
Application of Proteomics to the Study of the Therapeutics and Pathogenicity of Giardia duodenalis
 , and
, and
Abstract
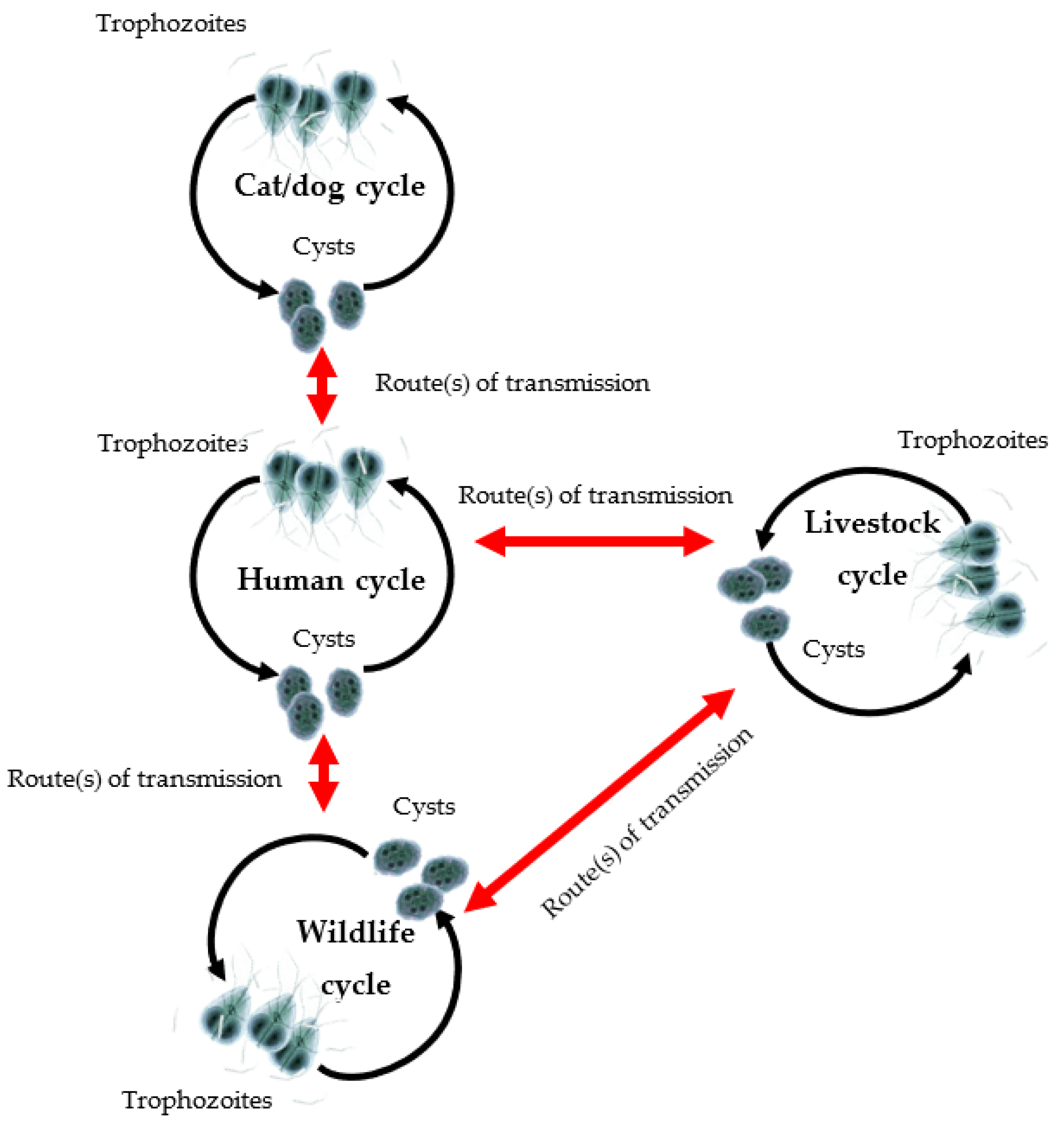
1. Introduction
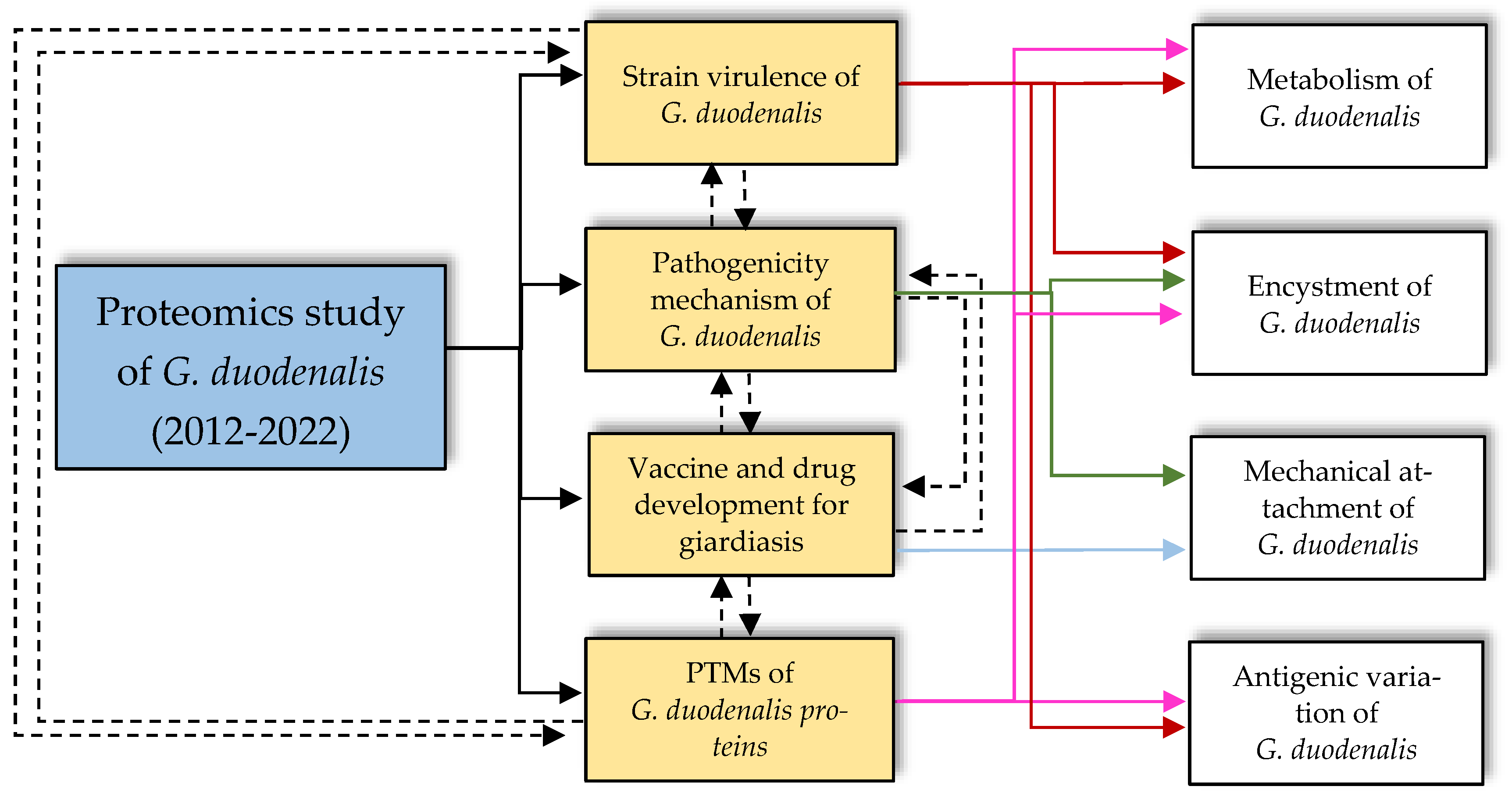
2. Applications of Proteomics
2.1. Investigating Protein Targets for Vaccine and Chemotherapy Development
2.2. Investigating the Strain Virulence of G. duodenalis
2.3. Understanding the Pathogenicity Mechanism of G. duodenalis
2.4. Investigating the Post-Translational Modifications of Giardia Proteins
3. Future Directions
4. Potential and Limitations of Current Proteomics Studies in Biomedical Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hemphill, A.; Müller, N.; Müller, J. Comparative pathobiology of the intestinal protozoan parasites Giardia lamblia, Entamoeba histolytica, and Cryptosporidium parvum. Pathogens 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Argüello-García, R.; Ortega-Pierres, M.G. Giardia duodenalis Virulence—“To Be, or Not To Be”. Curr. Trop. Med. Rep. 2021, 8, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Emery-Corbin, S.J.; Vuong, D.; Lacey, E.; Svärd, S.G.; Ansell, B.R.E.; Jex, A.R. Proteomic diversity in a prevalent human-infective Giardia duodenalis sub-species. Int. J. Parasitol. 2018, 48, 817–823. [Google Scholar] [CrossRef]
- Adam, R.D. The biology of Giardia spp. Microbiol. Rev. 1991, 55, 706–732. [Google Scholar] [CrossRef]
- Litleskare, S.; Rortveit, G.; Eide, G.E.; Hanevik, K.; Langeland, N.; Wensaas, K.A. Prevalence of irritable bowel syndrome and chronic fatigue 10 years after Giardia infection. Clin. Gastroenterol. Hepatol. 2018, 16, 1064–1072. [Google Scholar] [CrossRef]
- Hooshyar, H.; Rostamkhani, P.; Arbabi, M.; Delavari, M. Giardia lamblia infection review of current diagnostic strategies. Gastroenterol. Hepatol. Bed Bench 2019, 12, 3–12. [Google Scholar]
- Fink, M.Y.; Shapiro, D.; Singer, S.M. Maintenance, lifecycle induction, and infection of murine models. Curr. Protoc. Microbiol. 2020, 57, e102. [Google Scholar] [CrossRef]
- Einarsson, E.; Svärd, S.G.; Troell, K. UV irradiation responses in Giardia intestinalis. Exp. Parasitol. 2015, 154, 25–32. [Google Scholar] [CrossRef]
- Dubourg, A.; Xia, D.; Winpenny, J.P.; Al Naimi, S.; Bouzid, M.; Sexton, D.W.; Wastling, J.M.; Hunter, P.R.; Tyler, K.M. Giardia secretome highlights secreted tenascins as a key component of pathogenesis. Gigascience 2018, 7, giy003. [Google Scholar] [CrossRef]
- Emery, S.J.; Van Sluyter, S.; Haynes, P.A. Proteomic analysis in Giardia duodenalis yields insights into strain virulence and antigenic variation. Proteomics 2014, 14, 2323–2534. [Google Scholar] [CrossRef] [PubMed]
- Faso, C.; Bischof, S.; Hehl, A.B. The proteome landscape of Giardia lamblia encystation. PLoS ONE 2013, 8, e83207. [Google Scholar] [CrossRef] [PubMed]
- Ma’ayeh, S.Y.; Liu, J.; Peirasmaki, D.; Hörnaeus, K.; Bergström Lind, S.; Grabherr, M.; Bergquist, J.; Svärd, S.G. Characterization of the Giardia intestinalis secretome during interaction with human intestinal epithelial cells: The impact on host cells. PLoS Negl. Trop. Dis. 2017, 11, e0006120. [Google Scholar] [CrossRef] [PubMed]
- Einarsson, E.; Svärd, S.G. Encystation of Giardia intestinalis—A journey from the duodenum to the colon. Curr. Trop. Med. Rep. 2015, 2, 101–109. [Google Scholar] [CrossRef]
- Emery, S.J.; Baker, L.; Ansell, B.R.E.; Mirzaei, M.; Haynes, P.A.; McConville, M.J.; Svärd, S.G.; Jex, A.R. Differential protein expression and post-translational modifications in metronidazole-resistant Giardia duodenalis. Gigascience 2018, 7, giy024. [Google Scholar] [CrossRef]
- Balan, B.; Emery-Corbin, S.J.; Sandow, J.J.; Ansell, B.R.E.; Tichkule, S.; Webb, A.I.; Svärd, S.G.; Jex, A.R. Multimodal regulation of encystation in Giardia duodenalis revealed by deep proteomics. Int. J. Parasitol. 2021, 51, 809–824. [Google Scholar] [CrossRef]
- Emery, S.J.; Lacey, E.; Haynes, P.A. Quantitative proteomic analysis of Giardia duodenalis assemblage A: A baseline for host, assemblage, and isolate variation. Proteomic 2015, 15, 2281–2285. [Google Scholar] [CrossRef]
- Lourenço, D.; Andrade, I.D.S.; Terra, L.L.; Guimarães, P.R.; Zingali, R.B.; de Souza, W. Proteomic analysis of the ventral disc of Giardia lamblia. BMC Res. Notes 2012, 5, 41. [Google Scholar] [CrossRef]
- Camerini, S.; Bocedi, A.; Cecchetti, S.; Casella, M.; Carbo, M.; Morea, V.; Pozio, E.; Ricci, G.; Lalle, M. Proteomic and functional analyses reveal pleiotropic action of the anti-tumoral compound NBDHEX in Giardia duodenalis. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 147–158. [Google Scholar] [CrossRef]
- Davids, B.J.; Liu, C.M.; Hanson, E.M.; Le, C.H.Y.; Ang, J.; Hanevik, K.; Fischer, M.; Radunovic, M.; Langeland, N.; Ferella, M.; et al. Identification of conserved candidate vaccine antigens in the surface proteome of Giardia lamblia. Infect. Immun. 2019, 87, e00219-19. [Google Scholar] [CrossRef]
- Olson, M.E.; Ceri, H.; Morck, D.W. Giardia vaccination. Parasitol. Today 2000, 16, 213–217. [Google Scholar] [CrossRef]
- Ansell, B.R.E.; McConville, M.J.; Ma’ayeh, S.Y.; Dagley, M.J.; Gasser, R.B.; Svärd, S.G.; Jex, A.R. Drug resistance in Giardia duodenalis. Biotechnol. Adv. 2015, 33, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M. Giardiasis in the post genomic era: Treatment, drug resistance and novel therapeutic perspectives. Infect. Disord.-Drug Targets 2010, 10, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Tak, V.; Mirdha, B.R.; Makharia, G.K. Refractory giardiasis: A molecular appraisal from a tertiary care centre in India. Indian J. Med. Microbiol. 2014, 32, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Lingdan, L.; Pengtao, G.; Wenchao, L.; Jianhua, L.; Ju, Y.; Chengwu, L.; He, L.; Guocai, Z.; Wenzhi, R.; Yujiang, C.; et al. Differential dissolved protein expression throughout the life cycle of Giardia lamblia. Exp. Parasitol. 2012, 132, 465–469. [Google Scholar] [CrossRef]
- Müller, J.; Braga, S.; Heller, M.; Müller, N. Resistance formation to nitro drugs in Giardia lamblia: No common markers identified by comparative proteomics. Int. J. Parasitol. Drugs Drug Resist. 2019, 9, 112–119. [Google Scholar] [CrossRef]
- Paz-Maldonado, M.T.; Argüello-García, R.; Cruz-Soto, M.; Mendoza-Hernández, G.; Ortega-Pierres, G. Proteomic and transcriptional analyses of genes differentially expressed in Giardia duodenalis clones resistant to albendazole. Infect. Genet. Evol. 2013, 15, 10–17. [Google Scholar] [CrossRef]
- Matadamas-Martínez, F.; Castillo, R.; Hernández-Campos, A.; Méndez-Cuesta, C.; de Souza, W.; Gadelha, A.P.; Nogueda-Torres, B.; Hernández, J.M.; Yépez-Mulia, L. Proteomic and ultrastructural analysis of the effect of a new nitazoxanide-N-methyl-1H-benzimidazole hybrid against Giardia intestinalis. Res. Vet. Sci. 2016, 105, 171–179. [Google Scholar] [CrossRef]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and their applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef]
- Davids, B.J.; Mehta, K.; Fesus, L.; McCaffery, J.M.; Gillin, F.D. Dependence of Giardia lamblia encystation on novel transglutaminase activity. Mol. Biochem. Parasitol. 2004, 136, 173–180. [Google Scholar] [CrossRef]
- Lalle, M.; Bavassano, C.; Fratini, F.; Cecchetti, S.; Boisguerin, P.; Crescenzi, M.; Pozio, E. Involvement of 14-3-3 protein post-translational modifications in Giardia duodenalis encystation. Int. J. Parasitol. 2010, 40, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Camerini, S.; Cecchetti, S.; Finelli, R.; Sferra, G.; Müller, J.; Ricci, G.; Pozio, E. The FAD-dependent glycerol-3-phosphate dehydrogenase of Giardia duodenalis: An unconventional enzyme that interacts with the g14-3-3 and it is a target of the antitumoral compound NBDHEX. Front. Microbiol. 2015, 6, 544. [Google Scholar] [CrossRef] [PubMed]
- Langford, T.D.; Housley, M.P.; Boes, M.; Chen, J.; Kagnoff, M.F.; Gillin, F.D.; Eckmann, L. Central importance of immunoglobulin A in host defense against Giardia spp. Infect. Immun. 2002, 70, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E. Surface antigenic variation in Giardia lamblia. In Antigenic Variation; Elsevier: Amsterdam, The Netherlands, 2003; pp. 357–374. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef]
- Franzén, O.; Jerlström-Hultqvist, J.; Castro, E.; Sherwood, E.; Ankarklev, J.; Reiner, D.S.; Palm, D.; Andersson, J.O.; Andersson, B.; Svärd, S.G. Draft genome sequencing of Giardia intestinalis assemblage B isolate GS: Is human giardiasis caused by two different species? PLoS Pathog. 2009, 5, e1000560. [Google Scholar] [CrossRef]
- Jerlström-Hultqvist, J.; Franzén, O.; Ankarklev, J.; Xu, F.; Nohýnková, E.; Andersson, J.O.; Svärd, S.G.; Andersson, B. Genome analysis and comparative genomics of a Giardia intestinalis assemblage E isolate. BMC Genom. 2010, 11, 543. [Google Scholar] [CrossRef]
- Coelho, C.H.; Costa, A.O.; Silva, A.C.C.; Pucci, M.M.; Serufo, A.V.; Busatti, H.G.N.O.; Durigan, M.; Perales, J.; Chapeaurouge, A.; da Silva, E.; et al. Genotyping and descriptive proteomics of a potential zoonotic canine strain of Giardia duodenalis, infective to mice. PLoS ONE 2016, 11, e0164946. [Google Scholar] [CrossRef]
- Monis, P.; Caccio, S.; Thompson, R. Variation in Giardia; towards a taxonomic revision of the genus. Trends Parasitol. 2009, 25, 93–100. [Google Scholar] [CrossRef]
- Nolan, M.J.; Jex, A.R.; Upcroft, J.A.; Upcroft, P.; Gasser, R.B. Barcoding of Giardia duodenalis isolates and derived lines from an established cryobank by a mutatiob scanning-based approach. Electrophoresis 2011, 32, 2075–2090; [Google Scholar] [CrossRef]
- Upcroft, J.A.; McDonnell, P.A.; Gallagher, A.N.; Chen, N.; Upcroft, P. Lethal Giardia from a wild-caught sulphur-crested cockatoo (Cacatua galerita) established in vitro chronically infects mice. Parasitology 1997, 114, 407–412. [Google Scholar] [CrossRef]
- Ramírez, M.A.; Lorenz, M.C. Mutations in alternative carbon utilization pathways in Candida albicans attenuate virulence and confer pleiotropic phenotypes. Eukaryot. Cell 2007, 6, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Yanze-Kontchou, C.; Gschwind, N. Mineralization of the herbicide atrazine as a carbon source by a Pseudomonas strain. Appl. Environ. Microbiol. 1994, 60, 4297–4302. [Google Scholar] [CrossRef] [PubMed]
- Waligora, E.A.; Fisher, C.R.; Hanovice, N.J.; Rodou, A.; Wyckoff, E.E.; Payne, S.M. Role of intracellular carbon metabolism pathways in Shigella flexneri virulence. Infect. Immun. 2014, 82, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Itagaki, T.; Kawabata, T.; Konaka, T.; Muraoka, N.; Saeki, H.; Kanai, K.; Chikazawa, S.; Hori, Y.; Hoshi, F.; et al. Prevalence of intestinal parasites and genotyping of Giardia intestinalis in pet shop puppies in east Japan. Vet. Parasitol. 2011, 176, 74–78. [Google Scholar] [CrossRef]
- Paoletti, B.; Iorio, R.; Capelli, G.; Sparagano, O.A.E.; Giangaspero, A. Epidemiological scenario of giardiasis in dogs from central Italy. Ann. N. Y. Acad. Sci. 2008, 1149, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Jedelský, P.L.; Doležal, P.; Rada, P.; Pyrih, J.; Šmíd, O.; Hrdý, I.; Šedinová, M.; Marcinčiková, M.; Voleman, L.; Perry, A.J.; et al. The minimal proteome in the reduced mitochondrion of the parasitic protist Giardia intestinalis. PLoS ONE 2011, 6, e17285. [Google Scholar] [CrossRef]
- Lauwaet, T.; Smith, A.J.; Reiner, D.S.; Romijn, E.P.; Wong, C.C.L.; Davids, B.J.; Shah, S.A.; Yates, J.R.; Gillin, F.D. Mining the Giardia genome and proteome for conserved and unique basal body proteins. Int. J. Parasitol. 2011, 41, 1079–1092. [Google Scholar] [CrossRef]
- Stastna, M.; Van Eyk, J.E. Secreted proteins as a fundamental source for biomarker discovery. Proteomics 2012, 12, 722–735. [Google Scholar] [CrossRef]
- Liu, L.; Yang, Y.; Fang, R.; Zhu, W.; Wu, J.; Li, X.; Patankar, J.V.; Li, W. Giardia duodenalis and its secreted PPIB trigger inflammasome activation and pyroptosis in macrophages through TLR4-induced ROS signalling and A20-mediated NLRP3 deubiquitination. Cells 2021, 10, 3425. [Google Scholar] [CrossRef]
- Liu, J.; Ma’ayeha, S.; Peirasmakia, D.; Lundström-Stadelmannb, B.; Hellmana, L.; Svärda, S.G. Secreted Giardia intestinalis cysteine proteases disrupt intestinal epithelial cell junctional complexes and degrade chemokines. Virulence 2018, 9, 879–894. [Google Scholar] [CrossRef]
- Ringqvist, E.; Palm, J.E.D.; Skarin, H.; Hehl, A.B.; Weiland, M.; Davids, B.J.; Reiner, D.S.; Griffiths, W.J.; Eckmann, L.; Gillin, F.D.; et al. Release of metabolic enzymes by Giardia in response to interaction with intestinal epithelial cells. Mol. Biochem. Parasitol. 2008, 159, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yichoy, M.; Duarte, T.T.; de Chatterjee, A.; Mendez, T.L.; Aguilera, K.Y.; Roy, D.; Roychowdhury, S.; Aley, S.B.; Das, S. Lipid metabolism in Giardia: A post-genomic perspective. Parasitology 2011, 138, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.C.; Teoh, D.A.; Scott, K.G.E.; Meddings, J.B.; Macnaughton, W.K.; Buret, A.G. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infect. Immun. 2002, 70, 3673–3680. [Google Scholar] [CrossRef] [PubMed]
- Humen, M.A.; Pérez, P.F.; Liévin-Le Moal, V. Lipid raft-dependent adhesion of Giardia intestinalis trophozoites to a cultured human enterocyte-like Caco-2/TC7 cell monolayer leads to cytoskeleton-dependent functional injuries. Cell. Microbiol. 2011, 13, 1683–1702. [Google Scholar] [CrossRef] [PubMed]
- Maia-Brigagão, C.; Morgado-Díaz, J.A.; de Souza, W. Giardia disrupts the arrangement of tight, adherens and desmosomal junction proteins of intestinal cells. Parasitol. Int. 2012, 61, 280–287. [Google Scholar] [CrossRef]
- Bhargava, A.; Cotton, J.A.; Dixon, B.R.; Gedamu, L.; Yates, R.M.; Buret, A.G. Giardia duodenalis surface cysteine proteases induce cleavage of the intestinal epithelial cytoskeletal protein villin via myosin light chain kinase. PLoS ONE 2015, 10, 0136102. [Google Scholar] [CrossRef]
- Sonda, S.; Morf, L.; Bottova, I.; Baetschmann, H.; Rehrauer, H.; Caflisch, A.; Hakimi, M.A.; Hehl, A.B. Epigenetic mechanisms regulate stage differentiation in the minimized protozoan Giardia lamblia. Mol. Microbiol. 2010, 76, 48–67. [Google Scholar] [CrossRef]
- Slavin, I.; Saura, A.; Carranza, P.G.; Touz, M.C.; Nores, M.J.; Luján, H.D. Dephosphorylation of cyst wall proteins by a secreted lysosomal acid phosphatase is essential for excystation of Giardia lamblia. Mol. Biochem. Parasitol. 2002, 122, 95–98. [Google Scholar] [CrossRef]
- Fiorillo, A.; di Marino, D.; Bertuccini, L.; Via, A.; Pozio, E.; Camerini, S.; Ilari, A.; Lalle, M. The crystal structure of Giardia duodenalis 14-3-3 in the apo form: When protein post-translational modifications make the difference. PLoS ONE 2014, 9, e92902. [Google Scholar] [CrossRef]
- Merino, M.C.; Zamponi, N.; Vranych, C.V.; Touz, M.C.; Rópolo, A.S. Identification of Giardia lamblia DHHC proteins and the role of protein S-palmitoylation in the encystation process. PLoS Negl. Trop. Dis. 2014, 8, e2997. [Google Scholar] [CrossRef]
- Zhu, W.; Jiang, X.; Sun, H.; Li, Y.; Shi, W.; Zheng, M.; Liu, D.; Ma, A.; Feng, X. Global lysine acetylation and 2-hydroxyisobutyrylation profiling reveals the metabolism conversion mechanism in Giardia lamblia. Mol. Cell. Proteom. 2021, 20, e10043. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Braga, S.; Uldry, A.C.; Heller, M.; Müller, N. Comparative proteomics of three Giardia lamblia strains: Investigation of antigenic variation in the post-genomic era. Parasitology 2020, 147, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Emery-Corbin, S.J.; Hamey, J.J.; Balan, B.; Rojas-López, L.; Svärd, S.G.; Jex, A.R. Eukaryote-conserved histone post-translational modification landscape in Giardia duodenalis revealed by mass spectrometry. Int. J. Parasitol. 2021, 51, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Carranza, P.G.; Gargantini, P.R.; Prucca, C.G.; Torri, A.; Saura, A.; Svärd, S.; Lujan, H.D. Specific histone modifications play critical roles in the control of encystation and antigenic variation in the early-branching eukaryote Giardia lamblia. Int. J. Biochem. Cell Biol. 2016, 81, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Miyamoto, Y.; Leitsch, D.; Santini, J.; Debnath, A.; Gut, J.; McKerrow, J.H.; Reed, S.L.; Eckmann, L. A reprofiled drug, auranofin, is effective against metronidazole-resistant Giardia lamblia. Antimicrob. Agents Chemother. 2013, 57, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F. Cryptosporidium and Giardia: Treatment options and prospects for new drugs. Exp. Parasitol. 2010, 124, 44–53. [Google Scholar] [CrossRef]
- Najdrová, V.; Stairs, C.W.; Vinopalová, M.; Voleman, L.; Doležal, P. The evolution of the Puf superfamily of proteins across the tree of eukaryotes. BMC Biol. 2020, 18, 77. [Google Scholar] [CrossRef]
- Sudhakar, A.; Kamanna, S.; Bojja, M.; Tatu, U. Proteomic analysis of Giardia lamblia and Trichomonas vaginalis flagella reveal unique post-translational modifications in tubulin that provide clues to regulation of their motilities. Proteomics 2021, 21, 2100004. [Google Scholar] [CrossRef]
- Gourguechon, S.; Holt, L.J.; Cande, W.Z. The Giardia cell cycle progresses independently of the anaphase-promoting complex. J. Cell. Sci. 2013, 126, 2246–2255. [Google Scholar] [CrossRef]
- Peirasmaki, D.; Ma’ayeh, S.Y.; Xu, F.; Ferella, M.; Campos, S.; Liu, J.; Svärd, S.G. High cysteine membrane proteins (HCMPS) are up-regulated during Giardia-host cell interactions. Front. Genet. 2020, 11, 913. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.; Liu, M.; Qin, X.; Yu, X.; Zhao, H.; Li, X.; Li, W. COX-2 is required to mediate crosstalk of ROS-dependent activation of MAPK/NF-κB signaling with pro-inflammatory response and defense-related NO enhancement during challenge of macrophage-like cell line with Giardia duodenalis. PLoS Negl. Trop. Dis. 2022, 16, e0010402. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Purpose of Proteomics Study | Approach | Subtypes of G. duodenalis | Target Protein | Database | Software Programme | Remarks/Advantages | References | |
|---|---|---|---|---|---|---|---|---|---|
| Assemblage | Strain | ||||||||
| 1. | Mechanical attachment of G. duodenalis | 1D-sodium dodecyl-sulphate polyacrylamide gel electrophoresis (SDS PAGE), 2D-SDS PAGE, nano liquid chromatography-tandem mass spectrometry (nLC-MS/MS) and MALDI-TOF/TOF MS | A | WBC6 strain | Ventral disc | NCBI-nr | MASCOT |
| [18] |
| 2. | Encystment of G. duodenalis | iTRAQ labelling, SFX fractionation, and reversed-phase nano (RPLC-MS/MS) | D | Changchun strain | Soluble proteins from trophozoites and cysts | GiardiaDB | MASCOT |
| [25] |
| 3. | Evaluation of drug treatments/vaccine constructions on G. duodenalis | 2D-PAGE, isoelectrofocusing (IEF), and LC-MS/MS | A | WB strain | Albendazole (ABZ)-resistant and -susceptible trophozoites | NCB-nr | MASCOT v1.6b9 |
| [27] |
| 4. | 2D-PAGE and MALDI-TOF MS | A | WBC6 strain | Cytoskeleton proteins in Giardia trophozoites | NCBI-nr | MASCOT v2.3 |
| [28] | |
| 5. | 1D-NuPAGE 4-12%, Western blot analysis, and LC-MS/MS | A | WBC6 strain | Proteins in Giardia trophozoites | GiardiaDB, UniProt, Protein Data Bank (PDB) | ELM, BLASTp MutAlin, SwissModel, I-TASSER, HHpred and Phyre2 PsiPred, and Disopred3 |
| [19] | |
| 6. | Western blot analysis, streptavidin affinity chromatography, SDS-PAGE, and high-pressure LC-MS/MS (HPLC-MS/MS) | A | WBC6 strain | Surface proteins of Giardia trophozoites | GiardiaDB | ProteinPilot v2.0, SignalP server v3.0, and CSS-Palm |
| [20] | |
| B | GS/M strain | ||||||||
| 7. | nLC-MS/MS | A1 | C4 strain | Nitazoxanide (NTZ)- and metronidazole (MTZ)-resistant trophozoites | GiardiaDB | MaxQuant v1.5 |
| [26] | |
| 1062ID10 strain | |||||||||
| 713M3 strain | |||||||||
| 8. | Virulence of G. duodenalis | 1D-SDS PAGE, nLC-MS/MS, filter-aided sample preparation (FASP), and gas-phase fractionation (GPF) | A1 | BRIS/95/HEPU/2041 | Variant surface proteins (VSPs) | GiardiaDB | Global proteome machine (GPM) and X1Tandem |
| [11] |
| BRIS/83/HEPU/106 | |||||||||
| 9. | 2D-SDS PAGE and MALDI-TOF MS | A | BHF1C strain | Soluble and insoluble proteins in trophozoites | GiardiaDB | MASCOT v2.1, Protein Blast, and Panther |
| [38] | |
| Portland-1 strain | |||||||||
| 10. | Pathogenicity mechanism of G. duodenalis | nLC-MS/MS, ELISA, SDS-PAGE, and Western blot analysis | A | WBC6 strain | Excretory–secretory products (ESPs) of Giardia trophozoites | GiardiaDB | Proteome Discoverer v1.4 and MudPIT |
| [13] |
| B | GSH7 strain | ||||||||
| 11. | LC-MS/MS | A | WB-1B strain | Trophozoites, low-bile primed trophozoites, and the 16 h post-induction of encystation and mature cysts | UniProt, PFAM | MaxQuant v1.5.8.3, STRING v11, I-TASSER, and Auto Dock Tools |
| [16] | |
| 12. | Post-translational modifications (PTMs) of Giardia proteins | 1D NuPAGE 4-12% and rpLC-MS/MS | A | WBC6 strain | 14-3-3 proteins | PDB | Program Phaser, COOT, Refmac5, PyMOL, secondary structure matching (Superpose), and NAMD v2.8 |
| [60] |
| 13. | SDS-PAGE, Western blot, chloroform/methanol extraction, and flow cytometer | A | WBC1267 strain | DHHC proteins (also known as protein acyltransferases) in the trophozoites and encysting cells of Giardia | Ensembl, NCBI, PFAM, GiardiaDB and UniProt | CD-HIT, PROMALS3D, ProtTest, CSS-Palm v3.0, HMMTOP, SMART, TMHMM, TMPred, and Block Mapping and Gathering with Entropy (BMGE) |
| [61] | |
| 14. | Western blot analysis, SDS-PAGE and nLC-MS/MS | A1 | WB-M3 strain | MTZ-resistant and -susceptible trophozoites | GiardiaDB, UniProt and PRIDE | Search Tool for the Retrieval of Interacting Genes (STRING), Xcalibur, Proteome Discoverer v1.3, MASCOT, and DAVID bioinformatics resource |
| [15] | |
| BRIS/83/HEPU/106-2ID10 | |||||||||
| BRIS/83/HEPU/713-M3 | |||||||||
| 15. | Trifluoroacetic acid (TFA) extraction and LC-MS/MS | A | WBC6 strain | VSPs in G. duodenalis trophozoites | GiardiaDB | MaxQuant v1.5.4.1 |
| [63] | |
| WBA1 strain | |||||||||
| B | GS/M-83-H7 strain | ||||||||
| 16. | Acetone precipitation, SDS-PAGE, and nLC-MS/MS | A | WB-1B strain | Histone proteins | GiardiaDBand UniProt | MaxQuant v1.5.8.3, PRIDE, CLUSTALW and Aline programme v1.0.025 |
| [64] | |
| 17. | Reverse-phase high-performance liquid chromatography (rpHPLC), SDS-PAGE, and iTRAQ labelling | A | WBC2 strain | Trophozoite of G. duodenalis under sugar starvation | UniProt, CORUM, and STRING v10 | MaxQuant, R Project, and Cytoscape v3.2.1 |
| [62] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aziz, A.F.E.; Roshidi, N.; Othman, N.; Mohd Hanafiah, K.; Arifin, N. Application of Proteomics to the Study of the Therapeutics and Pathogenicity of Giardia duodenalis. Diagnostics 2022, 12, 2744. https://doi.org/10.3390/diagnostics12112744
Aziz AFE, Roshidi N, Othman N, Mohd Hanafiah K, Arifin N. Application of Proteomics to the Study of the Therapeutics and Pathogenicity of Giardia duodenalis. Diagnostics. 2022; 12(11):2744. https://doi.org/10.3390/diagnostics12112744
Chicago/Turabian StyleAziz, Ahmad Fudail Eiyad, Norhamizah Roshidi, Nurulhasanah Othman, Khayriyyah Mohd Hanafiah, and Norsyahida Arifin. 2022. "Application of Proteomics to the Study of the Therapeutics and Pathogenicity of Giardia duodenalis" Diagnostics 12, no. 11: 2744. https://doi.org/10.3390/diagnostics12112744
APA StyleAziz, A. F. E., Roshidi, N., Othman, N., Mohd Hanafiah, K., & Arifin, N. (2022). Application of Proteomics to the Study of the Therapeutics and Pathogenicity of Giardia duodenalis. Diagnostics, 12(11), 2744. https://doi.org/10.3390/diagnostics12112744