-Zając, E.; Bahn, P.; Py, J.-S.; Johne, A.; Gassilloud, B.; Różycki, M.; Cencek, T.; Mayer-Scholl, A.; Vallée, I. Species Identification of Trichinella Originated from Various Host and Different Geographical Location by MALDI-TOF. Exp. Parasitol. 2020, 213, 107890. [Google Scholar] [CrossRef]
- Marzano, V.; Pane, S.; Foglietta, G.; Levi Mortera, S.; Vernocchi, P.; Onetti Muda, A.; Putignani, L. Mass Spectrometry Based-Proteomic Analysis of Anisakis spp.: A Preliminary Study towards a New Diagnostic Tool. Genes 2020, 11, 693. [Google Scholar] [CrossRef]
- Khanmohammadi, M.; Falak, R.; Meamar, A.R.; Razmjou, E.; Mokhtarian, K.; Arshadi, M.; Shayanfar, N.; Akhlaghi, L. Application of Dirofilaria immitis Immunoreactive Proteins in Serodiagnosis. Parasite Immunol. 2019, 41, e12598. [Google Scholar] [CrossRef]
- Martini, F.; Eckmair, B.; Štefanić, S.; Jin, C.; Garg, M.; Yan, S.; Jiménez-Castells, C.; Hykollari, A.; Neupert, C.; Venco, L.; et al. Highly Modified and Immunoactive N-Glycans of the Canine Heartworm. Nat. Commun. 2019, 10, 75. [Google Scholar] [CrossRef]
- Wang, C.; Gao, W.; Yan, S.; Zhu, X.-Q.; Suo, X.; Liu, X.; Gupta, N.; Hu, M. N-Glycome and N-Glycoproteome of a Hematophagous Parasitic Nematode Haemonchus. Comput. Struct. Biotechnol. J. 2021, 19, 2486–2496. [Google Scholar] [CrossRef]
- Petralia, L.M.C.; van Diepen, A.; Lokker, L.A.; Nguyen, D.L.; Sartono, E.; Khatri, V.; Kalyanasundaram, R.; Taron, C.H.; Foster, J.M.; Hokke, C.H. Mass Spectrometric and Glycan Microarray-Based Characterization of the Filarial Nematode Brugia malayi Glycome Reveals Anionic and Zwitterionic Glycan Antigens. Mol. Cell. Proteom. 2022, 21, 100201. [Google Scholar] [CrossRef] [PubMed]
- Huguenin, A.; Depaquit, J.; Villena, I.; Ferté, H. MALDI-TOF Mass Spectrometry: A New Tool for Rapid Identification of Cercariae (Trematoda, Digenea). Parasite 2019, 26, 11. [Google Scholar] [CrossRef] [PubMed]
- Kästner, C.; Bahn, P.; Schönfelder, R.; Ozoliņa, Z.; Alksne, L.; Richter, M.H.; Deksne, G.; Mayer-Scholl, A.; Johne, A. Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Microorganisms 2021, 9, 1664. [Google Scholar] [CrossRef] [PubMed]
- Huguenin, A.; Kincaid-Smith, J.; Depaquit, J.; Boissier, J.; Ferté, H. MALDI-TOF: A New Tool for the Identification of Schistosoma Cercariae and Detection of Hybrids. bioRxiv 2022. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, Y.; Huang, Y.; Sun, F.; Tian, H.; Hu, N.; Shi, L.; Hua, H. Identification of Newly Developed Advanced Schistosomiasis with MALDI-TOF Mass Spectrometry and ClinProTools Analysis. Parasite 2019, 26, 33. [Google Scholar] [CrossRef]
- Sy, I.; Margardt, L.; Ngbede, E.O.; Adah, M.I.; Yusuf, S.T.; Keiser, J.; Rehner, J.; Utzinger, J.; Poppert, S.; Becker, S.L. Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry. Microorganisms 2020, 9, 82. [Google Scholar] [CrossRef]
- Mohan, C. Study of Purification of Hemoglobin in Fasciola gigantica (Trematoda:Fasciolida). Biospectra 2020, 15, 6. [Google Scholar]
- Stracke, K.; Adisakwattana, P.; Phuanukoonnon, S.; Yoonuan, T.; Poodeepiyasawat, A.; Dekumyoy, P.; Chaisiri, K.; Roth Schulze, A.; Wilcox, S.; Karunajeewa, H.; et al. Effective Low-Cost Preservation of Human Stools in Field-Based Studies for Helminth and Microbiota Analysis. Int. J. Parasitol. 2021, 51, 741–748. [Google Scholar] [CrossRef]
- Hu, X.L.; Liu, G.; Wang, W.X.; Zhou, R.; Liu, S.Q.; Li, L.H.; Hu, D.F. Methods of Preservation and Flotation for the Detection of Nematode Eggs and Coccidian Oocysts in Faeces of the Forest Musk Deer. J. Helminthol. 2016, 90, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.A.H.; Chapman, S.N.; Lummaa, V.; Lynsdale, C.L. Testing Storage Methods of Faecal Samples for Subsequent Measurement of Helminth Egg Numbers in the Domestic Horse. Vet. Parasitol. 2016, 221, 130–133. [Google Scholar] [CrossRef]
- Barda, B.; Albonico, M.; Ianniello, D.; Ame, S.M.; Keiser, J.; Speich, B.; Rinaldi, L.; Cringoli, G.; Burioni, R.; Montresor, A.; et al. How Long Can Stool Samples Be Fixed for an Accurate Diagnosis of Soil-Transmitted Helminth Infection Using Mini-FLOTAC? PLoS Negl. Trop. Dis. 2015, 9, e0003698. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Scholl, A.; Murugaiyan, J.; Neumann, J.; Bahn, P.; Reckinger, S.; Nöckler, K. Rapid Identification of the Foodborne Pathogen Trichinella spp. by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. PLoS ONE 2016, 11, e0152062. [Google Scholar] [CrossRef] [PubMed]
- Singhal, N.; Kumar, M.; Virdi, J.S. MALDI-TOF MS in Clinical Parasitology: Applications, Constraints and Prospects. Parasitology 2016, 143, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, J.; Roesler, U. MALDI-TOF MS Profiling-Advances in Species Identification of Pests, Parasites, and Vectors. Front. Cell. Infect. Microbiol. 2017, 7, 184. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Koné, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF Identification of Ticks and Tick-Associated Bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef]
- Benyahia, H.; Ouarti, B.; Diarra, A.Z.; Boucheikhchoukh, M.; Meguini, M.N.; Behidji, M.; Benakhla, A.; Parola, P.; Almeras, L. Identification of Lice Stored in Alcohol Using MALDI-TOF MS. J. Med. Entomol 2021, 58, 1126–1133. [Google Scholar] [CrossRef]
- Zurita, A.; Djeghar, R.; Callejón, R.; Cutillas, C.; Parola, P.; Laroche, M. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry as a Useful Tool for the Rapid Identification of Wild Flea Vectors Preserved in Alcohol. Med. Vet. Entomol. 2019, 33, 185–194. [Google Scholar] [CrossRef]
- Ndiaye, E.H.I.; Diatta, G.; Diarra, A.Z.; Berenger, J.M.; Bassene, H.; Mediannikov, O.; Bouganali, C.; Sokhna, C.; Parola, P. Morphological, Molecular and MALDI-TOF MS Identification of Bedbugs and Associated Wolbachia Species in Rural Senegal. J. Med. Entomol. 2022, 59, 1019–1032. [Google Scholar] [CrossRef]
- Smout, M.J.; Kotze, A.C.; McCarthy, J.S.; Loukas, A. A Novel High Throughput Assay for Anthelmintic Drug Screening and Resistance Diagnosis by Real-Time Monitoring of Parasite Motility. PLoS Negl. Trop. Dis. 2010, 4, e885. [Google Scholar] [CrossRef]
- Shepherd, F.; Chylinski, C.; Hutchings, M.R.; Lima, J.; Davidson, R.; Kelly, R.; Macrae, A.; Salminen, J.-P.; Engström, M.T.; Maurer, V.; et al. Comparative Analysis of the Anthelmintic Efficacy of European Heather Extracts on Teladorsagia circumcincta and Trichostrongylus colubriformis Egg Hatching and Larval Motility. Parasites Vectors 2022, 15, 409. [Google Scholar] [CrossRef]


{kind=link}
| Pathogens | Analyzed Material | Correct Identification Rate in % [LSV] | References | ||||
|---|---|---|---|---|---|---|---|
| Serum of Infected Individuals | Specific Proteins | Larval Stage | Adult Helminth | ||||
| Cestodes | |||||||
| Taenia crassiceps | √ | NA | Diaz-Zaragoza et al., 2020 | ||||
| Taenia saginata | √ | 97.2–99.7% [≥2.5] | Wendel et al., 2021 | ||||
| Trematodes | |||||||
| Alaria alata | √ | 94.73% [≥2] | Kästner et al., 2021 | ||||
| Cotylurus sp. | √ | 65.78% [≥1.7] | Huguenin et al., 2019 | ||||
| Diplostomum pseudospathaceum | √ | ||||||
| Echinostoma revolutum | √ | ||||||
| Echinoparyphium sp. | √ | ||||||
| Fasciola spp.: | 98.7–100% [1.73–2.23] | Sy et al., 2020 | |||||
| F. gigantica | √ | ||||||
| F. hepatica | √ | ||||||
| Parastrigea sp. | √ | 65.78% [≥1.7] | Huguenin et al., 2019 | ||||
| Petasiger phalacrocoracis | √ | ||||||
| Plagiorchis sp. | √ | ||||||
| Parastrigea sp. | √ | ||||||
| Posthodiplostomum sp. | √ | ||||||
| Petasiger phalacrocoracis | √ | ||||||
| Schistosoma: | 42.7% [≥1.7] ≥97% [NA] | Huguenin et al., 2022 | |||||
| S. bovis | √ | ||||||
| S. haematobium | √ | ||||||
| S. mansoni | √ | ||||||
| S. rodhaini | √ | ||||||
| S. bovis × S. haematobium (hybrid) | √ | ||||||
| Schistosoma japonicum | √ | 92–100% [NA] | Huang et al., 2019 | ||||
| Trichobilharrzia anseri | √ | 65.78% [≥1.7] | Huguenin et al., 2019 | ||||
| Tylodelphys sp. | √ | ||||||
| Nematodes | |||||||
| Anisakis spp. | √ | NA | Marzano et al., 2020 | ||||
| Brugia malayi | √ | NA | Petralia et al., 2022 | ||||
| Cyatosthomins: | 89% [1.13–2.44] | Bredtmann et al., 2019 | |||||
| Cylicostephanus longibursatus | √ | ||||||
| Cylicostephanus minutus | √ | ||||||
| Dirofilaria spp.: | NA 70–100% [NA] | Khanmohammadi et al., 2019 Martini et al., 2019 Nagorny et al., 2019 | |||||
| D. immitis | √ | √ | √ | ||||
| D. immitis | √ | ||||||
| D. repens | √ | ||||||
| Haemonchus contortus | √ | √ | NA | Wang et al., 2021 | |||
| Trichinella spp.: | 83.3–100% 100% [≥2] | Karadjian et al., 2020 | |||||
| T. britovi | √ | ||||||
| T. nativa | √ | ||||||
| T. patagoniensis | √ | ||||||
| T. pseudospiralis | √ | ||||||
| T. spiralis | √ | ||||||
| Trichinella sp. T8 | √ | ||||||
| Trichinella sp. T9 | √ | ||||||
| Trichuris spp.: | 100% [1.84–2.36] | Rivero et al., 2022 | |||||
| Trichuris sp. | √ | ||||||
| T. ovis | √ | ||||||
| T. suis | √ | ||||||
| T. trichiura | √ | ||||||
| T. vulpis | √ | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sy, I.; Conrad, L.; Becker, S.L. Recent Advances and Potential Future Applications of MALDI-TOF Mass Spectrometry for Identification of Helminths. Diagnostics 2022, 12, 3035. https://doi.org/10.3390/diagnostics12123035
Sy I, Conrad L, Becker SL. Recent Advances and Potential Future Applications of MALDI-TOF Mass Spectrometry for Identification of Helminths. Diagnostics. 2022; 12(12):3035. https://doi.org/10.3390/diagnostics12123035
Chicago/Turabian StyleSy, Issa, Lucie Conrad, and Sören L. Becker. 2022. "Recent Advances and Potential Future Applications of MALDI-TOF Mass Spectrometry for Identification of Helminths" Diagnostics 12, no. 12: 3035. https://doi.org/10.3390/diagnostics12123035
APA StyleSy, I., Conrad, L., & Becker, S. L. (2022). Recent Advances and Potential Future Applications of MALDI-TOF Mass Spectrometry for Identification of Helminths. Diagnostics, 12(12), 3035. https://doi.org/10.3390/diagnostics12123035