Genetic Variants in the 3’UTR of BRCA1 and BRCA2 Genes and Their Putative Effects on the microRNA Mechanism in Hereditary Breast and Ovarian Cancer

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Amplification of 3′UTRs BRCA1 and BRCA2 Regions
2.3. Sequencing and Analysis of 3′UTR Regions
2.4. Clinical and Statistical Analysis
3. Results
3.1. HBOC Population
3.2. Hardy-Weinberg Equilibrium (HWE) and Allelic Risk
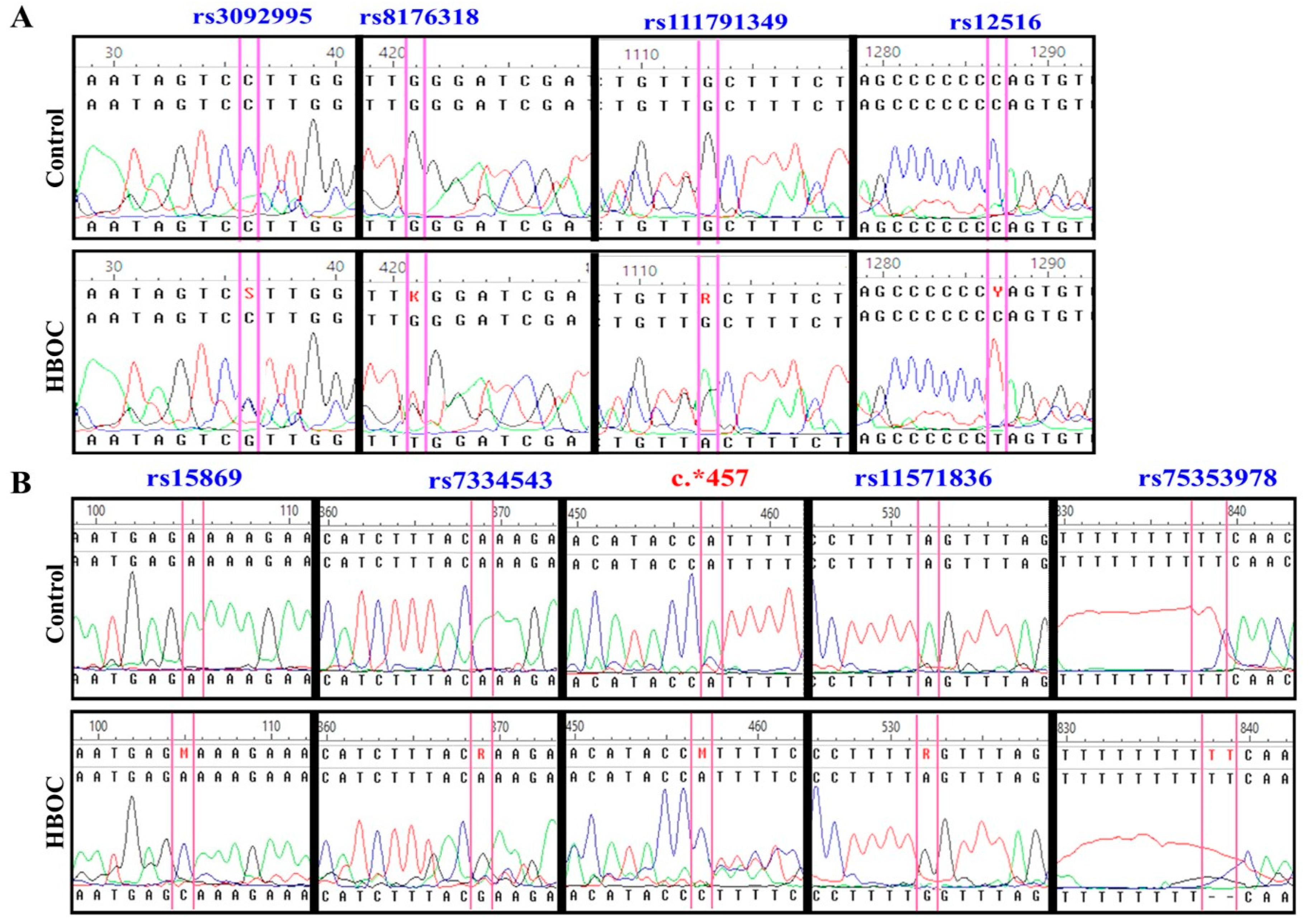
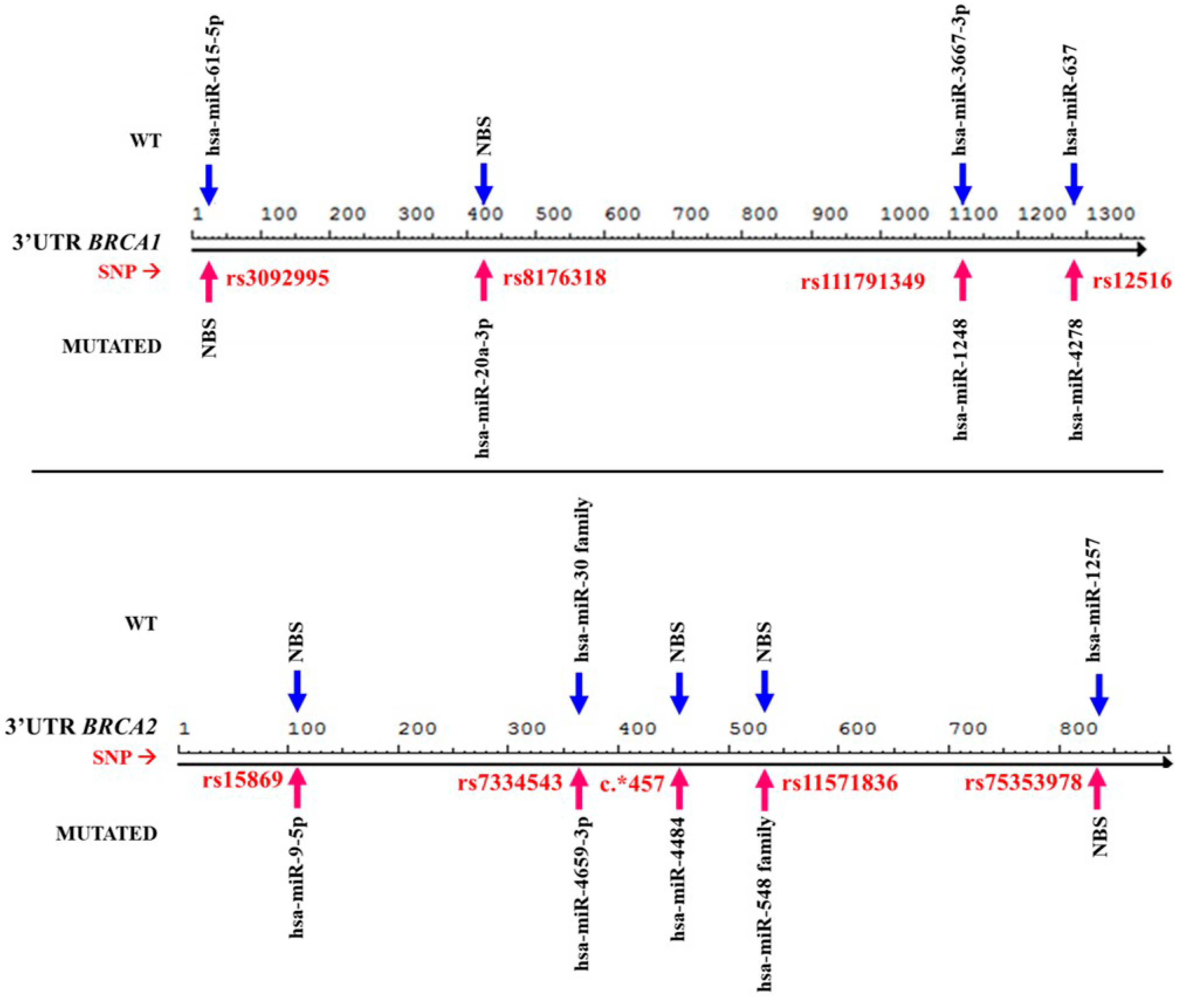
3.3. Presence of Genetic Changes Could Modify miRNA Binding Mechanisms
3.4. Trans-Mechanisms miRNAs Affect Molecular and Cellular Regulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oleg, C.; Bustreo, F. A Month to Remember–Breast Cancer Awareness Month. Promoting Health through the Life-Course 2019. Available online: https://www.who.int/life-course/news/commentaries/breastcancer/en/ (accessed on 6 November 2019).
- Lynch, H.T.; Lynch, J.F.; Lynch, P.M.; Attard, T. Hereditary colorectal cancer syndromes: Molecular genetics, genetic counseling, diagnosis and management. Fam. Cancer 2008, 7, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.C.; McGuffog, L.; Healey, S.; Friedman, E.; Laitman, Y.; Paluch–Shimon, S.; Kaufman, B.; SWE-BRCA, F.; Liljegren, A.; Lindblom, A.; et al. A nonsynonymous polymorphism in IRS1 modifies risk of developing breast and ovarian cancers in BRCA1 and ovarian cancer in BRCA2 mutation carriers. Cancer Epidemiol. Biomark. Prev. 2012, 21, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Pruthi, S.; Gostout, B.S.; Lindor, N.M. Identification and management of women with BRCA mutations or hereditary predisposition for breast and ovarian cancer. Mayo Clin. Proc. 2010, 85, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Ramus, S.J.; Kartsonaki, C.; Gayther, S.A.; Paul, D.P.; Pharoah, O.M.; Beesley, S.J.; Chen, X.; McGuffog, L.; Healey, S.; Couch, F.J.; et al. Genetic variation at 9p22.2 and ovarian cancer risk for BRCA1 and BRCA2 mutation carriers. J. Natl. Cancer Inst. 2011, 103, 105–116. [Google Scholar] [PubMed]
- Moskwa, P.; Buffa, F.M.; Pan, Y.; Panchakshari, R.; Gottipati, P.; Muschel, R.J.; Beech, J.; Kulshrestha, R.; Abdelmohsen, K.; Weinstock, D.M.; et al. miR-182-mediated downregulation of BRCA1 impacts DNA repair and sensitivity to PARP inhibitors. Mol. Cell 2011, 41, 210–220. [Google Scholar] [CrossRef]
- Yang, H.; Jeffrey, P.D.; Miller, J.; Kinnucan, E.; Sun, Y.; Thomä, N.H.; Zheng, N.; Chen, P.-L.; Lee, W.-H.; Pavletich, N.P. BRCA2 function in DNA binding and recombination from a BRCA2-DSS1-ssDNA structure. Science 2002, 297, 1837–1848. [Google Scholar] [CrossRef]
- Chen, K.; Rajewsky, N. Natural selection on human microRNA binding sites inferred from SNP data. Nat. Genet. 2006, 38, 1452–1456. [Google Scholar] [CrossRef]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.-M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef]
- Ha, T.Y. MicroRNAs in human diseases: From autoimmune diseases to skin, psychiatric and neurodegenerative diseases. Immune Netw. 2011, 11, 227–244. [Google Scholar] [CrossRef]
- Calderón-Garcidueñas, A.L.; Ruiz-Flores, P.; Cerda-Flores, R.M.; Barrera-Saldaña, H.A. Clinical follow up of Mexican women with early onset of breast cancer and mutations in the BRCA1 and BRCA2 genes. Salud Publica de Mexico 2005, 47, 110–115. [Google Scholar] [CrossRef]
- Erturk, E.; Cecener, E.; Polatkan, V.; Gokgoz, S.; Egeli, U.; Tunca, B.; Tezcan, G.; Demirdogen, E.; Ak, S.; Tasdelen, I.; et al. Evaluation of genetic variations in miRNA-binding sites of BRCA1 and BRCA2 genes as risk factors for the development of early-onset and/or familial breast cancer. Asian Pac. J. Cancer Prev. 2014, 15, 8319–8324. [Google Scholar] [CrossRef] [PubMed]
- Lheureux, S.; Lambert, B.; Krieger, S.; Legros, A.; Vaur, D.; Denoyelle, C.; Berthet, P.; Poulain, L.; Hardouin, A. Two novel variants in the 3’UTR of the BRCA1 gene in familial breast and/or ovarian cancer. Breast Cancer Res. Treat. 2011, 125, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, C.; Speed, W.C.; Paranjape, T.; Keane, K.; Blitzblau, R.; Hollestelle, A.; Safavi, K.; van den Ouweland, A.; Zelterman, D.; Slack, F.J.; et al. Rare BRCA1 haplotypes including 3’UTR SNPs associated with breast cancer risk. Cell Cycle 2011, 10, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Sole, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Estadística Geografía e Informática (INEGI). Estadísticas a Propósito del Día Mundial Contra El Cáncer (4 de Febrero); Instituto Nacional de Estadística Geografía e Informática, Ed.; Press comunication, INEGI: Aguascalientes, Mexico, 2018. [Google Scholar]
- Vaca-Paniagua, F.; Alvarez-Gomez, R.M.; Fragoso-Ontiveros, V.; Vidal-Millan, S.; Herrera, L.A.; Cantú, D.; Bargallo-Rocha, E.; Mohar, A.; López-Camarillo, C.; Pérez-Plasencia, C. Full-exon pyrosequencing screening of BRCA germline mutations in Mexican women with inherited breast and ovarian cancer. PLoS ONE 2012, 7, e37432. [Google Scholar] [CrossRef]
- Quiyono, M.O.; Colín, S.C.; De La Mora, M.T.G.; Galán, M.D.C.C.; Lladó, D.S. Características epidemiológicas del cáncer de mama y ovario hereditario y del cáncer de mama esporádico en una muestra de la población del ISSSTE. Rev. Esp. Méd. Quir. 2012, 17, 291–299. [Google Scholar]
- Atchley, D.P.; Albarracin, C.T.; Lopez, A.; Valero, V.; Amos, C.I.; Gonzalez-Angulo, A.M.; Hortobagyi, G.N.; Arun, B.K. Clinical and pathologic characteristics of patients with BRCA-positive and BRCA-negative breast cancer. J. Clin. Oncol. 2008, 26, 4282–4288. [Google Scholar] [CrossRef]
- Cao, J.; Luo, C.; Yan, R.; Peng, R.; Wang, K.; Wang, P.; Ye, H.; Song, C.; Oncology, M. rs15869 at miRNA binding site in BRCA2 is associated with breast cancer susceptibility. Med. Oncol. 2016, 33, 1–9. [Google Scholar] [CrossRef]
- Kontorovich, T.; Levy, A.; Korostishevsky, M.; Nir, U.; Friedman, E. Single nucleotide polymorphisms in miRNA binding sites and miRNA genes as breast/ovarian cancer risk modifiers in Jewish high-risk women. Int. J. Cancer 2010, 127, 589–597. [Google Scholar] [CrossRef]
- Brewster, B.L.; Rossiello, F.; French, J.D.; Edwards, S.L.; Wong, M.; Wronski, A.; Whiley, P.; Waddell, N.; Chen, X.; Bove, B.; et al. Identification of fifteen novel germline variants in the BRCA1 3’UTR reveals a variant in a breast cancer case that introduces a functional miR-103 target site. Hum. Mutat. 2012, 33, 1665–1675. [Google Scholar] [CrossRef]
- Garcia, A.I.; Buisson, M.; Damiola, F.; Tessereau, C.; Barjhoux, L.; Verny-Pierre, C.; Sornin, V.; Dondon, M.-G.; Eon-Marchais, S.; GENESIS investigators; et al. Mutation screening of MIR146A/B and BRCA1/2 3′-UTRs in the GENESIS study. Eur. J. Hum. Genet. 2016, 24, 1324–1329. [Google Scholar] [CrossRef] [PubMed]
- Newman, B.; Mu, H.; Butler, L.M.; Millikan, R.C.; Moorman, P.G.; King, M.C. Frequency of breast cancer attributable to BRCA1 in a population-based series of American women. JAMA 1998, 279, 915–921. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Primer Sequences 5′→3′ | Tm (°C) |
|---|---|---|
| BRCA1 | ACCTGATACCCCAGATCCCC | 60.0 |
| ATCCAAGCACTCTCCTTCC | 57.3 | |
| CCTGTGTTCACAAAGGCAGA | 62.0 | |
| TGATCTTGGCTCACTGCAAC | 57.3 | |
| GGCAGGAGAATCACTTCAGC | 57.9 | |
| GCAACAGCTTCCTTCCTGGTGGG | 56.1 | |
| GGAAAATGAAACTAGAAGAGATTT | 58.0 | |
| AGGCTCTGAGAAAGTCGGCT | 60.0 | |
| BRCA2 | GAACAGGAGAGTTCCCAGGC | 56.0 |
| CCCACCTCAGCTTCTCAAAG | 57.3 | |
| GGTGGCTCATGCCTGTAATC | 57.9 | |
| TTGCTCAAAAGGAAACACCA | 56.1 | |
| CAGTTATTTGATGCAGATTCC | 58.0 | |
| AATCAGTGCCAATTTGAAAGCA | 60.0 |
| Clinical Data | >% | Clinical Data | % |
|---|---|---|---|
| Diagnostic age (years) | Mean 37.28 ± 3.30 | Receptor status (ER, PR, HER2/neu) | |
| BMC (kg/m2) | Mean 27.76 ± 5.59 | ER+ PR+ HER2+ | 10% |
| Disease free-survival | ER+ PR+ | 40% | |
| >36 months | 29% | ER+ HER2+ | 2% |
| <36 months No data | 21% 50% | HER2+ | 6% |
| Menarche | 11.77 ± 2.48 | ER+ | 4% |
| Menopause | Triple negative | 34% | |
| Yes | 38% | Therapy | |
| No | 62% | Chemotherapy | |
| Oral-contraceptive use | AC(4)-Taxol(12) | 28% | |
| Yes (>5 years) | 74% | Anthracyclines | 18% |
| No | 20% | Taxanes | 10% |
| Inclusion criteria | Capecitabine | 2% | |
| Family history | 41% | Not especified | 10% |
| Diagnostic >40 years | 59% | No chemotherapy | 32% |
| Births | Radiotherapy | ||
| 0 | 20% | Adjuvant | 76% |
| 1,2 | 54% | Palliative | 2% |
| 3–5 | 20% | Adjuvant/Palliative | 2% |
| >5 | 3% | No radiotherapy | 16% |
| Screening method | Hormone therapy | ||
| Autoexploration | 88% | Yes (Tamoxifen) | 54% |
| Clinical finding | 2% | No hormone therapy | 42% |
| Mastography | 4% | TNM Score | |
| Smoke | I A | 4% | |
| Yes | 78% | I B | 10% |
| No | 22% | II A | 16% |
| Histology | II B | 20% | |
| Infiltrating ductal | 74% | III A | 34% |
| Medullar carcinoma | 4% | III B | 4% |
| Lobullar | 2% | III C | 4% |
| Ductal/Lobullar | 2% | IV | 6% |
| Grade of cell differentiation | Metastasis | ||
| G1 | 6% | Bone | 10% |
| G2 | 28% | Visceral | 2% |
| G3 | 36% | Bone, visceral | 2% |
| Gx | 28% | Bone, visceral, lymph node | 2% |
| Gene | Variant | HBOC Patients | Control | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genotype Frequency | MAF | Genotype Frequency | MAF | |||||||
| BCRA 1 | rs3092995 | C/C | C/G | G/G | G | C/C | C/G | G/G | G | |
| 82% | 0% | 18% | 18% | 76% | 8% | 16% | 20% | |||
| rs8176318 | C/C | C/T | T/T | T | C/C | C/T | T/T | T | ||
| 84% | 10% | 6% | 11% | 90% | 10% | 0% | 5% | |||
| rs111791349 | G/G | G/A | A/A | A | G/G | G/A | A/A | A | ||
| 68% | 12% | 20% | 26% | 76% | 18% | 6% | 15% | |||
| rs12516 | C/C | C/T | T/T | T | C/C | C/T | T/T | T | ||
| 82% | 4% | 14% | 16% | 84% | 16% | 0% | 8% | |||
| BCRA 2 | rs15869 | A/A | A/C | C/C | C | A/A | A/C | C/C | C | |
| 84% | 6% | 10% | 13% | 82% | 18% | 0% | 9% | |||
| rs7334543 | A/A | A/G | G/G | G | A/A | A/G | G/G | G | ||
| 94% | 0% | 6% | 6% | 94% | 0% | 6% | 6% | |||
| c.*457 | A/A | A/C | C/C | C | A/A | A/C | C/C | C | ||
| 66% | 12% | 22% | 28% | 68% | 30% | 2% | 17% | |||
| rs11571836 | A/A | A/G | G/G | G | A/A | A/G | G/G | G | ||
| 82% | 4% | 14% | 16% | 82% | 14% | 40% | 11% | |||
| rs75353978 ** | TT/TT | delTT | TT/TT | delTT | ||||||
| 32% | 68% | 32% | 92% | 8% | 8% | |||||
| Gene | Variant/Alleles | Risk Allele 1 | Risk Allele 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Allele Freq. Difference | Heterozygous | Homozygous | Allele Positivity | Allele Freq. Difference | Heterozygous | Homozygous | Allele Positivity | |||
| [1]↔[2] | [11]↔[12] | [11+]↔[22] | [11]↔[12+22] | [2]↔[1] | [22]↔[12] | [22]↔[11] | [11+12]↔[22] | |||
| BRCA1 | rs8176318 (C/T) | 2.4 (0.8–7.0) | 1.1 (0.3–4.0) | 7.5 (0.4–149.4) | 1.7(0.5–5.7) | 0.4 (0.1–1.3) | 0.1 (0.01–3.5) | 0.1 (0.01–2.7) | 0.1 (0.01–2.7) | |
| 0.118 (P) | 0.918 | 0.078 | 0.372 | 0.118 (P) | 0.1185 | 0.078 | 0.079 | |||
| rs12516 (C/T) | 2.2 (0.9–5.4) | 0.3 (0.05–1.3) | 15.4 (0.9–277.7) | 1.2 (0.4–3.3) | 0.5 (0.2–1.1) | 0.02 (0.001–0.5) | 0.1 (0.004–1.2) | 0.1 (0.003–1.0) | ||
| 0.082 (P) | 0.078 | 0.001 | 0.79 | 0.082 (P) | 0.001 | 0.0099 | 0.006 | |||
| BRCA2 | rs15869 (A/C) | 1.5 (0.6–3.7) | 0.3 (0.08–1.3) | 10.7 (0.6–200.5) | 0.9 (0.3–2.5) | 0.7 (0.3–1.6) | 0.03 (0.001–0.8) | 0.1 (0.005–1.7) | 0.1 (0.004–1.51) | |
| 0.366 (P) | 0.097 | 0.032 | 0.79 | 0.366 (P) | 0.005 | 0.032 | 0.0218 | |||
| c.*457 (A/C) | 1.9 (0.96–3.8) | 0.4 (0.1–1.2) | 11.3 (1.4–92.8) | 1.1 (0.5–2.5) | 0.5 (0.3–1.0) | 0.04 (0.004–0.4) | 0.1 (0.01–0.7) | 0.1 (0.01–0.6) | ||
| 0.063 (P) | 0.096 | 0.007 | 0.832 | 0.063 (P) | 0.0005 | 0.007 | 0.0021 | |||
| Gene | Variant | microRNA | ΔΔG (Kcal/mol) | Duplex SNP-miRNA | Effect | |
|---|---|---|---|---|---|---|
| BRCA1 | rs8176318 (G/T) | hsa-miR-20a-3p | Wt: 0.00 | miRNA: | 3′-gaaauuCACGAGU-AUUACGUCa-5′ | Created |
| |||:||| |||||| | ||||||
| SNP: −22.70 | UTR: | 5′-aaccctGTGTTCACAAATGCAGa-3′ | ||||
| rs12516 (C/T) | hsa-miR-4278 | Wt: −18.30 | miRNA: | 3′-guucccGUUUGGGGGAUc-5′ | Created | |
| :|: ||||||| | ||||||
| SNP: −21.60 | UTR: | 5′-cttcccTAGCCCCCCTAg-3′ | ||||
| hsa-miR-637 | Wt: −28.30 | miRNA: | 3′-ugcgucucGGGCUUUCGGGGGUCa-5′ | Eliminated | ||
| ||| |: ||||X|| | ||||||
| SNP: −16.00 | UTR: | 5′-taggtcttCCCTAGCCCCCCTAGt-3′ | ||||
| BRCA2 | rs15869 (A/C) | hsa-miR-9-5p | Wt: −14.00 | miRNA: | 3′-ucuuugguuaucUAGCUGUAUga-5′ | Created |
| || :|| | | ||||||
| SNP: −23.20 | UTR: | 5′-ttatgttgcacaATGAGCAAAga-3′ | ||||
| c.* 457 (A/C) | hsa-miR-4484 | Wt: −11.10 | miRNA: | 3′-ACCCCGAAGAGGGCGGAAAA-5′ | Created | |
| :| |||||| | ||||||
| SNP: −23.70 | UTR: | 5′-TGAAATAAACATACCCTTTT-3′ | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Chaparro, M.M.; Garza-Veloz, I.; Zayas-Villanueva, O.A.; Martinez-Fierro, M.L.; Delgado-Enciso, I.; Gomez-Govea, M.A.; Martínez-de-Villarreal, L.E.; Reséndez-Pérez, D.; Rodríguez-Sánchez, I.P. Genetic Variants in the 3’UTR of BRCA1 and BRCA2 Genes and Their Putative Effects on the microRNA Mechanism in Hereditary Breast and Ovarian Cancer. Diagnostics 2020, 10, 298. https://doi.org/10.3390/diagnostics10050298
Sánchez-Chaparro MM, Garza-Veloz I, Zayas-Villanueva OA, Martinez-Fierro ML, Delgado-Enciso I, Gomez-Govea MA, Martínez-de-Villarreal LE, Reséndez-Pérez D, Rodríguez-Sánchez IP. Genetic Variants in the 3’UTR of BRCA1 and BRCA2 Genes and Their Putative Effects on the microRNA Mechanism in Hereditary Breast and Ovarian Cancer. Diagnostics. 2020; 10(5):298. https://doi.org/10.3390/diagnostics10050298
Chicago/Turabian StyleSánchez-Chaparro, María Marisela, Idalia Garza-Veloz, Omar Alejandro Zayas-Villanueva, Margarita L. Martinez-Fierro, Iván Delgado-Enciso, Mayra Alejandra Gomez-Govea, Laura Elia Martínez-de-Villarreal, Diana Reséndez-Pérez, and Iram Pablo Rodríguez-Sánchez. 2020. "Genetic Variants in the 3’UTR of BRCA1 and BRCA2 Genes and Their Putative Effects on the microRNA Mechanism in Hereditary Breast and Ovarian Cancer" Diagnostics 10, no. 5: 298. https://doi.org/10.3390/diagnostics10050298
APA StyleSánchez-Chaparro, M. M., Garza-Veloz, I., Zayas-Villanueva, O. A., Martinez-Fierro, M. L., Delgado-Enciso, I., Gomez-Govea, M. A., Martínez-de-Villarreal, L. E., Reséndez-Pérez, D., & Rodríguez-Sánchez, I. P. (2020). Genetic Variants in the 3’UTR of BRCA1 and BRCA2 Genes and Their Putative Effects on the microRNA Mechanism in Hereditary Breast and Ovarian Cancer. Diagnostics, 10(5), 298. https://doi.org/10.3390/diagnostics10050298