Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review

Abstract
1. Introduction
2. Methods
2.1. Data Sources and Eligibility Criteria
2.2. Search Strategy
2.3. Study Selection
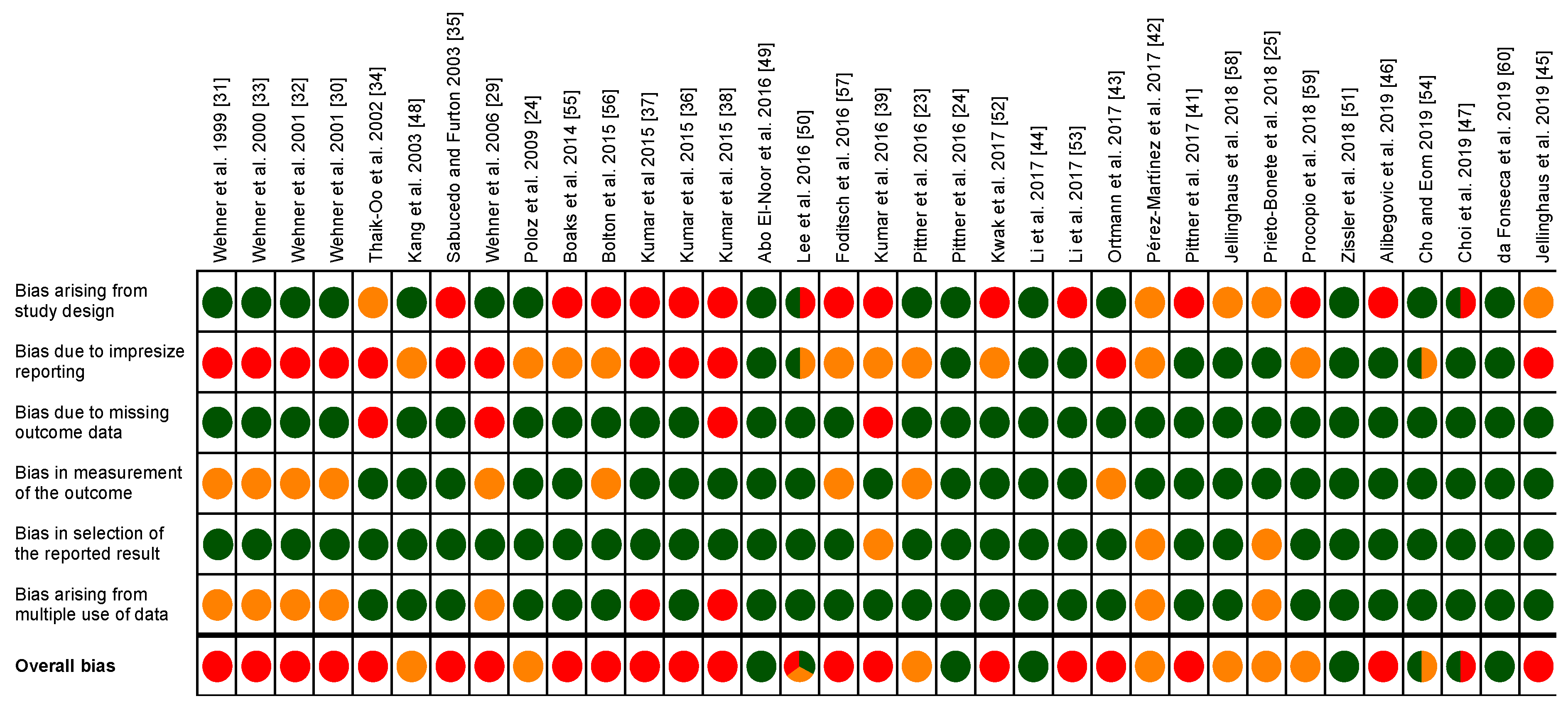
2.4. Risk of Bias Assessment
2.5. Data Extraction and Synthesis
3. Results
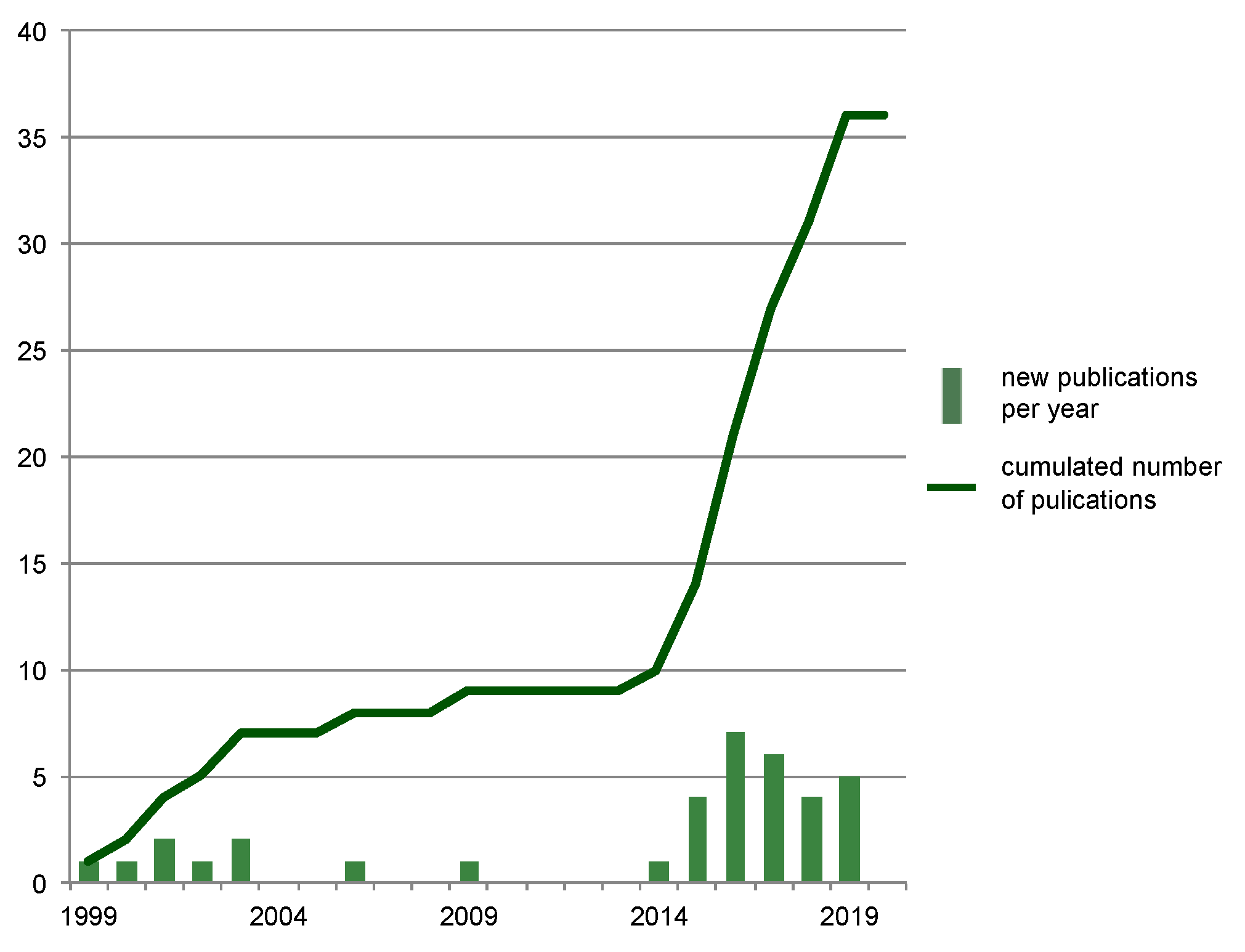
3.1. Study Selection
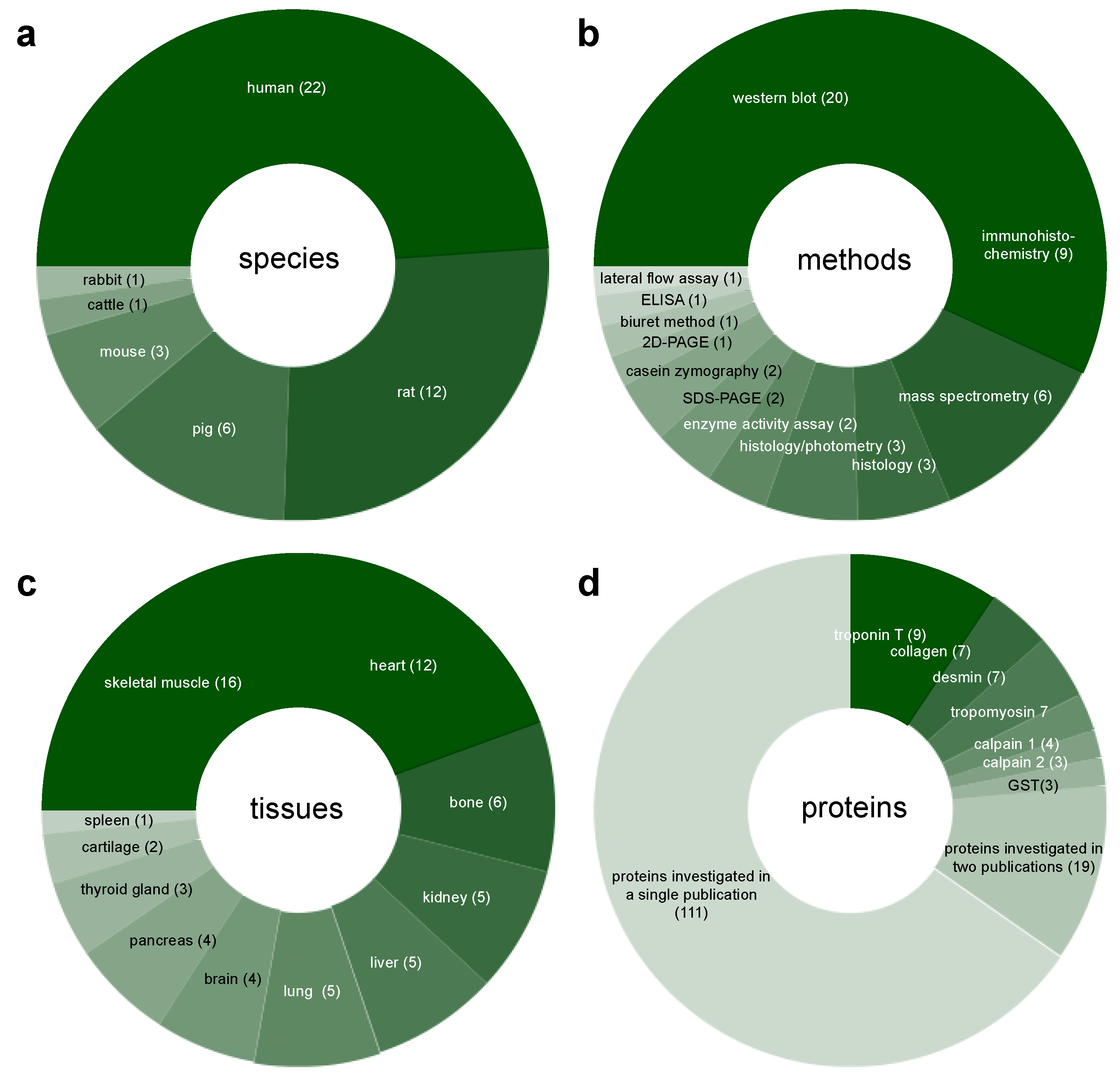
3.2. Study Characteristics
3.3. Risk of Bias Assessment
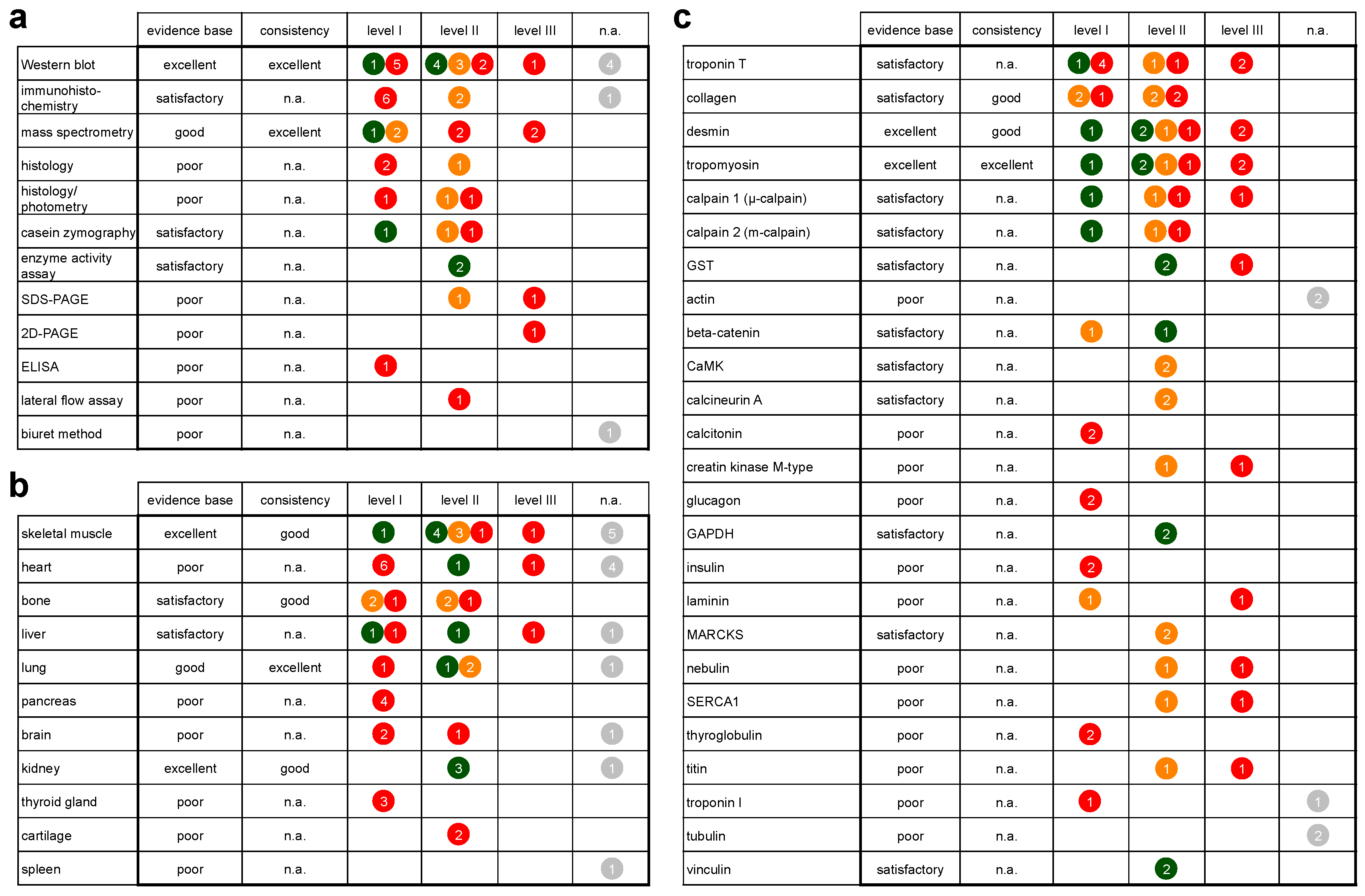
3.4. Protein Degradation as a Tool for Postmortem Interval (PMI) Estimation
4. Discussion
4.1. Body of Evidence
4.1.1. Evidence Base and Consistency
4.1.2. Generalizability
4.2. Methods
4.2.1. Sample Size
4.2.2. The Use of Control Samples
4.2.3. Animal Models versus Human Corpses
4.2.4. Protein Identification
4.2.5. Data Assessment and Analysis
4.2.6. Applicability of Methods for Forensic Case Work and Future Investigation
4.3. Tissues
4.3.1. Rate of Tissue Decomposition
4.3.2. Sampling Site
4.3.3. Applicability of Tissues in Forensic Case Work and Future Investigation
4.4. Proteins
4.4.1. Rate and Pattern of Protein Degradation
4.4.2. Degradation Kinetics of Individual Proteins
4.4.3. Postmortem Increase of Proteins
4.4.4. Applicability of Proteins in Forensic Case Work and Future Investigations
5. Limitations of the Present Review
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Madea, B. Methods for determining time of death. Forensic Sci. Med. Pathol. 2016, 12, 451–485. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Agrawal, Y.K. An overview of methods used for estimation of time since death. Aust. J. Forensic Sci. 2011, 43, 275–285. [Google Scholar] [CrossRef]
- Donaldson, A.E.; Lamont, I.L. Biochemistry Changes That Occur after Death: Potential Markers for Determining Post-Mortem Interval. PLoS ONE 2013, 8, e82011. [Google Scholar] [CrossRef] [PubMed]
- Henssge, C.; Knight, B.; Krompecher, T.H.; Madea, B.; Nokes, L. The Estimation of the Time since Death in the Early Post-Mortem Period; Eward Arnold: London, UK, 1995. [Google Scholar]
- Megyesi, M.S.; Nawrocki, S.P.; Haskell, N.H. Using accumulated degree-days to estimate the postmortem interval from decomposed human remains. J. Forensic Sci. 2005, 50, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, A.E.; Lamont, I.L. Estimation of post-mortem interval using biochemical markers. Aust. J. Forensic Sci. 2014, 46, 8–26. [Google Scholar] [CrossRef]
- Swain, R.; Kumar, A.; Sahoo, J.; Lakshmy, R.; Gupta, S.K.; Bhardwaj, D.N.; Pandey, R.M. Estimation of post-mortem interval: A comparison between cerebrospinal fluid and vitreous humour chemistry. J. Forensic Leg. Med. 2015, 36, 144–148. [Google Scholar] [CrossRef]
- Chen, J.-H.; Inamori-Kawamoto, O.; Michiue, T.; Ikeda, S.; Ishikawa, T.; Maeda, H. Cardiac biomarkers in blood, and pericardial and cerebrospinal fluids of forensic autopsy cases: A reassessment with special regard to postmortem interval. Leg. Med. 2015, 17, 343–350. [Google Scholar] [CrossRef]
- Tumram, N.K.; Bardale, R.V.; Dongre, A.P. Postmortem analysis of synovial fluid and vitreous humour for determination of death interval: A comparative study. Forensic Sci. Int. 2011, 204, 186–190. [Google Scholar] [CrossRef]
- Kikuchi, K.; Kawahara, K.-I.; Biswas, K.K.; Ito, T.; Tancharoen, S.; Shiomi, N.; Koda, Y.; Matsuda, F.; Morimoto, Y.; Oyama, Y.; et al. HMGB1: A new marker for estimation of the postmortem interval. Exp. Ther. Med. 2010, 1, 109–111. [Google Scholar] [CrossRef]
- Kumar, S.; Verma, A.K. Estimation of postmortem interval using the data of insulin level in the cadaver’s blood. Data Brief. 2016, 7, 354–356. [Google Scholar] [CrossRef]
- Costa, I.; Carvalho, F.; Magalhães, T.; Pinho, P.G.; Silvestre, R.; Dinis-Oliveira, R.J. Promising blood-derived biomarkers for estimation of the postmortem interval. Toxicol. Res. 2015, 4, 1443–1452. [Google Scholar] [CrossRef]
- Peregrín-Alvarez, J.M.; Sanford, C.; Parkinson, J. The conservation and evolutionary modularity of metabolism. Genome Biol. 2009, 10, R63. [Google Scholar] [CrossRef] [PubMed]
- Varshavsky, A. Proteolysis. In Encyclopedia of Genetics; Brenner, S., Miller, J.H., Eds.; Academic Press: New York, NY, USA, 2001; pp. 1573–1575. ISBN 978-0-12-227080-2. [Google Scholar]
- Lecker, S.H.; Goldberg, A.L.; Mitch, W.E. Protein Degradation by the Ubiquitin–Proteasome Pathway in Normal and Disease States. J. Am. Soc. Nephrol. 2006, 17, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Wendt, A.; Thompson, V.F.; Goll, D.E. Interaction of calpastatin with calpain: A review. Biol. Chem. 2004, 385, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.-R.; Takahashi, K. Changes in concentration of sarcoplasmic free calcium during post-mortem ageing of meat. Meat Sci. 2006, 73, 395–403. [Google Scholar] [CrossRef]
- Lonergan, E. The Relationship between the Calpain Enzyme System and the Postmortem Degradation of Selected Myofibrillar Proteins. Ph.D. Thesis, Iowa State Universit, Ames, IA, USA, 1995. [Google Scholar]
- Sorimachi, Y.; Harada, K.; Yoshida, K. Involvement of calpain in postmortem proteolysis in the rat brain. Forensic Sci. Int. 1996, 81, 165–174. [Google Scholar] [CrossRef]
- Aufderheide, A.C. The Scientific Study of Mummies; Cambridge University Press: Cambridge, UK, 2003; ISBN 978-0-521-81826-1. [Google Scholar]
- Pittner, S.; Monticelli, F.C.; Pfisterer, A.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem degradation of skeletal muscle proteins: A novel approach to determine the time since death. Int. J. Legal Med. 2016, 130, 421–431. [Google Scholar] [CrossRef]
- Poloz, Y.O.; O’Day, D.H. Determining time of death: Temperature-dependent postmortem changes in calcineurin A, MARCKS, CaMKII, and protein phosphatase 2A in mouse. Int. J. Legal Med. 2009, 123, 305–314. [Google Scholar] [CrossRef]
- Prieto-Bonete, G.; Pérez-Cárceles, M.D.; Maurandi-López, A.; Pérez-Martínez, C.; Luna, A. Association between protein profile and postmortem interval in human bone remains. J. Proteomics 2018. [Google Scholar] [CrossRef]
- Higgins, J.; Sterne, J.; Savović, J.; Page, M.; Hróbjartsson, A.; Boutron, I.; Reeves, B.; Eldridge, S. A revised tool for assessing risk of bias in randomized trials. Cochrane Database Syst. Rev. 2016, 10 (Suppl.1). [Google Scholar] [CrossRef]
- Li, C.; Wang, Q.; Zhang, Y.; Lin, H.; Zhang, J.; Huang, P.; Wang, Z. Research progress in the estimation of the postmortem interval by Chinese forensic scholars. Forensic Sci. Res. 2016, 1, 3–13. [Google Scholar] [CrossRef] [PubMed]
- National Health and Medical Research Council (NHMRC). Levels of Evidence and Grades for Recommendations for Guideline Developers. 2009. Available online: https://www.nhmrc.gov.au/_files_nhmrc/file/guidelines/developers/nhmrc_levels_grades_evidence_120423.pdf (accessed on 6 February 2020).
- Wehner, F.; Steinriede, A.; Martin, D.; Wehner, H.-D. Two-tailed delimitation of the time of death by immunohistochemical detection of somatostatin and GFAP. Forensic Sci. Med. Pathol. 2006, 2, 241–247. [Google Scholar] [CrossRef]
- Wehner, F.; Wehner, H.-D.; Subke, J. Delimitation of the time of death by immunohistochemical detection of glucagon in pancreatic α-cells. Forensic Sci. Int. 2001, 124, 192–199. [Google Scholar] [CrossRef]
- Wehner, F.; Wehner, H.-D.; Schieffer, M.C.; Subke, J. Delimitation of the time of death by immunohistochemical detection of insulin in pancreatic β-cells. Forensic Sci. Int. 1999, 105, 161–169. [Google Scholar] [CrossRef]
- Wehner, F.; Wehner, H.-D.; Subke, J. Delimitation of the time of death by immunohistochemical detection of calcitonin. Forensic Sci. Int. 2001, 122, 89–94. [Google Scholar] [CrossRef]
- Wehner, F.; Wehner, H.-D.; Schieffer, M.C.; Subke, J. Delimitation of the time of death by immunohistochemical detection of thyroglobulin. Forensic Sci. Int. 2000, 110, 199–206. [Google Scholar] [CrossRef]
- Thaik-Oo, M.; Tanaka, E.; Tsuchiya, T.; Kominato, Y.; Honda, K.; Yamazaki, K.; Misawa, S. Estimation of postmortem interval from hypoxic inducible levels of vascular endothelial growth factor. J. Forensic Sci. 2002, 47, 186–189. [Google Scholar] [CrossRef]
- Sabucedo, A.J.; Furton, K.G. Estimation of postmortem interval using the protein marker cardiac Troponin I. Forensic Sci. Int. 2003, 134, 11–16. [Google Scholar] [CrossRef]
- Kumar, S.; Ali, W.; Bhattacharya, S.; Singh, U.S.; Kumar, A.; Verma, A.K. The effect of elapsed time on cardiac troponin-T (cTnT) degradation and its relation to postmortem interval in cases of electrocution. J. Forensic Leg. Med. 2015, 34, 45–49. [Google Scholar] [CrossRef]
- Kumar, S.; Ali, W.; Singh, U.S.; Kumar, A.; Bhattacharya, S.; Verma, A.K. The effect of elapsed time on the cardiac Troponin-T (cTnT) proteolysis in case of death due to burn: A study to evaluate the potential forensic use of cTnT to determine the postmortem interval. Sci. Justice 2015, 55, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ali, W.; Singh, U.S.; Verma, A.K.; Bhattacharya, S.; Kumar, A.; Singh, R.; Rupani, R. Time-dependent Degradation Pattern of Cardiac Troponin T in Cases of Death by Burn. West. Indian Med. J. 2015, 64, 226–229. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kumar, S.; Ali, W.; Singh, U.S.; Kumar, A.; Bhattacharya, S.; Verma, A.K.; Rupani, R. Temperature-Dependent Postmortem Changes in Human Cardiac Troponin-T (cTnT): An Approach in Estimation of Time Since Death. J. Forensic Sci. 2016, 61, S241–S245. [Google Scholar] [CrossRef] [PubMed]
- Pittner, S.; Ehrenfellner, B.; Monticelli, F.C.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem muscle protein degradation in humans as a tool for PMI delimitation. Int. J. Legal Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Pittner, S.; Ehrenfellner, B.; Zissler, A.; Racher, V.; Trutschnig, W.; Bathke, A.C.; Sänger, A.M.; Stoiber, W.; Steinbacher, P.; Monticelli, F.C. First application of a protein-based approach for time since death estimation. Int. J. Legal Med. 2017, 131, 479–483. [Google Scholar] [CrossRef]
- Pérez-Martínez, C.; Pérez-Cárceles, M.D.; Legaz, I.; Prieto-Bonete, G.; Luna, A. Quantification of nitrogenous bases, DNA and Collagen type I for the estimation of the postmortem interval in bone remains. Forensic Sci. Int. 2017, 281, 106–112. [Google Scholar] [CrossRef]
- Ortmann, J.; Doberentz, E.; Madea, B. Immunohistochemical methods as an aid in estimating the time since death. Forensic Sci. Int. 2017, 273, 71–79. [Google Scholar] [CrossRef]
- Li, C.; Li, Z.; Tuo, Y.; Ma, D.; Shi, Y.; Zhang, Q.; Zhuo, X.; Deng, K.; Chen, Y.; Wang, Z.; et al. MALDI-TOF MS as a Novel Tool for the Estimation of Postmortem Interval in Liver Tissue Samples. Sci. Rep. 2017, 7, 4887. [Google Scholar] [CrossRef]
- Jellinghaus, K.; Urban, P.K.; Hachmann, C.; Bohnert, M.; Hotz, G.; Rosendahl, W.; Wittwer-Backofen, U. Collagen degradation as a possibility to determine the post-mortem interval (PMI) of human bones in a forensic context—A survey. Leg. Med. 2019, 36, 96–102. [Google Scholar] [CrossRef]
- Alibegović, A.; Blagus, R.; Martinez, I.Z. Safranin O without fast green is the best staining method for testing the degradation of macromolecules in a cartilage extracellular matrix for the determination of the postmortem interval. Forensic Sci. Med. Pathol. 2019. [Google Scholar] [CrossRef]
- Choi, K.-M.; Zissler, A.; Kim, E.; Ehrenfellner, B.; Cho, E.; Lee, S.; Steinbacher, P.; Yun, K.N.; Shin, J.H.; Kim, J.Y.; et al. Postmortem proteomics to discover biomarkers for forensic PMI estimation. Int. J. Legal Med. 2019, 133, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Kassam, N.; Gauthier, M.L.; O’Day, D.H. Post-mortem changes in calmodulin binding proteins in muscle and lung. Forensic Sci. Int. 2003, 131, 140–147. [Google Scholar] [CrossRef]
- Abo El-Noor, M.M.; Elhosary, N.M.; Khedr, N.F.; El-Desouky, K.I. Estimation of Early Postmortem Interval Through Biochemical and Pathological Changes in Rat Heart and Kidney. Am. J. Forensic Med. Pathol. 2016, 37, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-G.; Yang, K.E.; Hwang, J.W.; Kang, H.-S.; Lee, S.-Y.; Choi, S.; Shin, J.; Jang, I.-S.; An, H.J.; Chung, H.; et al. Degradation of Kidney and Psoas Muscle Proteins as Indicators of Post-Mortem Interval in a Rat Model, with Use of Lateral Flow Technology. PLoS ONE 2016, 11, e0160557. [Google Scholar] [CrossRef] [PubMed]
- Zissler, A.; Ehrenfellner, B.; Foditsch, E.E.; Monticelli, F.C.; Pittner, S. Does altered protein metabolism interfere with postmortem degradation analysis for PMI estimation? Int. J. Legal Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.-H.; Kim, H.K.; Kim, K.; Noh, B.R.; Cheon, H.I.; Yeo, M.; Shakya, R.; Shrestha, S.A.; Kim, D.; Choe, S.; et al. Proteomic Evaluation of Biomarkers to Determine the Postmortem Interval. Anal. Lett. 2017, 50, 207–218. [Google Scholar] [CrossRef]
- Li, C.; Ma, D.; Deng, K.; Chen, Y.; Huang, P.; Wang, Z. Application of MALDI-TOF MS for Estimating the Postmortem Interval in Rat Muscle Samples. J. Forensic Sci. 2017, 62, 1345–1350. [Google Scholar] [CrossRef]
- Cho, H.-W.; Eom, Y.-B. Potential Forensic Application of Receptor for Advanced Glycation End Products (RAGE) as a Novel Biomarker for Estimating Postmortem Interval. J. Forensic Sci. 2019, 64, 1878–1883. [Google Scholar] [CrossRef]
- Boaks, A.; Siwek, D.; Mortazavi, F. The temporal degradation of bone collagen: A histochemical approach. Forensic Sci. Int. 2014, 240, 104–110. [Google Scholar] [CrossRef]
- Bolton, S.N.; Whitehead, M.P.; Dudhia, J.; Baldwin, T.C.; Sutton, R. Investigating the Postmortem Molecular Biology of Cartilage and its Potential Forensic Applications. J. Forensic Sci. 2015, 60, 1061–1067. [Google Scholar] [CrossRef]
- Foditsch, E.E.; Saenger, A.M.; Monticelli, F.C. Skeletal muscle proteins: A new approach to delimitate the time since death. Int. J. Legal Med. 2016, 130, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Jellinghaus, K.; Hachmann, C.; Höland, K.; Bohnert, M.; Wittwer-Backofen, U. Collagen degradation as a possibility to determine the post-mortem interval (PMI) of animal bones: A validation study referring to an original study of Boaks et al. (2014). Int. J. Legal Med. 2018, 132, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Procopio, N.; Williams, A.; Chamberlain, A.T.; Buckley, M. Forensic proteomics for the evaluation of the post-mortem decay in bones. J. Proteom. 2018, 177, 21–30. [Google Scholar] [CrossRef] [PubMed]
- da Fonseca, C.A.R.; Paltian, J.; dos Reis, A.S.; Bortolatto, C.F.; Wilhelm, E.A.; Luchese, C. Na+/K+-ATPase, acetylcholinesterase and glutathione S-transferase activities as new markers of postmortem interval in Swiss mice. Leg. Med. 2019, 36, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Procopio, N.; Chamberlain, A.T.; Buckley, M. Exploring Biological and Geological Age-related Changes through Variations in Intra- and Intertooth Proteomes of Ancient Dentine. J. Proteome Res. 2018, 17, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Fucci, N.; Campobasso, C.P.; Mastrogiuseppe, L.; Puccinelli, C.; Marcheggiani, S.; Mancini, L.; Marino, L.; Pascali, V.L. Diatoms in drowning cases in forensic veterinary context: A preliminary study. Int. J. Legal Med. 2017. [Google Scholar] [CrossRef]
- Brooks, J.W. Postmortem Changes in Animal Carcasses and Estimation of the Postmortem Interval. Vet. Pathol. 2016, 53, 929–940. [Google Scholar] [CrossRef]
- Wescott, D.J. Recent advances in forensic anthropology: Decomposition research. Forensic Sci. Res. 2018, 3, 278–293. [Google Scholar] [CrossRef]
- Ehrenfellner, B.; Zissler, A.; Steinbacher, P.; Monticelli, F.C.; Pittner, S. Are animal models predictive for human postmortem muscle protein degradation? Int. J. Legal Med. 2017, 131, 1615–1621. [Google Scholar] [CrossRef]
- Janes, K.A. An analysis of critical factors for quantitative immunoblotting. Sci. Signal. 2015, 8, rs2. [Google Scholar] [CrossRef]
- Dent, B.B.; Forbes, S.L.; Stuart, B.H. Review of human decomposition processes in soil. Environ. Geol. 2004, 45, 576–585. [Google Scholar] [CrossRef]
- Forensic Taphonomy: The Postmortem Fate of Human Remains, 1st ed.; Sorg, M.H., Haglund, W.D., Eds.; CRC Press: Boca Raton, FL, USA, 1997; ISBN 978-0-8493-9434-8. [Google Scholar]
- Benbow, M.E.; Tomberlin, J.K.; Tarone, A.M. Carrion Ecology, Evolution, and Their Applications; CRC Press: Boca Raton, FL, USA, 2015; ISBN 978-1-4665-7547-9. [Google Scholar]
- Dettmeyer, R.; Verhoff, M.A.; Schütz, H.F. Forensic Medicine: Fundamentals and Perspectives; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-38817-0. [Google Scholar]
- Cocariu, E.A.; Mageriu, V.; Stăniceanu, F.; Bastian, A.; Socoliuc, C.; Zurac, S. Correlations Between the Autolytic Changes and Postmortem Interval in Refrigerated Cadavers. Rom. J. Intern. Med. 2016, 54, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.; Henckel, P.; Purslow, P.P. Effect of muscle type on the rate of post-mortem proteolysis in pigs. Meat Sci. 2004, 66, 595–601. [Google Scholar] [CrossRef]
- Tomaszewska-Gras, J.; Schreurs, F.J.G.; Kijowski, J. Post mortem development of meat quality as related to changes in cytoskeletal proteins of chicken muscles. Br. Poult. Sci. 2011, 52, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Marino, R.; della Malva, A.; Albenzio, M. Proteolytic changes of myofibrillar proteins in Podolian meat during aging: Focusing on tenderness. J. Anim. Sci. 2015, 93, 1376–1387. [Google Scholar] [CrossRef]
- Pittner, S.; Gotsmy, W.; Zissler, A.; Ehrenfellner, B.; Baumgartner, D.; Schrüfer, A.; Steinbacher, P.; Monticelli, F. Intra- and intermuscular variations of postmortem protein degradation for PMI estimation. Int. J. Legal Med. 2020, 134, 1775–1782. [Google Scholar] [CrossRef]
- Fountoulakis, M.; Hardmeier, R.; Höger, H.; Lubec, G. Postmortem Changes in the Level of Brain Proteins. Exp. Neurol. 2001, 167, 86–94. [Google Scholar] [CrossRef]
- Chandana, R.; Mythri, R.B.; Mahadevan, A.; Shankar, S.K.; Bharath, M.M.S. Biochemical analysis of protein stability in human brain collected at different post-mortem intervals. Indian J. Med. Res. 2009, 129, 189–199. [Google Scholar]
- Geesink, G.H.; Koohmaraie, M. Postmortem proteolysis and calpain/calpastatin activity in callipyge and normal lamb biceps femoris during extended postmortem storage. J. Anim. Sci. 1999, 77, 1490–1501. [Google Scholar] [CrossRef]
- Hwan, S.-F.; Bandman, E. Studies of Desmin and α-Actinin Degradation in Bovine Semitendinosus Muscle. J. Food Sci. 1989, 54, 1426–1430. [Google Scholar] [CrossRef]
- Ferrer, I.; Santpere, G.; Arzberger, T.; Bell, J.; Blanco, R.; Boluda, S.; Budka, H.; Carmona, M.; Giaccone, G.; Krebs, B.; et al. Brain Protein Preservation Largely Depends on the Postmortem Storage Temperature: Implications for Study of Proteins in Human Neurologic Diseases and Management of Brain Banks: A BrainNet Europe Study. J. Neuropathol. Exp. Neurol. 2007, 66, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.A.; Wang, C.; Hernandez, D.; Siedlak, S.L.; Rodgers, M.S.; Achar, R.K.; Fahmy, L.M.; Torres, S.L.; Petersen, R.B.; Zhu, X.; et al. Individual Case Analysis of Postmortem Interval Time on Brain Tissue Preservation. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Magliery, T.J. Protein stability: Computation, sequence statistics, and new experimental methods. Curr. Opin. Struct. Biol. 2015, 33, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, M.; Du, M.; Shen, Q.W.; Zhang, D. Dephosphorylation enhances postmortem degradation of myofibrillar proteins. Food Chem. 2018, 245, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Falahati, A.; Sharkey, S.W.; Christensen, D.; McCoy, M.; Miller, E.A.; Murakami, M.A.; Apple, F.S. Implementation of serum cardiac troponin I as marker for detection of acute myocardial infarction. Am. Heart J. 1999, 137, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.Y.; Stromer, M.H.; Robson, R.M. Identification of the 30 kDa polypeptide in post mortem skeletal muscle as a degradation product of troponin-T. Biochimie 1994, 76, 369–375. [Google Scholar] [CrossRef]
- Koohmaraie, M. The role of Ca(2+)-dependent proteases (calpains) in post mortem proteolysis and meat tenderness. Biochimie 1992, 74, 239–245. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Y.; Liang, R.; Ren, L.; Zhu, H.; Li, K.; Zhu, L.; Luo, X. Effect of Rapid Chilling on Beef Quality and Cytoskeletal Protein Degradation in M. longissimus of Chinese Yellow Crossbred Bulls. Asian-Australas. J. Anim. Sci. 2012, 25, 1197–1204. [Google Scholar] [CrossRef][Green Version]
- Zhao, L.; Xing, T.; Huang, J.; Qiao, Y.; Chen, Y.; Huang, M. Involvement of μ/m-calpain in the proteolysis and meat quality changes during postmortem storage of chicken breast muscle. Anim. Sci. J. 2018, 89, 423–431. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Chou, R.-G.R. Postmortem role of calpains in Pekin duck skeletal muscles. J. Sci. Food Agric. 2012, 92, 1620–1626. [Google Scholar] [CrossRef]
- Amin, H.A.A.; El-Hennawy, A.M.Y.; Nakhla, G.A.A.; Tabak, S.A.-H.; Hassan, H.H. Immuno-histochemistry in the detection of early myocardial infarction (a post-mortem study). Egypt. J. Forensic Sci. 2011, 1, 5–12. [Google Scholar] [CrossRef][Green Version]
- Fishbein, M.C.; Wang, T.; Matijasevic, M.; Hong, L.; Apple, F.S. Myocardial tissue troponins T and I: An immunohistochemical study in experimental models of myocardial ischemia. Cardiovasc. Pathol. 2003, 12, 65–71. [Google Scholar] [CrossRef]
- Ricchiuti, V.; Zhang, J.; Apple, F.S. Cardiac troponin I and T alterations in hearts with severe left ventricular remodeling. Clin. Chem. 1997, 43, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Gill-King, H. Chemical and ultrastructural aspects of decomposition. In Forensic Taphonomy: The Postmortem Fate of Human Remains; Haglund, W.D., Sorg, M.H., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 93–108. [Google Scholar]
- Contreras-Castillo, C.J.; Lomiwes, D.; Wu, G.; Frost, D.; Farouk, M.M. The effect of electrical stimulation on post mortem myofibrillar protein degradation and small heat shock protein kinetics in bull beef. Meat Sci. 2016, 113, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Wojtysiak, D.; Połtowicz, K. Effect of Ageing Time on Microstructure, Rate of Desmin Degradation and Meat Quality of Pig Longissimus Lumborum and Adductor Muscles. Folia Biol. (Praha) 2015, 63, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Tomisaka, Y.; Ahhmed, A.M.; Tabata, S.; Kawahara, S.; Muguruma, M. Changes in water-holding capacity and textural properties of chicken gizzard stored at 4 °C. Anim. Sci. J. 2010, 81, 362–368. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Stromer, M.H.; Chou, R.-G.R. μ-Calpain is involved in the postmortem proteolysis of gizzard smooth muscle. Food Chem. 2013, 139, 384–388. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Hsu, D.-H.; Stromer, M.H.; Chou, R.-G.R. Postmortem calpain changes in ostrich skeletal muscle. Meat Sci. 2016, 117, 117–121. [Google Scholar] [CrossRef]
- Goll, D.E.; Thompson, V.F.; Li, H.; Wei, W.; Cong, J. The calpain system. Physiol. Rev. 2003, 83, 731–801. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Chou, R.-G.R. Postmortem degradation of desmin and calpain in breast and leg and thigh muscles from Taiwan black-feathered country chickens. J. Sci. Food Agric. 2010, 90, 2664–2668. [Google Scholar] [CrossRef]
- Boehm, M.L.; Kendall, T.L.; Thompson, V.F.; Goll, D.E. Changes in the calpains and calpastatin during postmortem storage of bovine muscle. J. Anim. Sci. 1998, 76, 2415–2434. [Google Scholar] [CrossRef] [PubMed]
- Geesink, G.H.; Taylor, R.G.; Koohmaraie, M. Calpain 3/p94 is not involved in postmortem proteolysis. J. Anim. Sci. 2005, 83, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Bee, G.; Anderson, A.L.; Lonergan, S.M.; Huff-Lonergan, E. Rate and extent of pH decline affect proteolysis of cytoskeletal proteins and water-holding capacity in pork. Meat Sci. 2007, 76, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Sanoudou, D.; Kang, P.B.; Haslett, J.N.; Han, M.; Kunkel, L.M.; Beggs, A.H. Transcriptional profile of postmortem skeletal muscle. Physiol. Genom. 2004, 16, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Poloz, Y.; O’Day, D.H. The Use of Protein Markers for the Estimation of the Postmortem Interval. In Forensic Pathology Reviews; Turk, E.E., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 277–294. ISBN 978-1-61779-249-6. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author and Year | Study Type | Tissue | Method | Model | Sample Size and Study Groups |
|---|---|---|---|---|---|
| Wehner et al., 1999 [29] | research article | pancreas | immunohisto-chemistry | H | n = 128 individuals with varying PMI |
| Wehner et al., 2000 [31] | research article | thyroid gland | immunohisto-chemistry | H | n = 147 individuals with varying PMI |
| Wehner et al., 2001 [30] | research article | thyroid gland | immunohisto-chemistry | H | n = 214 individuals with varying PMI |
| Wehner et al., 2001 [28] | research article | pancreas | immunohisto-chemistry | H | n = 136 individuals with varying PMI |
| Thaik-Oo et al., 2002 [32] | research article | brain, heart, liver, lung, kidney | enzyme-linked immunosorbent assay (ELISA) | H | n = 19 individuals with varying PMI |
| Kang et al., 2003 [46] | research article | lung, skeletal muscle | Western blot | A | n = 16 individuals; several groups/samples at different time points pm |
| Sabucedo and Furton, 2003 [33] | research article | heart | Western blot | A, H | animal: n = 3 hearts of 3 individuals; samples at different time points pm; human: 7 hearts of 7 individuals; several samples at different time points pm, samples of cases with varying PMI |
| Wehner et al., 2006 [27] | research article | brain, pancreas | immunohisto-chemistry | H | n = 500 individuals with varying PMI |
| Poloz and O’Day, 2009 [22] | research article | lung, skeletal muscle | Western blot | A | n = 40 individuals; several groups/samples at different time points pm, different temperature regimes |
| Boaks et al., 2014 [53] | research article | bone | histology/photometry | A | n = 12 bones of unknown (5?) individuals; several groups/samples at different time points pm |
| Bolton et al., 2015 [54] | research article | cartilage | Western blot | A | n = 33 trotters (individuals unknown); several groups/samples at different time points pm |
| Kumar et al., 2015 [35] | research article | heart | Western blot | H | n = 9 hearts of 9 individuals with varying PMI, several samples at different time points pm |
| Kumar et al., 2015 [34] | research article | heart | Western blot | H | n = 5 hearts of 5 individuals with varying PMI; several samples at different time points pm |
| Kumar et al., 2015 [36] | research article | heart | Western blot | H | n = 10 hearts of 10 individuals with varying PMI; several samples at different time points pm |
| Abo El-Noor et al., 2016 [47] | research article | heart, kidney | enzyme activity assay | A | n = 84 individuals; several samples at different time points pm |
| Lee et al., 2016 [48] | research article | kidney, skeletal muscle | immunohisto-chemistry, lateral flow assay, Western blot | A | kidney: n = 48 individuals, skeletal muscle: n = 40 individuals; several groups/samples at different time points pm |
| Li et al., 2016 [25] | review article | biuret method, immunohisto-chemistry, Western blot | A | n.i. | |
| Foditsch et al., 2016 [55] | research article | skeletal muscle | sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), Western blot | A | n = 2 individuals; samples at different time points pm, different temperature regimes |
| Kumar et al., 2016 [37] | research article | heart | Western blot | H | n = 6 hearts of 6 individuals; samples at differnet time points pm of cases with varying PMI, different temperature regimes |
| Pittner et al., 2016 [21] | research article | skeletal muscle | casein zymography, SDS-PAGE, Western blot | A | n = 6 hind limbs of 3 individuals; several groups/samples at different time points pm |
| Pittner et al., 2016 [38] | research article | skeletal muscle | casein zymography, Western blot | H | n = 40 individuals with varying PMI |
| Kwak et al., 2017 [50] | research article | liver | 2-dimensional (2D)-PAGE, mass spectrometry | A | n = 3 individuals; samples at different time points pm |
| Li et al., 2017 [42] | research article | liver | mass spectrometry | A, H | animal: n = 36 individuals, several groups/samples at different time points pm; human: n = 24 + 4 individuals |
| Li et al., 2017 [51] | research article | skeletal muscle | mass spectrometry | A | n = 4 individuals; samples at different time points pm |
| Ortmann et al., 2017 [41] | research article | pancreas, thyroid gland | immunohisto-chemistry | H | n = 105 individuals with varying PMI |
| Pérez-Martínez et al., 2017 [40] | research article | bone | mass spectrometry | H | n = 80 bones of 80 individuals with varying PMI |
| Pittner et al., 2017 [39] | case study | skeletal muscle | Western blot | H | n = 2 individuals with unknown PMI, forensic case |
| Jellinghaus et al., 2018 [56] | research article | bone | histology/photometry, histology | H | n = 16 bones (individuals unclear); samples at differnent time points pm |
| Prieto-Bonete et al., 2018 [23] | research article | bone | mass spectrometry | H | n = 40 bones of 40 individuals with varying PMI |
| Procopio et al., 2018 [57] | research article | bone | mass spectrometry | A | n = 8 bones of 4 individuals; samples at different time points pm |
| Zissler et al., 2018 [49] | research article | skeletal muscle | Western blot | A | n = 20 individuals; several groups/samples at different time points pm |
| Alibegovic et al., 2019 [44] | research article | cartilage | histology | H | n = 3 |
| Cho and Eom, 2019 [52] | research article | lung | immunohisto-chemistry, Western blot | A | n = 45 individuals; several groups/samples at different time points |
| Choi et al., 2019 [45] | research article | skeletal muscle | mass spectrometry, Western blot | A, H | n = 20 (rat), n = 10 (mouse), n = 3 human |
| da Fonseca et al., 2019 [58] | research article | brain, liver, skeletal muscle, kidney | enzyme activity assay | A | n = 24–28 (unclear); several groups/samples |
| Jellinghaus et al., 2019 [43] | research article | bone | histology/photometry, histology | H | n = 35 forensic; n = 11 museum |
| Author and Year | Method | Target (Species, Specific Tissue Location) | PMI | Samples and Sampling Frequency | Storage Conditions | Investigated Proteins | Type of Study Outcome | Main Study Outcome |
|---|---|---|---|---|---|---|---|---|
| Wehner et al., 1999 [29] | immunohistochemistry | human pancreatic α-cells | 1–45 ± 1 dpm | 128 samples; 1–13 samples per time point (41) | varying | Insulin | positive and negative immunostaining | loss of stainability over PMI |
| Wehner et al., 2000 [31] | immunohistochemistry | human thyroid gland | 1–21 ± 1 dpm | 147 samples; 1–20 samples per time point (21) | varying | Thyroglobulin | positive and negative immunostaining | loss of stainability over PMI |
| Wehner et al., 2001 [30] | immunohistochemistry | human thyroid gland | 1–21 ± 1 dpm | 214 samples; 1–20 samples per time point (21) | varying | Calcitonin | positive and negative immunostaining | loss of stainability over PMI |
| Wehner et al., 2001 [28] | immunohistochemistry | human pancreatic β- cells | 1–21 ± 1 dpm | 1–29 samples per time point (21); 1 sample per individual (136) | varying | Glucagon | positive and negative immunostaining | loss of stainability over PMI |
| Thaik-Oo et al., 2002 [32] | ELISA | human frontal lobe of the cerebrum, middle lobe of the right lung, apex of the heart, lower part of the liver, upper part of the kidney | 2.75–120 hpm | 19 samples per tissue; PMI between 2.75 and 120 hpm | varying | vascular endothelial growth factor VEGF | concentration of protein ng/mL | increase and subsequent decrease over PMI |
| Kang et al., 2003 [46] | Western blot | rat lung and skeletal muscle | 0–96 hpm | 4 samples per time point (4) per tissue | laboratory-controlled, 21 °C | calcineurin A (CnA), myristoylated alanine-rich C-kinase substrate (MARCKS), Ca2+/calmodulin-dependent protein kinase II (CaMKII), inducible nitric oxide synthase (iNOS) | band intensity, % of intact protein | significant decrease of band intensity over PMI; degradation products |
| Sabucedo and Furton, 2003 [33] | Western blot | bovine heart, human heart | bovine:0–6 dpm, “zero-hour” donor: 0–168 hpm; “PMI” donors: 8–12 hpm + 24 h incubation | animal: several samples at several time points, human: several samples of one individual at several postmortem time points + 12 samples of 6 individuals at different PMI | varying before autopsy; laboratory-controlled after autopsy, 20 ± 2 °C, | Troponin I (TnI) | band intensity, % of intact protein | pseudo-first order relationship between % of intact protein and log of time, decrease and fragmentation of protein over PMI |
| Wehner et al., 2006 [27] | immunohistochemistry | human frontal cortex and pancreas | 1–23 ± 1 dpm | number of samples per time point is unknown; 1 sample per tissue per individual | varying | glial fibrillary acidic protein, somatostatin | positive and negative immunostaining | loss of stainability over PMI, temperature dependence (winter vs. summer) |
| Poloz and O’Day, 2009 [22] | Western blot | mouse lung and skeletal muscle | 0–96 hpm | 4 samples per time point (4) and per temperature (3) | laboratory-controlled; 5 °C, 10 °C, 21 °C | CnA, MARCKS, CaMKII, protein phosphatase 2A (PP2A) | band intensity, % of intact protein | significant decrease of band intensity over PMI; degradation products; significant effects of temperature |
| Boaks et al., 2014 [53] | histology/photometry | porcine long bones (extremities) | 0–12 month pm | 2 samples per time point (6) | outdoor; cadavers stored in kennels at surface of field, environmental conditions (e.g., temperature) unknown | Collagen | ratio of collagenous to non collagenous protein (Co/NCo) concentration | decrease in (Co/NCo) ratio |
| Bolton et al., 2015 [54] | Western blot | cartilage of porcine metacarpotarsal and metacarpophalangeal joints | 0–6 weeks pm | 2 samples per time point (7) | outdoor; buried in different soil environments, soil: 9–18 °C, air: 8–28 °C | Aggrecan | qualitative assessment of band presence/absence over PMI | loss of protein over PMI |
| Kumar et al., 2015 [35] | Western blot | human heart | unclear: probably up to 88.4 hpm | several samples at several time points | varying before autopsy; laboratory controlled after autopsy, 20 ± 2 °C | cardiac troponin T (cTnT) | band intensity, % of intact protein; migration distance of bands | significant decrease of band intensity over PMI, degradation products |
| Kumar et al., 2015 [34] | Western blot | human heart | approx. 5–230 hpm | several samples at several time points | varying before autopsy; laboratory controlled after autopsy, 20 ± 2 °C | cTnT | band intensity, % of intact protein; migration distance of bands | significant decrease of band intensity over PMI, degradation products |
| Kumar et al., 2015 [36] | Western blot | human heart | unclear: probably up to 88.4 hpm | several samples at several time points | varying before autopsy; room temperature after autopsy | cTnT | band intensity, % of intact protein; migration distance of bands | significant decrease of band intensity over PMI, degradation products |
| Abo El-Noor et al., 2016 [47] | enzyme activity assay | rat heart and kidney | 0–7 hpm | 12 samples per time point (7) | laboratory-controlled; 22 °C, 15% relative humidity | catalase, gluthatione-S-transferase, glutathione reductase | change of enzyme activity | significant decrease of enzyme activity over PMI |
| Lee et al., 2016 [48] | immunohistochemistry, lateral flow assay (LFA), Western blot | rat kidney and psoas muscle | 0–96 hpm | 5–6 samples per time points (8) | laboratory controlled; 23 ± 1 °C | glyceraldehyde-3-phosphat-dehydrogenase (GAPDH), caspase-3, peroxisome proliferator-activated receptor-γ (PPAR-γ), glycogen synthase, glycogen synthase kinase-3β (GSK-3β), p53, 5’ AMP-activated kinase α (AMPKα), beta catenin | WB: band intensity, % of intact protein, calculation of half maximum intensity of intact protein PMI50; IHC: qualitative assessment of staining intensity; pilot experiments to develop LFA-based chip | WB: significant decrease of band intensity over PMI; IHC: loss of stainability over PMI; LFA-based chip: development of rGAPDH immunosensor |
| Li et al., 2016 [25] | Biuret method, immunohistochemistry, Western blot | rat cardiac muscle, brain, liver, lung, kidney, skeletal muscle, and spleen; human cardiac and skeletal muscle; rabbit skeletal muscle | 48 hpm, 54 hpm, 72 hpm, 5 dpm, 7 dpm | not indicated | not indicated | actin, tubulin, myoglobin, troponin I | not indicated, varying | not indicated, varying |
| Foditsch et al., 2016 [55] | SDS PAGE, Western blot | porcine biceps femoris muscle | 4 °C: 0–21 dpm, 22 °C: 0–5 dpm | 1 sample per temperature and per time point | 4 ± 1 °C, 22 ± 2 °C | α-actinin, calsequestrin 1, desmin, nebulin, titin, sarcoplasmic/endoplasmic reticulum Ca2+ ATPase-1 (SERCA-1), SERCA-2, tropomyosin, cTNT, laminin, µ-calpain | qualitative assessment of band presence/absence over PMI | loss of proteins over PMI, degradation products |
| Kumar et al., 2016 [37] | Western blot | human heart | unclear: probably up to 189 hpm | 6 individuals; several samples at several time points and temperatures | 12 °C, 20 ± 2°C, 25 °C, 37 °C | cTnT | not defined, probably % of intact protein | significant decrease/loss of intact protein over PMI, degradation products, effect of temperature |
| Pittner et al., 2016 [21] | casein zymography, SDS PAGE, Western blot | porcine biceps femoris muscle | 0–10 dpm | 6 samples per time point (17) | 21 ± 1 °C, 35% relative humidity | titin, tropomyosin, nebulin, desmin, cTNT, SERCA-1, capain-1, calpain-2 | mean time points (+ 95% confidence interval) of band change (presence to absence and vice versa) | different mean time points of band change of different proteins and degradation products |
| Pittner et al., 2016 [38] | casein zymography, Western blot | human vastus lateralis muscle | 4–93 hpm | 40 samples at differnet time points | varying, accumulated degree days calculated | desmin, tropomyosin, calpain-1, calpain-2 | presence and absence probability of bands at different accumulated degree days; correlation of band presence and absence with ADD | different probability of presence for different proteins and degradation products over ADD; significant correlations between proteins/degradation products and ADD |
| Kwak et al., 2017 [50] | 2D-PAGE | rat liver and heart | 0–48 hpm | 3 samples per time point (3) | laboratory-controlled; 23 ± 3 °C, relative humidity 60 ± 5% | listed in supplementary material | spot intensity | increase and decrease of spot intensity over PMI |
| Li et al., 2017 [42] | mass spectrometry | rat liver, human right posterior liver lobe | rats: 0–144 hpm, human: estimated PMI + 0–144 hpm (corresponds to 10–168 hpm) | several samples per time point (4) | laboratory-controlled; 23 ± 1 °C, 30–45% relative humidity | listed in supplementary material | signal strength of peptide/protein peaks | decrease of signal strength of various protein/peptide peaks over PMI |
| Li et al., 2017 [51] | mass spectrometry | rat quadriceps femoris muscle | 0–144 hpm | 1 sample per time point (4) | laboratory controlled; 23 ± 1 °C, 30–45% relative humidity | listed in supplementary material | signal strength of peptide/protein peaks | decrease of signal strength of various protein/peptide peaks over PMI |
| Ortmann et al., 2017 [41] | immunohistochemistry | human pancreas, human thyroid gland | up to 22 dpm | 1–18 samples per time point (12) | varying | calcitonin, thyroglobulin, insulin, glucagon | positive and negative immunostaining | loss of stainability over PMI |
| Pérez-Martínez et al., 2017 [40] | mass spectrometry | human femur, tibia and humerus | 5–47 years pm | 80 samples at different time points | Outdoor; cemetery, winters 5–19 °C, summers 22–40 °C | collagen type I | abundance of proteins (number of peptides) | significant decrease of collagen concentration over PMI |
| Pittner et al., 2017 [39] | Western blot | human vastus lateralis muscle | unknown | 1 sample per individual | partly unknown | desmin, troponin T, calpain 1, calpain 2, tropomyosin | presence and absence of bands in two individuals to trace progression of events in murder-suicide case | difference (presence and absence of proteins) in protein profile of two individuals |
| Jellinghaus et al., 2018 [56] | histology/photometry, histology | porcine right and left femur | 0–3 month pm | 8 samples per time point (4) | buried in boxes; 13–34 °C (monitored); 2 groups with different (hay and distilled) water infusion | collagen | ratio of collagenous to non collagenous protein (Co/NCo) concentration | decrease in (Co/NCo) ratio |
| Prieto-Bonete et al., 2018 [23] | mass spectrometry | human proximal femur | 5–20 years pm | 40 samples at differnet time points/PMI | outdoor; cemetery, weather data available online | listed in supplementary material | presence and absence of proteins at different PMI | loss of proteins over PMI |
| Procopio et al., 2018 [57] | mass spectrometry | porcine tibia | 0–1 year pm | 1 respectively 4 samples per time point (5) | outdoor, buried in soil, temperature data not available | listed in supplementary material | abundance of different proteins at different PMI | decrease in protein amount with PMI |
| Zissler et al., 2018 [49] | Western blot | rat quadriceps femoris muscle | 0–4 dpm | 4 samples per time point (5) | laboratory-controlled; 20 °C | desmin, vinculin, tropomyosin | change of band pattern (presence and absence of bands) over PMI; band intensity in % of intact protein | decrease of band intenstiy over PMI; significant loss of protein bands over PMI; degradation products |
| Alibegovic et al., 2019 [44] | histology | cartilage of human trochlea, medial and lateral condyle | estimated PMI (30–48 hpm) + 1–36 dpm | 3 samples per time point (3) and per temperature | varying before autopsy; laboratory-controlled after autopsy, storage of samples in tubes, 11 ± 2 °C, 35 ± 2 °C | collagen, proteoglycan | intensity of histological staining; Bern grading scale | significant decrease in staining intensity over PMI; no significant difference between temperatures |
| Cho and Eom, 2019 [52] | immunohistochemistry, Western blot | rat lung | 1–7 dpm | 5 samples per time point (7) | laboratory-controlled; immersion in sea water, 15 ± 5 °C | RAGE | Western blot: band intensity in reference to housekeeping protein (GAPDH); immunohistochemistry: qualitative assessment of staining intensity | decrease of band intensity over PMI; decrease of IHC staining intensity |
| Choi et al., 2019 [45] | mass spectrometry, Western blot | mouse, rat, human vastus lateralis muscle | 0–96 hpm | mouse: 2 samples per time point (5); rat: 2 samples per time point (5) for MS, 5 samples per time point (5) for WB, human: 3 samples at differnt time points | laboratory-controlled in rat: 20 °C, mouse: 25 °C; human: varying | eukaryotic translation elongation factor 1A2 (eEF1A2), GAPDH, tropomyosin, desmin, vinculin | MS: decrease of abundance over PMI, WB: change of band pattern (presence and absence of bands) over PMI | MS: decrease of proteins over PMI; WB: significant loss of bands over PMI; degradation products |
| da Fonseca et al., 2019 [58] | enzyme activity assay | mouse liver, whole brain, gastrocnemius muscle, kidney | 0–48 hpm | 6–7 samples per time point (4) | laboratory-controlled; 22 ± 2 °C | Na+/K+ ATPase, Acethylcholinesterase, gluthatione-S-transferase | change of enzyme activity | significant increase and decrease of enzyme activity over PMI/at time points pm |
| Jellinghaus et al., 2019 [43] | histology/photometry, histology | human femur | up to 171 years pm | 46 samples at different time points | outdoor; cemetery and archeological samples (museum) | collagen | ratio of collagenous to non collagenous protein (Co/NCo) concentration | decrease in (Co/NCo) ratio |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zissler, A.; Stoiber, W.; Steinbacher, P.; Geissenberger, J.; Monticelli, F.C.; Pittner, S. Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review. Diagnostics 2020, 10, 1014. https://doi.org/10.3390/diagnostics10121014
Zissler A, Stoiber W, Steinbacher P, Geissenberger J, Monticelli FC, Pittner S. Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review. Diagnostics. 2020; 10(12):1014. https://doi.org/10.3390/diagnostics10121014
Chicago/Turabian StyleZissler, Angela, Walter Stoiber, Peter Steinbacher, Janine Geissenberger, Fabio C. Monticelli, and Stefan Pittner. 2020. "Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review" Diagnostics 10, no. 12: 1014. https://doi.org/10.3390/diagnostics10121014
APA StyleZissler, A., Stoiber, W., Steinbacher, P., Geissenberger, J., Monticelli, F. C., & Pittner, S. (2020). Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review. Diagnostics, 10(12), 1014. https://doi.org/10.3390/diagnostics10121014