Nitrates as a Potential N Supply for Microbial Ecosystems in a Hyperarid Mars Analog System


Abstract
1. Introduction
2. Materials and Methods
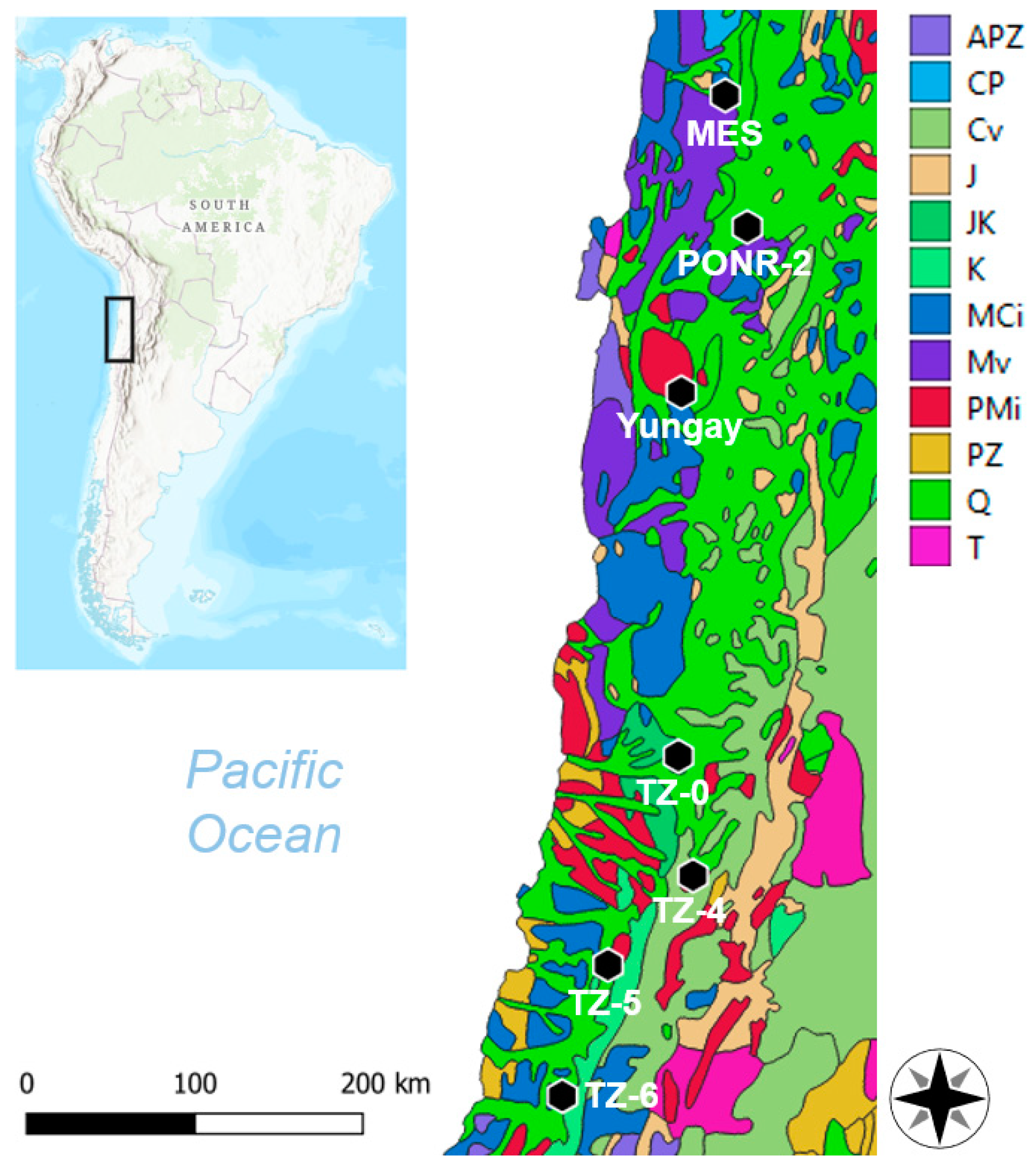
2.1. Study Sites
2.2. Soil Characterization
2.3. Nitrate and Ammonium Measurements
2.4. Total Organic Carbon (TOC) and Total Nitrogen (TN)
2.5. Nitrate Amendments and Colony-Forming Units (CFUs)
2.6. Hierarchical Clustering, Correlation and Linear Regression Analyses
3. Results
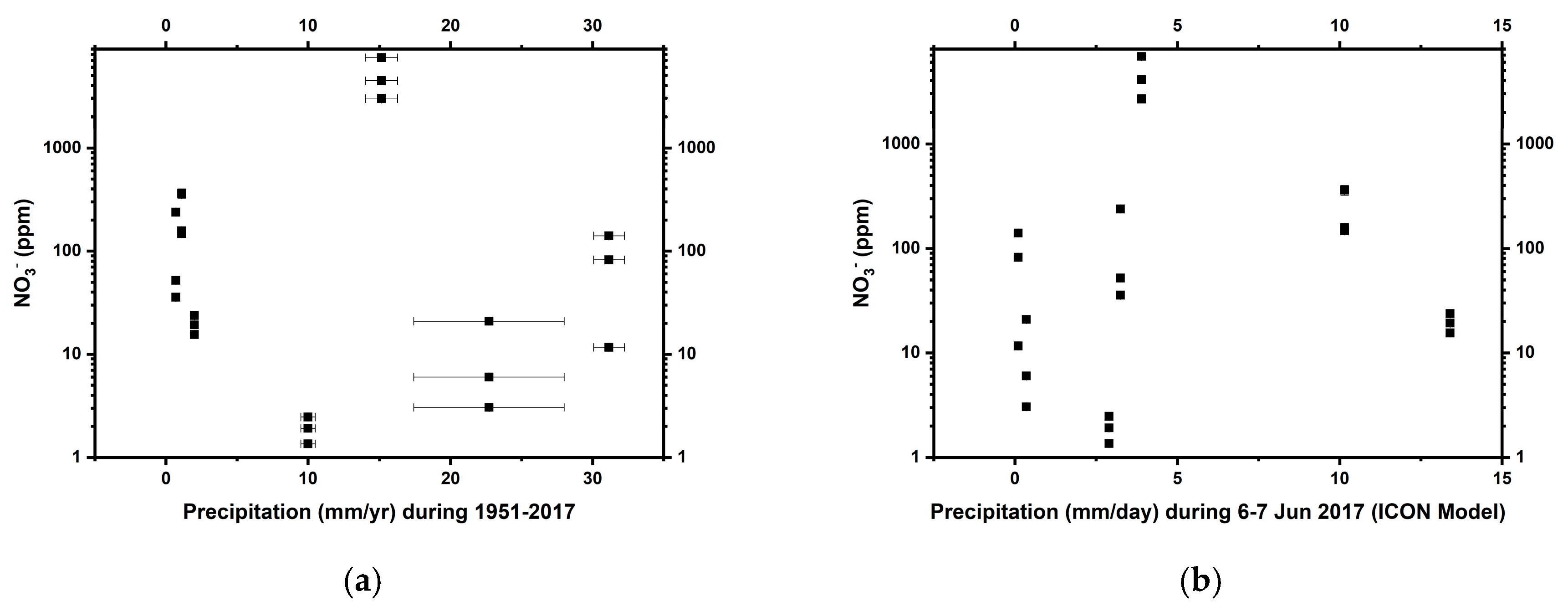
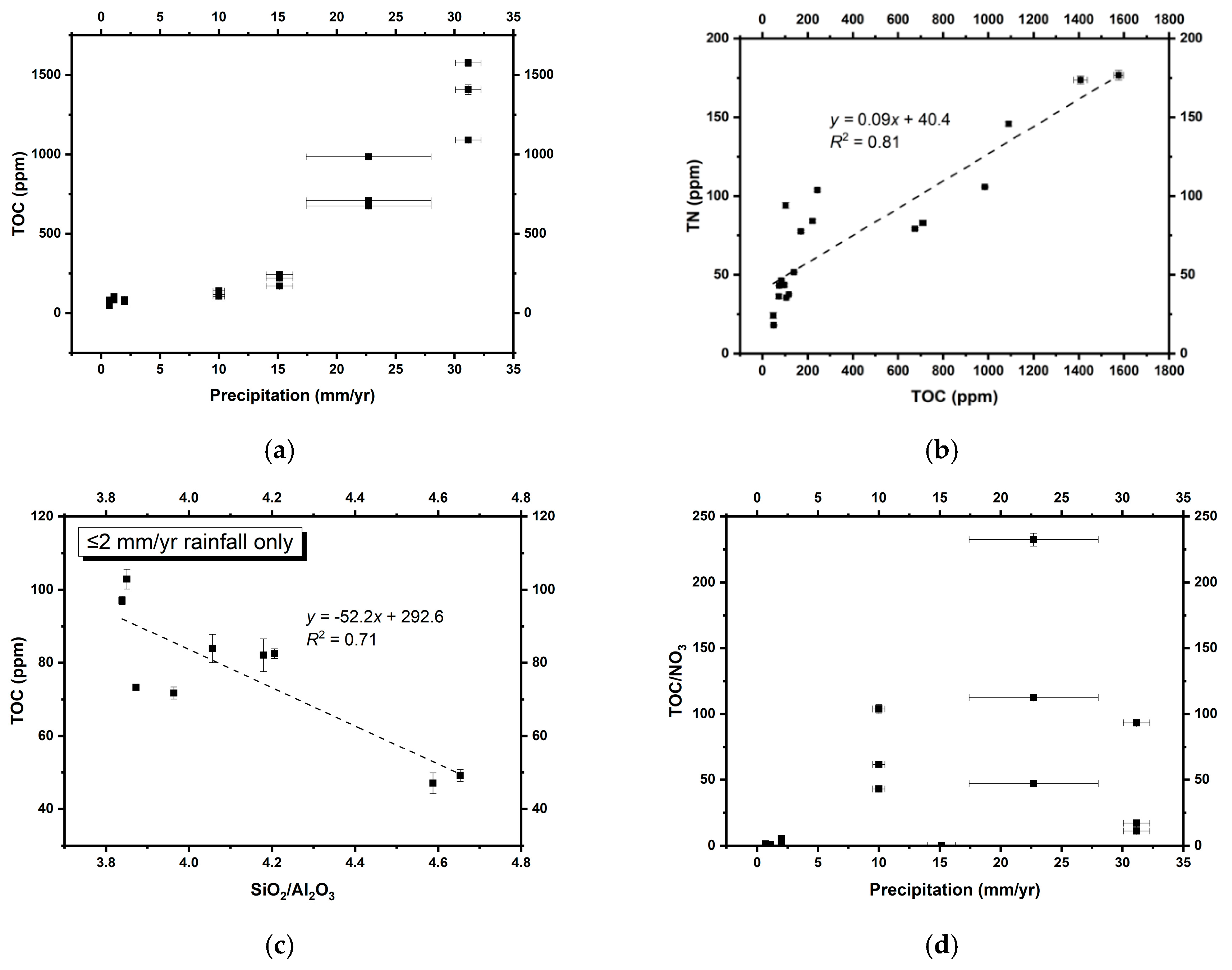
3.1. Physical and Chemical Properties of the Soils
3.2. Cell Counts on Agar Plates
4. Discussion
4.1. Site Categorization
4.2. Nitrate Distribution and Effects on Microbial N Assimilation
4.3. Growth of Cell Cultures with Nitrate Amendments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franz, H.B.; Trainer, M.G.; Malespin, C.A.; Mahaffy, P.R.; Atreya, S.K.; Becker, R.H.; Benna, M.; Conrad, P.G.; Eigenbrode, J.L.; Freissinet, C.; et al. Initial SAM calibration gas experiments on Mars: Quadrupole mass spectrometer results and implications. Planet. Space Sci. 2017, 138, 44–54. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Mancinelli, R.L.; Kern, R.G.; Rothschild, L.J.; McKay, C.P. Survival of endospores of Bacillus subtilis on spacecraft surfaces under simulated martian environments: Implications for the forward contamination of Mars. Icarus 2003, 165, 253–276. [Google Scholar] [CrossRef]
- Stern, J.C.; Sutter, B.; Freissinet, C.; Navarro-Gonzalez, R.; McKay, C.P.; Archer, P.D.; Buch, A.; Brunner, A.E.; Coll, P.; Eigenbrode, J.L.; et al. Evidence for indigenous nitrogen in sedimentary and aeolian deposits from the Curiosity rover investigations at Gale crater, Mars. Proc. Natl. Acad. Sci. USA 2015, 112, 4245–4250. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.H.; Head, J.W. Geologic history of Mars. Earth Planet. Sci. Lett. 2010, 294, 185–203. [Google Scholar] [CrossRef]
- Fuller, E.R.; Head, J.W. Amazonis Planitia: The role of geologically recent volcanism and sedimentation in the formation of the smoothest plains on Mars. J. Geophys. Res. Planet. 2002, 107. [Google Scholar] [CrossRef]
- Clifford, S.M.; Parker, T.J. The evolution of the Martian hydrosphere: Implications for the fate of a primordial ocean and the current state of the northern plains. Icarus 2001, 154, 40–79. [Google Scholar] [CrossRef]
- Di Achille, G.; Hynek, B.M. Ancient ocean on Mars supported by global distribution of deltas and valleys. Nat. Geosci. 2010, 3, 459–463. [Google Scholar] [CrossRef]
- Malin, M.C.; Edgett, K.S. Evidence for persistent flow and aqueous sedimentation on early Mars. Science 2003, 302, 1931–1934. [Google Scholar] [CrossRef]
- Irwin, R.P.; Maxwell, T.A.; Howard, A.D.; Craddock, R.A.; Leverington, D.W. A large paleolake basin at the head of Ma’adim Vallis, Mars. Science 2002, 296, 2209–2212. [Google Scholar] [CrossRef]
- Craddock, R.A.; Howard, A.D. The case for rainfall on a warm, wet early Mars. J. Geophys. Res. Planet. 2002, 107. [Google Scholar] [CrossRef]
- Craddock, R.A.; Lorenz, R.D. The changing nature of rainfall during the early history of Mars. Icarus 2017, 293, 172–179. [Google Scholar] [CrossRef]
- Leshin, L.A.; Mahaffy, P.R.; Webster, C.R.; Cabane, M.; Coll, P.; Conrad, P.G.; Archer, P.D.; Atreya, S.K.; Brunner, A.E.; Buch, A.; et al. Volatile, Isotope, and Organic Analysis of Martian Fines with the Mars Curiosity Rover. Science 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Meslin, P.Y.; Gasnault, O.; Forni, O.; Schroder, S.; Cousin, A.; Berger, G.; Clegg, S.M.; Lasue, J.; Maurice, S.; Sautter, V.; et al. Soil Diversity and Hydration as Observed by ChemCam at Gale Crater, Mars. Science 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Tice, A.R. Analysis of Water in the Martian Regolith. J. Mol. Evol. 1979, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Martin-Torres, F.J.; Zorzano, M.P.; Valentin-Serrano, P.; Harri, A.M.; Genzer, M.; Kemppinen, O.; Rivera-Valentin, E.G.; Jun, I.; Wray, J.; Madsen, M.B.; et al. Transient liquid water and water activity at Gale crater on Mars. Nat. Geosci. 2015, 8, 357–361. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Caro-Lara, L.; Vicuna, R. Discovery and microbial content of the driest site of the hyperarid Atacama Desert, Chile. Environ. Microbiol. Rep. 2015, 7, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Silva, B.; Rainey, F.A.; Warren-Rhodes, K.A.; McKay, C.P.; Navarro-González, R. Atacama Desert soil microbiology. In Microbiology of Extreme Soils; Springer: Berlin/Heidelberg, Germany, 2008; pp. 117–132. [Google Scholar]
- Navarro-Gonzalez, R.; Rainey, F.A.; Molina, P.; Bagaley, D.R.; Hollen, B.J.; de la Rosa, J.; Small, A.M.; Quinn, R.C.; Grunthaner, F.J.; Caceres, L.; et al. Mars-like soils in the Atacama Desert, Chile, and the dry limit of microbial life. Science 2003, 302, 1018–1021. [Google Scholar] [CrossRef]
- Veblen, T.T.; Young, K.R.; Orme, A.R. The Physical Geography of South. America; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- McKay, C.P.; Friedmann, E.I.; Gomez-Silva, B.; Caceres-Villanueva, L.; Andersen, D.T.; Landheim, R. Temperature and moisture conditions for life in the extreme arid region of the Atacama Desert: Four years of observations including the El Nino of 1997–1998. Astrobiology 2003, 3, 393–406. [Google Scholar] [CrossRef]
- Hartley, A.J.; Chong, G.; Houston, J.; Mather, A.E. 150 million years of climatic stability: Evidence from the Atacama Desert, northern Chile. J. Geol. Soc. Lond. 2005, 162, 421–424. [Google Scholar] [CrossRef]
- Houston, J. Variability of precipitation in the Atacama desert: Its causes and hydrological impact. Int. J. Clim. 2006, 26, 2181–2198. [Google Scholar] [CrossRef]
- Sun, T.; Bao, H.M.; Reich, M.; Hemming, S.R. More than ten million years of hyper-aridity recorded in the Atacama Gravels. Geochim. Cosmochim. Acta 2018, 227, 123–132. [Google Scholar] [CrossRef]
- Cordero, R.R.; Damiani, A.; Jorquera, J.; Sepulveda, E.; Caballero, M.; Fernandez, S.; Feron, S.; Llanillo, P.J.; Carrasco, J.; Laroze, D.; et al. Ultraviolet radiation in the Atacama Desert. Antonie Van Leeuwenhoek 2018, 111, 1301–1313. [Google Scholar] [CrossRef]
- Lester, Y.; Sharpless, C.M.; Mamane, H.; Linden, K.G. Production of Photo-oxidants by Dissolved Organic Matter During UV Water Treatment. Environ. Sci. Technol. 2013, 47, 11726–11733. [Google Scholar] [CrossRef]
- Quinn, R.C.; Ehrenfreund, P.; Grunthaner, F.J.; Taylor, C.L.; Zent, A.P. Decomposition of aqueous organic compounds in the Atacama Desert and in Martian soils. J. Geophys. Res. Biogeosci. 2007, 112. [Google Scholar] [CrossRef]
- Quinn, R.C.; Zent, A.P.; Grunthaner, F.J.; Ehrenfreund, P.; Taylor, C.L.; Garry, J.R.C. Detection and characterization of oxidizing acids in the Atacama Desert using the Mars Oxidation Instrument. Planet. Space Sci. 2005, 53, 1376–1388. [Google Scholar] [CrossRef]
- Michalski, G.; Bohlke, J.K.; Thiemens, M. Long term atmospheric deposition as the source of nitrate and other salts in the Atacama Desert, Chile: New evidence from mass-independent oxygen isotopic compositions. Geochim. Cosmochim. Acta 2004, 68, 4023–4038. [Google Scholar] [CrossRef]
- Ericksen, G.E. Geology and Origin of the Chilean Nitrate Deposits; United States Geological Survey Professional Paper; United States Government Printing Office: Washington, DC, USA, 1981. [Google Scholar]
- Ericksen, G.E. The Chilean Nitrate Deposits. Am. Sci. 1983, 71, 366–374. [Google Scholar]
- Walvoord, M.A.; Phillips, F.M.; Stonestrom, D.A.; Evans, R.D.; Hartsough, P.C.; Newman, B.D.; Striegl, R.G. A reservoir of nitrate beneath desert soils. Science 2003, 302, 1021–1024. [Google Scholar] [CrossRef]
- Jackson, W.A.; Bohlke, J.K.; Andraski, B.J.; Fahlquist, L.; Bexfield, L.; Eckardt, F.D.; Gates, J.B.; Davila, A.F.; McKay, C.P.; Rao, B.; et al. Global patterns and environmental controls of perchlorate and nitrate co-occurrence in arid and semi-arid environments. Geochim. Cosmochim. Acta 2015, 164, 502–522. [Google Scholar] [CrossRef]
- Kounaves, S.P.; Carrier, B.L.; O’Neil, G.D.; Stroble, S.T.; Claire, M.W. Evidence of martian perchlorate, chlorate, and nitrate in Mars meteorite EETA79001: Implications for oxidants and organics. Icarus 2014, 229, 206–213. [Google Scholar] [CrossRef]
- Jaramillo, E.A.; Royle, S.H.; Claire, M.W.; Kounaves, S.P.; Sephton, M.A. Indigenous Organic-Oxidized Fluid Interactions in the Tissint Mars Meteorite. Geophys. Res. Lett. 2019, 46, 3090–3098. [Google Scholar] [CrossRef]
- Ewing, S.A.; Michalski, G.; Thiemens, M.; Quinn, R.C.; Macalady, J.L.; Kohl, S.; Wankel, S.D.; Kendall, C.; Mckay, C.P.; Amundson, R. Rainfall limit of the N cycle on Earth. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Fairen, A.G.; Gonzalez-Silva, C.; Ascaso, C.; Carrizo, D.; Fernandez-Martinez, M.A.; Fernandez-Sampedro, M.; Garcia-Descalzo, L.; Garcia-Villadangos, M.; Martin-Redondo, M.P.; et al. Unprecedented rains decimate surface microbial communities in the hyperarid core of the Atacama Desert. Sci. Rep. 2018, 8, 16706. [Google Scholar] [CrossRef]
- Bohlke, J.K.; Ericksen, G.E.; Revesz, K. Stable isotope evidence for an atmospheric origin of desert nitrate deposits in northern Chile and southern California, USA. Chem. Geol. 1997, 136, 135–152. [Google Scholar] [CrossRef]
- Catling, D.C.; Claire, M.W.; Zahnle, K.J.; Quinn, R.C.; Clark, B.C.; Hecht, M.H.; Kounaves, S. Atmospheric origins of perchlorate on Mars and in the Atacama. J. Geophys. Res. Planet. 2010, 115. [Google Scholar] [CrossRef]
- Mancinelli, R.L.; Mckay, C.P. The Evolution of Nitrogen Cycling. Orig. Life Evol. Biosph. 1988, 18, 311–325. [Google Scholar] [CrossRef]
- Manning, C.V.; Mckay, C.P.; Zahnle, K.J. The nitrogen cycle on Mars: Impact decomposition of near-surface nitrates as a source for a nitrogen steady state. Icarus 2008, 197, 60–64. [Google Scholar] [CrossRef]
- Manning, C.V.; Zahnle, K.J.; Mckay, C.P. Impact processing of nitrogen on early Mars. Icarus 2009, 199, 273–285. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, R.; Navarro, K.F.; Coll, P.; McKay, C.P.; Stern, J.C.; Sutter, B.; Archer, P.D.; Buch, A.; Cabane, M.; Conrad, P.G.; et al. Abiotic Input of Fixed Nitrogen by Bolide Impacts to Gale Crater During the Hesperian: Insights From the Mars Science Laboratory. J. Geophys Res. Planet. 2019, 124, 94–113. [Google Scholar] [CrossRef]
- Lin, J.T.; Stewart, V. Nitrate assimilation by bacteria. Adv. Microb. Physiol. 1998, 39, 1–30. [Google Scholar]
- Hutchins, S.R. Biodegradation of Monoaromatic Hydrocarbons by Aquifer Microorganisms Using Oxygen, Nitrate, or Nitrous-Oxide as the Terminal Electron-Acceptor. Appl. Environ. Microb. 1991, 57, 2403–2407. [Google Scholar]
- Shapleigh, J.P. Dissimilatory and assimilatory nitrate reduction in the purple photosynthetic bacteria. In The Purple Phototrophic Bacteria; Springer: Berlin/Heidelberg, Germany, 2009; pp. 623–642. [Google Scholar]
- van de Graaf, A.A.; Mulder, A.; de Bruijn, P.; Jetten, M.S.; Robertson, L.A.; Kuenen, J.G. Anaerobic oxidation of ammonium is a biologically mediated process. Appl. Environ. Microbiol. 1995, 61, 1246–1251. [Google Scholar]
- Belay, N.; Jung, K.Y.; Rajagopal, B.S.; Kremer, J.D.; Daniels, L. Nitrate as a Sole Nitrogen-Source for Methanococcus-Thermolithotrophicus and Its Effect on Growth of Several Methanogenic Bacteria. Curr. Microbiol. 1990, 21, 193–198. [Google Scholar] [CrossRef]
- Blasco, R.; Martinez-Luque, M.; Madrid, M.P.; Castillo, F.; Moreno-Vivian, C. Rhodococcus sp RB1 grows in the presence of high nitrate and nitrite concentrations and assimilates nitrate in moderately saline environments. Arch. Microbiol. 2001, 175, 435–440. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef]
- Devol, A.H. Nitrogen cycle: Solution to a marine mystery. Nature 2003, 422, 575–576. [Google Scholar] [CrossRef]
- Vidiella, P.E.; Armesto, J.J.; Gutierrez, J.R. Vegetation changes and sequential flowering after rain in the southern Atacama Desert. J. Arid Environ. 1999, 43, 449–458. [Google Scholar] [CrossRef]
- Schenk, C.J.; Viger, R.J.; Anderson, C.P. Maps Showing Geology, Oil and Gas Fields, and Geological Provinces of South. America; US. Geological Survey: Reston, VA, USA, 1999. [Google Scholar]
- Tapia, J.; Gonzalez, R.; Townley, B.; Oliveros, V.; Alvarez, F.; Aguilar, G.; Menzies, A.; Calderon, M. Geology and geochemistry of the Atacama Desert. Antonie Leeuwenhoek 2018, 111, 1273–1291. [Google Scholar] [CrossRef]
- Shirey, T.B. Investigating Microbial Communities and the Environmental Factors Influencing Them in Manmade and Naturally Occurring Systems; University of Alabama: Tuscaloosa, Alabama, 2013. [Google Scholar]
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; de Vera, J.P.; Schmitt-Kopplin, P.; Grossart, H.P.; Parro, V.; Kaupenjohann, M.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Robinson, C.K.; Barnum, T.; Fricke, W.F.; Davila, A.F.; Jedynak, B.; McKay, C.P.; DiRuggiero, J. Colonization patterns of soil microbial communities in the Atacama Desert. Microbiome 2013, 1, 28. [Google Scholar] [CrossRef]
- Wierzchos, J.; Camara, B.; de Los Rios, A.; Davila, A.F.; Almazo, I.M.S.; Artieda, O.; Wierzchos, K.; Gomez-Silva, B.; Mckay, C.; Ascaso, C. Microbial colonization of Ca-sulfate crusts in the hyperarid core of the Atacama Desert: Implications for the search for life on Mars. Geobiology 2011, 9, 44–60. [Google Scholar] [CrossRef]
- Lester, E.D.; Satomi, M.; Ponce, A. Microflora of extreme arid Atacama Desert soils. Soil Biol. Biochem. 2007, 39, 704–708. [Google Scholar] [CrossRef]
- Parro, V.; de Diego-Castilla, G.; Moreno-Paz, M.; Blanco, Y.; Cruz-Gil, P.; Rodriguez-Manfredi, J.A.; Fernandez-Remolar, D.; Gomez, F.; Gomez, M.J.; Rivas, L.A.; et al. A Microbial Oasis in the Hypersaline Atacama Subsurface Discovered by a Life Detector Chip: Implications for the Search for Life on Mars. Astrobiology 2011, 11, 969–996. [Google Scholar] [CrossRef]
- Zangl, G.; Reinert, D.; Ripodas, P.; Baldauf, M. The ICON (ICOsahedral Non-hydrostatic) modelling framework of DWD and MPI-M: Description of the non-hydrostatic dynamical core. Q. J. R. Meteorl. Soc. 2015, 141, 563–579. [Google Scholar] [CrossRef]
- Jochum, K.P.; Nohl, L.; Herwig, K.; Lammel, E.; Stoll, B.; Hofmann, A.W. GeoReM: A new geochemical database for reference materials and isotopic standards. Geostand. Geoanal. Res. 2005, 29, 333–338. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Bagaley, D.R. Uncovering Bacterial Diversity on and below the Surface of A Hyper-Arid Environment, the Atacama Desert, Chile. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Origin(Pro); OriginLab Corporation: Northampton, MA, USA, 2018.
- IBM SPSS Statistics for Windows, 25.0; IBM Corp.: Armonk, NY, USA, 2017.
- Oberlin, E.A.; Claire, M.W.; Kounaves, S.P. Evaluation of the Tindouf Basin Region in Southern Morocco as an Analog Site for Soil Geochemistry on Noachian Mars. Astrobiology 2018, 18, 1318–1328. [Google Scholar] [CrossRef]
- Davis, W.L.; de Pater, I.; McKay, C.P. Rain infiltration and crust formation in the extreme arid zone of the Atacama Desert, Chile. Planet. Space Sci. 2010, 58, 616–622. [Google Scholar] [CrossRef]
- Marion, G.M.; Verburg, P.S.J.; McDonald, E.V.; Arnone, J.A. Modeling salt movement through a Mojave Desert soil. J. Arid Environ. 2008, 72, 1012–1033. [Google Scholar] [CrossRef]
- Orlando, J.; Alfaro, M.; Bravo, L.; Guevara, R.; Caru, M. Bacterial diversity and occurrence of ammonia-oxidizing bacteria in the Atacama Desert soil during a “desert bloom” event. Soil Biol. Biochem. 2010, 42, 1183–1188. [Google Scholar] [CrossRef]
- Olivares, D.; Ferrada, P.; de Matos, C.; Marzo, A.; Cabrera, E.; Portillo, C.; Llanos, J. Characterization of soiling on PV modules in the Atacama Desert. Energy Proced. 2017, 124, 547–553. [Google Scholar] [CrossRef]
- Broadhurst, F.M.; Loring, D.H. Rates of Sedimentation in Upper Carboniferous of Britain. Lethaia 1970, 3, 1. [Google Scholar] [CrossRef]
- Xiao, J.; Porter, S.C.; An, Z.S.; Kumai, H.; Yoshikawa, S. Grain-Size of Quartz as an Indicator of Winter Monsoon Strength on the Loess Plateau of Central China during the Last 130,000-Yr. Quat. Res. 1995, 43, 22–29. [Google Scholar] [CrossRef]
- Quinn, R.C. Experimental Characterization and in situ Measurements of Chemical Processes in the Martian Surface Environment. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2005. [Google Scholar]
- Ewing, S.A.; Macalady, J.L.; Warren-Rhodes, K.; McKay, C.P.; Amundson, R. Changes in the soil C cycle at the arid-hyperarid transition in the Atacama Desert. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Gonzalez-Silva, C.; Arenas-Fajardo, C.; Vicuna, R. Extreme environments as potential drivers of convergent evolution by exaptation: The Atacama Desert Coastal Range case. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Xiao, S.H.; Knoll, A.H.; Yuan, X.L.; Pueschel, C.M. Phosphatized multicellular algae in the Neoproterozoic Doushantuo Formation, China, and the early evolution of florideophyte red algae. Am. J. Bot. 2004, 91, 214–227. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Gonzalez-Silva, C.; Mancilla, R.A.; Salas, L.; Palma, R.E.; Wynne, J.J.; McKay, C.P.; Vicuna, R. Ancient Photosynthetic Eukaryote Biofilms in an Atacama Desert Coastal Cave. Microb. Ecol. 2009, 58, 485–496. [Google Scholar] [CrossRef]
- Paulino-Lima, I.G.; Azua-Bustos, A.; Vicuna, R.; Gonzalez-Silva, C.; Salas, L.; Teixeira, L.; Rosado, A.; Leitao, A.A.D.; Lage, C. Isolation of UVC-Tolerant Bacteria from the Hyperarid Atacama Desert, Chile. Microb. Ecol. 2013, 65, 325–335. [Google Scholar] [CrossRef]
- Paulino-Lima, I.G.; Fujishima, K.; Navarrete, J.U.; Galante, D.; Rodrigues, F.; Azua-Bustos, A.; Rothschild, L.J. Extremely high UV-C radiation resistant microorganisms from desert environments with different manganese concentrations. J. Photoch. Photobio. B 2016, 163, 327–336. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Gonzalez-Silva, C.; Corsini, G. The Hyperarid Core of the Atacama Desert, an Extremely Dry and Carbon Deprived Habitat of Potential Interest for the Field of Carbon Science. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Olivera-Ardid, S.; Klumpp, E.; Knief, C.; Huil, P.W.; Lehndorff, E.; Bol, R. Moisture activation and carbon use efficiency of soil microbial communities along an aridity gradient in the Atacama Desert. Soil Biol. Biochem. 2018, 117, 68–71. [Google Scholar] [CrossRef]
- Williams, L.B.; Ferrell, R.E. Ammonium Substitution in Illite during Maturation of Organic-Matter. Clays Clay Min. 1991, 39, 400–408. [Google Scholar] [CrossRef]
- Schroeder, P.A.; McLain, A.A. Illite-smectites and the influence of burial diagenesis on the geochemical cycling of nitrogen. Clay Min. 1998, 33, 539–546. [Google Scholar] [CrossRef]
- Bebout, G.E.; Fogel, M.L. Nitrogen-Isotope Compositions of Metasedimentary Rocks in the Catalina Schist, California-Implications for Metamorphic Devolatilization History. Geochim. Cosmochim. Acta 1992, 56, 2839–2849. [Google Scholar] [CrossRef]
- Farmer, J.D.; Des Marais, D.J. Exploring for a record of ancient Martian life. J. Geophys. Res. Planet. 1999, 104, 26977–26995. [Google Scholar] [CrossRef]
- Hays, L.E.; Graham, H.V.; Marais, D.J.D.; Hausrath, E.M.; Horgan, B.; McCollom, T.M.; Parenteau, M.N.; Potter-McIntyre, S.L.; Williams, A.J.; Lynch, K.L. Biosignature Preservation and Detection in Mars Analog Environments. Astrobiology 2017, 17, 363. [Google Scholar] [CrossRef]
- Dong, H.L. Clay-Microbe Interactions and Implications for Environmental Mitigation. Elements 2012, 8, 113–118. [Google Scholar] [CrossRef]
- Cook, F.C. Partition of the nitrogen of plant, yeast and meat extracts. J. Am. Chem. Soc. 1914, 36, 1551–1556. [Google Scholar] [CrossRef]
- Keyser, H.H.; Li, F.D. Potential for Increasing Biological Nitrogen-Fixation in Soybean. Plant. Soil 1992, 141, 119–135. [Google Scholar] [CrossRef]
- Glass, C.; Silverstein, J. Denitrification of high-nitrate, high-salinity wastewater. Water Res. 1999, 33, 223–229. [Google Scholar] [CrossRef]
- He, Q.; He, Z.L.; Joyner, D.C.; Joachimiak, M.; Price, M.N.; Yang, Z.K.; Yen, H.C.B.; Hemme, C.L.; Chen, W.Q.; Fields, M.W.; et al. Impact of elevated nitrate on sulfate-reducing bacteria: A comparative Study of Desulfovibrio vulgaris. ISME J. 2010, 4, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- Redding, A.M.; Mukhopadhyay, A.; Joyner, D.C.; Hazen, T.C.; Keasling, J.D. Study of nitrate stress in Desulfovibrio vulgaris Hildenborough using iTRAQ proteomics. Brief. Funct. Genom. 2006, 5, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Vriezen, J.A.C.; de Bruijn, F.J.; Nusslein, K. Responses of rhizobia to desiccation in relation to osmotic stress, oxygen, and temperature. Appl. Environ. Microb. 2007, 73, 3451–3459. [Google Scholar] [CrossRef] [PubMed]
- Connon, S.A.; Lester, E.D.; Shafaat, H.S.; Obenhuber, D.C.; Ponce, A. Bacterial diversity in hyperarid Atacama Desert soils. J. Geophys. Res. Biogeosci. 2007, 112. [Google Scholar] [CrossRef]
- Neilson, J.W.; Califf, K.; Cardona, C.; Copeland, A.; van Treuren, W.; Josephson, K.L.; Knight, R.; Gilbert, J.A.; Quade, J.; Caporaso, J.G.; et al. Significant Impacts of Increasing Aridity on the Arid Soil Microbiome. Msystems 2017, 2. [Google Scholar] [CrossRef]
- Neilson, J.W.; Quade, J.; Ortiz, M.; Nelson, W.M.; Legatzki, A.; Tian, F.; LaComb, M.; Betancourt, J.L.; Wing, R.A.; Soderlund, C.A.; et al. Life at the hyperarid margin: Novel bacterial diversity in arid soils of the Atacama Desert, Chile. Extremophiles 2012, 16, 553–566. [Google Scholar] [CrossRef]
- Drees, K.P.; Neilson, J.W.; Betancourt, J.L.; Quade, J.; Henderson, D.A.; Pryor, B.M.; Maier, R.M. Bacterial community structure in the hyperarid core of the Atacama Desert, Chile. Appl. Environ. Microb. 2006, 72, 7902–7908. [Google Scholar] [CrossRef]
- Warren-Rhodes, K.A.; Rhodes, K.L.; Pointing, S.B.; Ewing, S.A.; Lacap, D.C.; Gomez-Silva, B.; Amundson, R.; Friedmann, E.I.; McKay, C.P. Hypolithic cyanobacteria, dry limit of photosynthesis, and microbial ecology in the hyperarid Atacama Desert. Microb. Ecol. 2006, 52, 389–398. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Gonzalez-Silva, C.; Mancilla, R.A.; Salas, L.; Gomez-Silva, B.; McKay, C.P.; Vicuna, R. Hypolithic Cyanobacteria Supported Mainly by Fog in the Coastal Range of the Atacama Desert. Microb. Ecol. 2011, 61, 568–581. [Google Scholar] [CrossRef]
- Lacap, D.C.; Warren-Rhodes, K.A.; McKay, C.P.; Pointing, S.B. Cyanobacteria and chloroflexi-dominated hypolithic colonization of quartz at the hyper-arid core of the Atacama Desert, Chile. Extremophiles 2011, 15, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Wierzchos, J.; Ascaso, C.; McKay, C.P. Endolithic cyanobacteria in halite rocks from the hyperarid core of the Atacama Desert. Astrobiology 2006, 6, 415–422. [Google Scholar] [CrossRef]
- Dong, H.L.; Rech, J.A.; Jiang, H.C.; Sun, H.; Buck, B.J. Endolithic cyanobacteria in soil gypsum: Occurrences in Atacama (Chile), Mojave (United States), and Al-Jafr Basin (Jordan) deserts. J. Geophys. Res. Biogeosci. 2007, 112. [Google Scholar] [CrossRef]
- Orlando, J.; Caru, M.; Pommerenke, B.; Braker, G. Diversity and activity of denitrifiers of Chilean arid soil ecosystems. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Prado, B.; Delmoral, A.; Quesada, E.; Rios, R.; Monteolivasanchez, M.; Campos, V.; Ramoscormenzana, A. Numerical Taxonomy of Moderately Halophilic Gram-Negative Rods Isolated from the Salar-De-Atacama, Chile. Syst. Appl. Microbiol. 1991, 14, 275–281. [Google Scholar] [CrossRef]
- Lizama, C.; Monteoliva-Sanchez, M.; Prado, B.; Ramos-Cormenzana, A.; Weckesser, J.; Campos, V. Taxonomic study of extreme halophilic archaea isolated from the “Salar de Atacama”, Chile. Syst. Appl. Microbiol. 2001, 24, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Lizama, C.; Monteoliva-Sanchez, M.; Suarez-Garcia, A.; Rosello-Mora, R.; Aguilera, M.; Campos, V.; Ramos-Cormenzana, A. Halorubrum tebenquichense sp nov., a novel halophilic archaeon isolated from the Atacama Saltern, Chile. Int. J. Syst. Evol. Microbiol. 2002, 52, 149–155. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Lati-Tude | Longi-Tude | Alti-Tude (m) | Annual Precipi-Tation (mm/yr) | Modeled Precipitation (mm/day) (6–7 June 2017) | Median Grain Size (μm) |
|---|---|---|---|---|---|---|
| MES | 22.2641° S | 69.7243° W | 1493 ± 8 | 0.7 | 3.3 | 444 |
| PONR-2 | 23.0726° S | 69.5892° W | 1493 ± 8 | 1.1 | 10.1 | 486 |
| Yungay | 24.0884° S | 69.9945° W | 1007 ± 10 | 2.0 | 13.4 | 464 |
| TZ-0 | 26.3222° S | 70.0128° W | 1106 ± 10 | 10.0 ± 0.5 | 2.9 | 493 |
| TZ-4 | 27.0565° S | 69.9228° W | 1658 ± 10 | 15.1 ± 1.1 | 3.9 | 291 |
| TZ-5 | 27.6051° S | 70.4458° W | 588 ± 8 | 22.7 ± 5.3 | 0.4 | 322 |
| TZ-6 | 28.4100° S | 70.7270° W | 658 ± 6 | 31.2 ± 1.1 | 0.1 | 416 |
| Pit Name | pH | Conductivity (mS/cm) | NO3− (ppm) | NH4+ (ppm) | TOC (ppm) | TN (ppm) | SiO2/Al2O3 |
|---|---|---|---|---|---|---|---|
| M1 | 8.96 ± 0.00 | 1.41 ± 0.07 | 35.7 ± 0.3 | <0.5 | 49.1 ± 1.7 | 18.0 ± 0.4 | 4.65 |
| M2 | 8.29 ± 0.02 | 8.69 ± 0.08 | 238.4 ± 3.6 | <0.5 | 82.0 ± 4.5 | 45.2 ± 1.1 | 4.18 |
| M3 | 8.38 ± 0.01 | 9.69 ± 0.09 | 52.1 ± 0.2 | <0.5 | 47.0 ± 2.8 | 24.1 ± 1.9 | 4.59 |
| P1 | 8.29 ± 0.01 | 12.97 ± 0.03 | 147.5 ± 2.5 | <0.5 | 82.5 ± 1.3 | 46.2 ± 0.2 | 4.21 |
| P2 | 8.20 ± 0.01 | 22.68 ± 0.11 | 362.0 ± 39.1 | <0.5 | 102.9 ± 2.7 | 94.2 ± 1.9 | 3.85 |
| P3 | 8.09 ± 0.00 | 22.10 ± 0.12 | 157.7 ± 2.6 | <0.5 | 97.1 ± 1.1 | 43.5 ± 0.9 | 3.84 |
| Y1 | 8.03 ± 0.00 | 20.45 ± 0.20 | 23.8 ± 0.1 | <0.5 | 73.3 ± 0.6 | 43.2 ± 0.1 | 3.87 |
| Y2 | 8.09 ± 0.00 | 17.14 ± 0.07 | 19.3 ± 1.0 | <0.5 | 71.7 ± 1.6 | 36.4 ± 1.4 | 3.96 |
| Y3 | 8.11 ± 0.00 | 17.66 ± 0.16 | 15.5 ± 0.3 | <0.5 | 83.9 ± 3.8 | 45.5 ± 2.1 | 4.06 |
| T01 | 9.29 ± 0.01 | 0.86 ± 0.01 | 1.9 ± 0.0 | <0.5 | 117.6 ± 2.1 | 37.7 ± 0.6 | 3.91 |
| T02 | 9.37 ± 0.01 | 0.89 ± 0.06 | 1.4 ± 0.0 | <0.5 | 140.6 ± 4.9 | 51.6 ± 0.4 | 4.13 |
| T03 | 9.83 ± 0.00 | 1.06 ± 0.06 | 2.5 ± 0.0 | <0.5 | 106.1 ± 4.2 | 35.6 ± 0.3 | 3.77 |
| T41 | 8.39 ± 0.01 | 13.62 ± 0.38 | 2686.8 ± 141.6 | <0.5 | 220.8 ± 7.4 | 84.1 ± 1.2 | 3.76 |
| T42 | 8.07 ± 0.01 | 26.02 ± 0.48 | 6802.6 ± 290.2 | <0.5 | 242.5 ± 2.0 | 103.7 ± 0.9 | 3.55 |
| T43 | 8.26 ± 0.00 | 17.20 ± 0.28 | 4100.0 ± 160.4 | <0.5 | 170.6 ± 2.3 | 77.4 ± 0.8 | 3.66 |
| T51 | 9.60 ± 0.00 | 0.82 ± 0.06 | 3.0 ± 0.1 | <0.5 | 708.7 ± 14.8 | 82.9 ± 1.8 | 4.49 |
| T52 | 8.96 ± 0.01 | 4.08 ± 0.13 | 6.0 ± 0.1 | <0.5 | 674.9 ± 7.2 | 79.0 ± 0.7 | 4.44 |
| T53 | 8.79 ± 0.00 | 20.24 ± 0.45 | 20.9 ± 0.2 | <0.5 | 985.3 ± 2.4 | 105.7 ± 0.5 | 4.47 |
| T61 | 9.46 ± 0.00 | 1.69 ± 0.09 | 11.7 ± 0.2 | <0.5 | 1089.9 ± 12.3 | 145.9 ± 1.3 | 4.23 |
| T62 | 8.60 ± 0.01 | 16.23 ± 0.35 | 140.6 ± 1.3 | <0.5 | 1576.1 ± 21.4 | 176.7 ± 3.0 | 4.36 |
| T63 | 9.20 ± 0.00 | 6.07 ± 0.14s | 82.4 ± 0.8 | <0.5 | 1407.2 ± 30.7 | 173.6 ± 2.5 | 4.32 |
| Type of Culture Plate | Amendment | M3 | P3 | Y3 | T03 | T43 | T53 | T63 |
|---|---|---|---|---|---|---|---|---|
| Ultrapure agarose | None | 0 | 17 | 13 | 93 | 1.62 × 103 | 80 | 1.42 × 103 |
| Water | 0 | 15 | 106 | 111 | 3.57 × 103 | 87 | 667 | |
| Nitrate | 2 | 2 | 7 | 1.33 × 103 | 3.15 × 103 | 1.62 × 103 | 2.93 × 103 | |
| Tryptic soy agar | None | 37 | 53 | 169 | 514 | 1.60 × 105 | 1.33 × 105 | 3.21 × 105 |
| Water | 5.76 × 103 | 3.39 × 103 | 1.14 × 104 | 312 | 1.65 × 106 | 5.22 × 104 | 5.09 × 105 | |
| Nitrate | 12 | 1.48 × 104 | 5.20 × 104 | 123 | 8.41 × 104 | 7.61 × 104 | 4.38 × 105 | |
| LB agar | None | 17 | 28 | 33 | 91 | 2.39 × 105 | 2.95 × 103 | 5.41 × 104 |
| Water | 7 | 7.98 × 103 | 3.71 × 103 | 491 | 1.23 × 106 | 1.17 × 103 | 1.54 × 105 | |
| Nitrate | 0 | 9.43 × 103 | 197 | 290 | 4.88 × 103 | 1.00 × 104 | 3.27 × 105 | |
| Plate count agar | None | 167 | 190 | 5.17 × 103 | 353 | 2.90 × 106 | 6.00 × 103 | 2.72 × 105 |
| Water | 1.36 × 103 | 8.56 × 104 | 6.96 × 104 | 638 | 1.33 × 107 | 1.25 × 105 | 4.12 × 106 | |
| Nitrate | 15 | 44 | 29 | 343 | 2.47 × 106 | 6.96 × 104 | 5.22 × 105 |
| Pit Name | Ultrapure Agarose Plates | Tryptic Soy Agar Plates | LB Agar Plates | Plate Count Agar Plates |
|---|---|---|---|---|
| M3 | 0.38 | −2.68 ** | −0.86 | −1.97 * |
| P3 | −0.78 | 0.64 | 0.07 | −3.29 *** |
| Y3 | −1.17 * | 0.66 | −1.28 * | −3.38 *** |
| T03 | 1.08 | −0.40 | −0.23 | −0.27 |
| T43 | −0.05 | −1.29 * | −2.40 ** | −0.73 |
| T53 | 1.27 | 0.16 | 0.93 | −0.25 |
| T63 | 0.64 | −0.07 | 0.33 | −0.90 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Zerkle, A.L.; Stueeken, E.; Claire, M.W. Nitrates as a Potential N Supply for Microbial Ecosystems in a Hyperarid Mars Analog System. Life 2019, 9, 79. https://doi.org/10.3390/life9040079
Shen J, Zerkle AL, Stueeken E, Claire MW. Nitrates as a Potential N Supply for Microbial Ecosystems in a Hyperarid Mars Analog System. Life. 2019; 9(4):79. https://doi.org/10.3390/life9040079
Chicago/Turabian StyleShen, Jianxun, Aubrey L. Zerkle, Eva Stueeken, and Mark W. Claire. 2019. "Nitrates as a Potential N Supply for Microbial Ecosystems in a Hyperarid Mars Analog System" Life 9, no. 4: 79. https://doi.org/10.3390/life9040079
APA StyleShen, J., Zerkle, A. L., Stueeken, E., & Claire, M. W. (2019). Nitrates as a Potential N Supply for Microbial Ecosystems in a Hyperarid Mars Analog System. Life, 9(4), 79. https://doi.org/10.3390/life9040079