
Detection of Babesia odocoilei in Humans with Babesiosis Symptoms


Abstract
1. Introduction
2. Results
3. Discussion
3.1. Meeting Ethical Requirements for Babesia Study
3.2. Age as a Factor
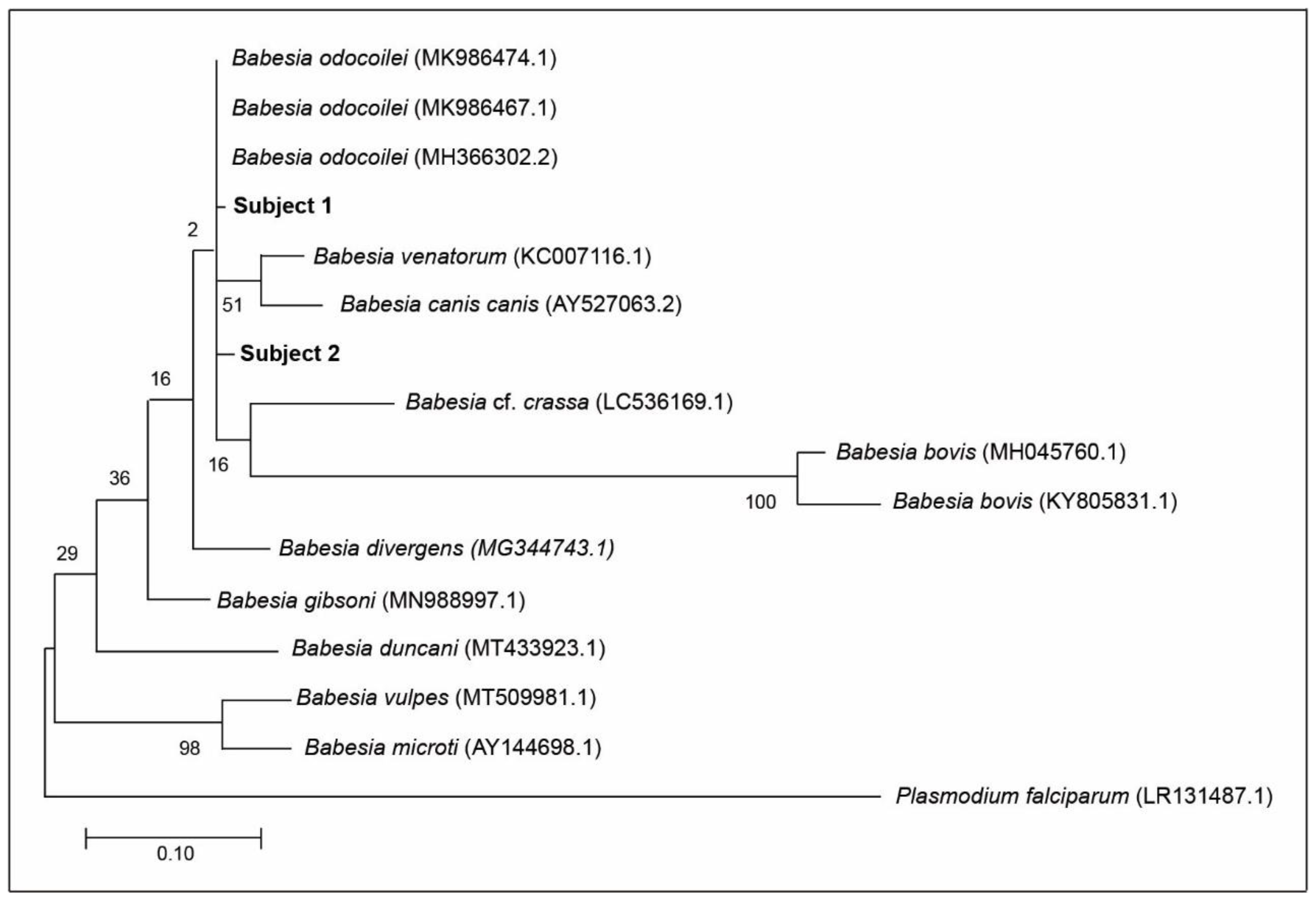
3.3. Source of B. odocoilei Infection
3.4. Testing as a Factor
3.5. Serology as a Factor
3.6. Cross-Reactivity
3.7. Blood Smear Performance
3.8. Virulence of B. odocoilei
3.9. Contracting Human Babesiosis in Non-Endemic Areas
3.10. Persistence of B. odocoilei
3.11. Molecular Identification
3.12. Number of Subjects
4. Materials and Methods
4.1. Participation in Study
4.2. Collecting Blood Samples
4.3. Informed Consent
4.4. Babesia Detection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babes, V. Sur l’hémoglobinurie bactérienne du boeuf. C. R. Acad. Sci. 1888, 107, 692–694. [Google Scholar]
- Smith, T.; Kilborne, F.L. Investigation into the Nature, Causation, and Prevention of Southern Cattle Fever, 1st ed.; United States Government Printing Office: Washington, DC, USA, 1893; pp. 85–116.
- Škrabalo, Z.; Deanović, Z. Piroplasmosis in man: Report on a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar] [PubMed]
- Scholtens, R.G.; Braff, E.H.; Healy, R.R.; Gleason, N. A case of babesiosis in man in the United States. Am. J. Trop. Med. Hyg. 1968, 17, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Western, K.A.; Benson, G.D.; Gleason, N.N.; Healy, G.R.; Schultz, M.G. Babesiosis in a Massachusetts resident. N. Engl. J. Med. 1970, 283, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Villatoro, T.; Karp, J.K. Transfusion-transmitted babesiosis. Arch. Pathol. Lab. Med. 2019, 143, 130–134. [Google Scholar] [CrossRef]
- Tonnetti, L.; Townsend, R.L.; Dodd, R.Y.; Stramer, S.L. Characteristics of transfusion-transmitted Babesia microti, American Red Cross, 2010−2017. Transfusion 2019, 59, 2908–2912. [Google Scholar] [CrossRef]
- Brennan, M.B.; Herwaldt, B.L.; Kazmierczak, J.J.; Weiss, J.W.; Klein, C.L.; Leith, C.P.; He, R.; Oberley, M.J.; Tonnetti, L.; Wilkins, P.P.; et al. Transmission of Babesia microti parasites by solid organ transplantation. Emerg. Infect. Dis. 2016, 22, 1869–1876. [Google Scholar] [CrossRef]
- New, D.L.; Quinn, J.B.; Qureshi, M.Z.; Sigler, S.J. Vertically transmitted babesiosis. J. Pediatr. 1997, 131, 163–164. [Google Scholar] [CrossRef]
- Fox, L.M.; Winger, S.; Ahmed, A.; Arnold, A.; Chou, J.; Rhein, L.; Levy, O. Neonatal babesiosis: Case report and review of the literature. Pediatr. Infect. Dis. J. 2006, 25, 169–173. [Google Scholar] [CrossRef]
- Cornett, J.K.; Malhotra, A.; Hart, D. Vertical transmission of babesiosis from a pregnant, splenectomized mother to her neonate. Infect. Dis. Clin. Pract. 2012, 20, 408–410. [Google Scholar] [CrossRef]
- Iyer, S.; Goodman, K. Congenital babesiosis from maternal exposure: A case report. J. Emerg. Med. 2019, 56, E39–E41. [Google Scholar] [CrossRef]
- Vannier, E.; Krause, P.J. Human babesiosis. N. Engl. J. Med. 2012, 366, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Kinderlehrer, D.A. Babesia. In Recovery from Lyme Disease: The Integrative Medicine Guide to Diagnosing and Treating Tick-Borne Illness; Skyhorse Publishing: New York, NY, USA, 2021; pp. 93–101. ISBN 978-1-5107-6205-3. [Google Scholar]
- Boustani, M.; Gelfand, J.A. Babesiosis. Clin. Infect. Dis. 1996, 22, 611–615. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, Z.; Li, H.; Gao, X.; Bian, A.; Yan, H.; Kong, D.; Liu, X. Human babesiosis in China: A systematic review. Parasitol. Res. 2019, 118, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.M.; Liu, L.P.; Chung, W.C.; Ong, S.J.; Wang, C.C. Human babesiosis in Taiwan: Asymptomatic infection with a Babesia microti-like organism in a Taiwanese woman. J. Clin. Microbiol. 1997, 35, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Conrad, P.A. Human babesiosis: An emerging tick-borne disease. Int. J. Parasitol. 2000, 30, 1323–1337. [Google Scholar] [CrossRef]
- Herwaldt, B.L.; Persing, D.H.; Precigout, E.A.; Goff, W.L.; Mathiesen, D.A.; Taylor, P.W.; Eberhard, M.L.; Gorenflot, A.F. A fatal case of babesiosis in Missouri: Identification of another piroplasm that infects humans. Ann. Intern. Med. 1996, 124, 643–650. [Google Scholar] [CrossRef]
- Ord, R.L.; Lobo, C.A. Human babesiosis: Pathogens, prevalence, diagnosis, and treatment. Cur. Clin. Microbiol. Rep. 2015, 2, 173–181. [Google Scholar] [CrossRef]
- Holman, P.J.; Madeley, J.; Craig, T.M.; Allsopp, B.A.; Allsopp, M.T.; Petrini, K.R.; Waghela, S.D.; Wagner, G.G. Antigenic, phenotypic and molecular characterization confirms Babesia odocoilei isolated from three cervids. J. Wildl. Dis. 2000, 36, 518–530. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Shein, E. The piroplasms: Life cycle and sexual stages. Adv. Parasitol. 1985, 23, 37–103. [Google Scholar]
- Nicholson, W.A.; Sonenshine, D.E.; Noden, B.H. Ticks (Ixodida). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: London, UK, 2019; pp. 603–672. ISBN 978-0-12-814043-7. [Google Scholar]
- Kaufman, R.W. Integument and ecdysis. In Biology of Ticks, 2nd ed.; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: Oxford, UK, 2013; Volume 1, pp. 99–196. ISBN 9780199744053. [Google Scholar]
- Perry, B.D.; Nichols, D.K.; Cullom, E.S. Babesia odocoilei Emerson and Wright, 1970 in white-tailed deer, Odocoileus virginianus (Zimmermann), in Virginia. J. Wildl. Dis. 1985, 21, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Holman, P.J.; Waldrup, K.A.; Wagner, G.G. In vitro cultivation of a Babesia isolated from a white-tailed deer (Odocoileus virginianus). J. Parasitol. 1988, 74, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Waldrup, K.A.; Kocan, A.A.; Qureshi, T.; Davis, D.S.; Baggett, D.; Wagner, G.G. Serological prevalence and isolation of Babesia odocoilei among white-tailed deer (Odocoileus virginianus) in Texas and Oklahoma. J. Wildl. Dis. 1989, 25, 194–201. [Google Scholar] [CrossRef]
- Pattullo, K.M.; Wobeser, G.; Lockerbie, B.P.; Burgess, H.J. Babesia odocoilei infection in a Saskatchewan elk (Cervus elaphus canadensis) herd. J. Vet. Diagn. Investig. 2013, 25, 535–540. [Google Scholar] [CrossRef]
- Bartlett, S.; Abou-Madi, N.; Messick, J.B.; Birkenheuer, A. Diagnosis and Treatment of Babaesia odocoilei in captive reindeer (Rangifer tarandus tarandus) and recognition of three novel host species. J. Zoo Wildl. Med. 2009, 40, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Schoelkopf, L.; Hutchinson, C.E.; Bendele, K.G.; Goff, W.L.; Willette, M.; Rasmussen, J.M.; Holman, P.J. New ruminant hosts and wider geographic range identified for Babesia odocoilei (Emerson and Wright 1970). J. Wildl. Dis. 2005, 41, 683–690. [Google Scholar] [CrossRef]
- Thomford, J.W.; Conrad, P.A.; Boyce, W.M.; Holman, P.J.; Jessup, D.A. Isolation and in vitro cultivation of Babesia parasites from free-ranging desert bighorn sheep (Ovis canadensis nelsoni) and mule deer (Odocoileus hemionus) in California. J. Parasitol. 1993, 79, 77–84. [Google Scholar] [CrossRef]
- Eshoo, M.W.; Carolan, H.E.; Massire, C.; Chou, D.M.; Crowder, C.D.; Rounds, M.A.; Phillipson, C.A.; Schutzer, S.E.; Ecker, D.J. Survey of Ixodes pacificus ticks in California reveals a diversity of microorganisms and a novel and widespread Anaplasmataceae species. PLoS ONE 2015, 10, e0135828. [Google Scholar] [CrossRef]
- Scott, J.D.; Clark, K.L.; Foley, J.E.; Bierman, B.C.; Durden, L.A. Far-reaching dispersal of Borrelia burgdorferi sensu lato-infected blacklegged ticks by migratory songbirds in Canada. Healthcare 2018, 6, 89. [Google Scholar] [CrossRef]
- Scott, J.D.; Clark, K.L.; Coble, N.M.; Ballantyne, T.R. Detection and transstadial passage of Babesia species and Borrelia burgdorferi sensu lato in ticks collected from avian and mammalian hosts in Canada. Healthcare 2019, 7, 155. [Google Scholar] [CrossRef]
- Milnes, E.L.; Thornton, G.; Léveillé, A.N.; Delinatte, P.; Barta, J.R.; Smith, D.A.; Nemeth, N. Babesia odocoilei and zoonotic pathogens identified from Ixodes scapularis ticks in southern Ontario, Canada. Ticks Tick Borne Dis. 2019, 10, 670–676. [Google Scholar] [CrossRef]
- Scott, J.D.; Clark, K.L.; Coble, N.M.; Ballantyne, T.R. Presence of Babesia odocoilei and Borrelia burgdorferi sensu stricto in a tick and dual parasitism of Amblyomma inornatum and Ixodes scapularis on a bird in Canada. Healthcare 2019, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Pascoe, E.L.; Sajid, M.S.; Foley, J.E. Detection of Babesia odocoilei in Ixodes scapularis ticks collected from songbirds in Ontario and Quebec, Canada. Pathogens 2020, 9, 781. [Google Scholar] [CrossRef]
- Scott, J.D.; Pascoe, E.L.; Sajid, M.S.; Foley, J.E. Detection of Babesia odocoilei in Ixodes scapularis ticks collected in southern Ontario, Canada. Pathogens 2021, 10, 327. [Google Scholar] [CrossRef]
- Berger, B.W. Dermatologic manifestations of Lyme disease. Rev. Infect. Dis. 1989, 11, S1475–S1481. [Google Scholar] [CrossRef]
- Johnson, L.; Shapiro, M.; Mankoff, J. Removing the mask of average treatment effects in chronic Lyme disease research using big data and subgroup analysis. Healthcare 2018, 6, 124. [Google Scholar] [CrossRef]
- Shah, J.S.; Mark, O.; Caolli, E.; Porui, A.; Horowitz, R.I.; Ashbaugh, A.D.; Ramasamy, R. A fluorescence in situ hybridization (FISH) test for diagnosing babesiosis. Diagnostics 2020, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Penzhorn, B.L.; Oosthuizen, M.C. Babesia species of domestic cats: Molecular characterization has opened Pandora’s box. Front. Vet. Sci. 2020, 7, 134. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Gen. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Cacciò, S.; Gherlinzoni, F.; Aspöck, H.; Siemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; et al. Molecular characterization of a non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.-C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2016, 13, 65–70. [Google Scholar]
- Wilson, M.; Glaser, K.C.; Adams-Fish, D.; Boley, M.; Mayda, M.; Molestina, R.E. Development of droplet digital PCR for the detection of Babesia microti and Babesia duncani. Exp. Parasitol. 2015, 149, 24–31. [Google Scholar] [CrossRef]
- Rawls, W. Understanding Babesia. Available online: www.rawlsmd.com (accessed on 3 March 2017).
- Wozniak, R.A.; Lowenstine, L.J.; Hemmer, R.; Robinson, T.; Conrad, P.A. Comparative pathogenesis of human WA1 and Babesia microti isolates in a Syrian hamster model. Lab. Anim. Sci. 1996, 46, 507–515. [Google Scholar]
- Abraham, A.; Brasov, I.; Thekkiniath, J.; Kilian, N.; Lawres, L.; Gao, R.; DeBus, K.; He, L.; Yu, X.; Zhu, G.; et al. Establishment of a continuous in vitro culture of Babesia duncani in human erythrocytes reveals unusually high tolerance to recommended therapies. J. Biol. Chem. 2018, 293, 19974–19981. [Google Scholar] [CrossRef]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B.; Augagneur, Y.; Brès, V.; Duclos, A.; Randazzo, S.; et al. Sequencing of the smallest apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 18. [Google Scholar] [CrossRef]
- Gelfand, J.A. Babesia. In Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases, 4th ed.; Mandell, G.L., Bennett, J.E., Doliln, R., Eds.; Churchill Livingstone: New York, NY, USA, 1995; Volume 2, pp. 2497–2500. [Google Scholar]
- Bloch, E.M.; Kumar, S.; Krause, P.J. Persistence of Babesia microti infection in humans. Pathogens 2019, 8, 102. [Google Scholar] [CrossRef]
- Wang, S.; Li, M.; Luo, X.; Yu, L.; Nie, Z.; Liu, Q.; An, X.; Ao, Z.; Liu, Q.; Chen, J.; et al. Inhibitory effects of fosmidomycin against Babesia microti in vitro. Front. Cell Dev. Biol. 2020, 8, 247. [Google Scholar] [CrossRef]
- Falagas, M.E.; Klempner, M.S. Babesiosis in patients with AIDS: A chronic infection presenting as fever of unknown origin. Clin. Infec. Dis. 1996, 22, 809–812. [Google Scholar] [CrossRef]
- Lobo, C.A.; Cursino-Santos, J.F.; Singh, M.; Rodriguez, M. Babesia divergens: A drive to survive. Pathogens 2019, 8, 95. [Google Scholar] [CrossRef]
- Krause, J.K.; Lepore, T.; Sikand, V.; Gadbaw, J.; Gurke, G.; Telford, S.R.; Brassard, P.; Pearl, D.; Azlanzadeh, J.; Christianson, D.; et al. Atovaquone and azithromycin for the treatment of babesiosis. N. Eng. J. Med. 2000, 343, 1454–1458. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Subject | Age | Gender | Tick Bite | Lab Testing | Prior Treatment | Symptoms | GenBank Accession No. |
|---|---|---|---|---|---|---|---|
| 1 | 23 | female | not recalled | none, clinical diagnosis | atovaquone, symptoms unresolved | -night sweats -chills -fevers -profound fatigue -increased thirst -muscle aches -sleep disturbance | MW368483 -near match to type strains of B. odocoilei |
| 2 | 74 | male | I. scapularis, nymph | Yes, positive for Babesia duncani by serology; positive for Babesia by F.I.S.H. | atovaquone- azithromycin, symptoms unresolved; atovaquone-proguanil, symptoms unresolved | -night sweats -chills -fevers -profound fatigue -increased thirst -muscle aches -sleep disturbance | MW368482 -near match to type strains of B. odocoilei |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, J.D.; Sajid, M.S.; Pascoe, E.L.; Foley, J.E. Detection of Babesia odocoilei in Humans with Babesiosis Symptoms. Diagnostics 2021, 11, 947. https://doi.org/10.3390/diagnostics11060947
Scott JD, Sajid MS, Pascoe EL, Foley JE. Detection of Babesia odocoilei in Humans with Babesiosis Symptoms. Diagnostics. 2021; 11(6):947. https://doi.org/10.3390/diagnostics11060947
Chicago/Turabian StyleScott, John D., Muhammad S. Sajid, Emily L. Pascoe, and Janet E. Foley. 2021. "Detection of Babesia odocoilei in Humans with Babesiosis Symptoms" Diagnostics 11, no. 6: 947. https://doi.org/10.3390/diagnostics11060947
APA StyleScott, J. D., Sajid, M. S., Pascoe, E. L., & Foley, J. E. (2021). Detection of Babesia odocoilei in Humans with Babesiosis Symptoms. Diagnostics, 11(6), 947. https://doi.org/10.3390/diagnostics11060947