A Statistical Approach to Illustrate the Challenge of Astrobiology for Public Outreach



Abstract
1. Introduction
2. General Overview and Probabilistic Concept
2.1. On the Conditions Required for Life to Appear
2.2. On the Probability of the Occurrence of Extraterrestrial Life
2.3. The More Complex, the Less Probable
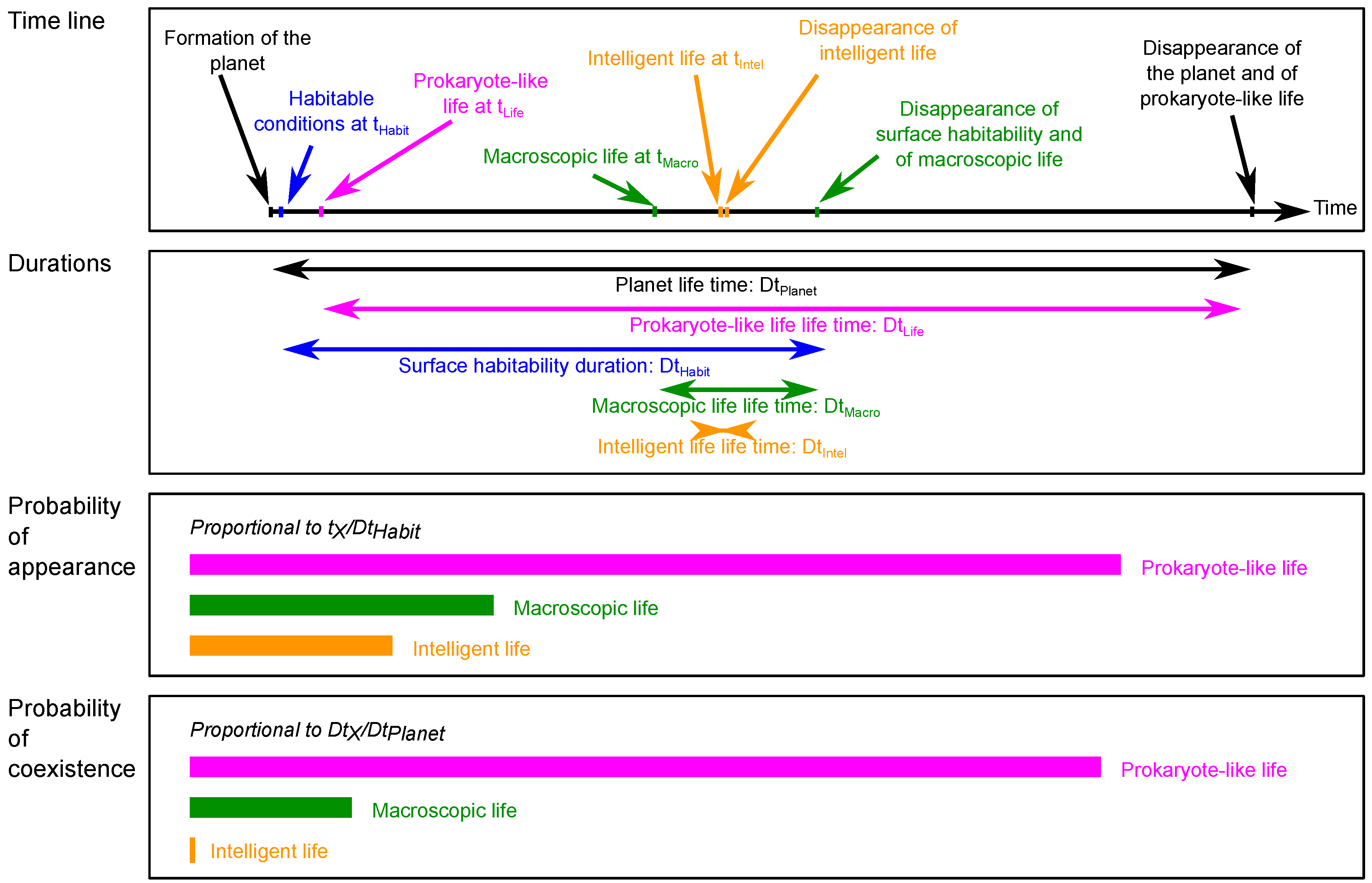
3. Equations for Life in the Universe
3.1. Probability Equation for Life in the Universe
- : the proportion of stellar systems having a star compatible with the occurrence of the considered stage of life.
- : the proportion of these stellar systems having at least one rocky body located at a distance from the star compatible with the considered stage of life, i.e., within its habitable zone.
- : the proportion of these bodies compatible with the emergence of life.
- : the probability that life appeared on these bodies.
- : the probability that life reached the considered evolutionary stage on these bodies.
- : the probability that life at this evolutionary stage is still active on these bodies. This is equivalent to the probability of co-existence occurrence of this stage of life on several bodies simultaneously.
3.2. Converting Probabilities into Distances
4. Solving the Equations
4.1. Astronomical Terms
4.2. Empirical Terms
4.3. Results
4.3.1. Prokaryote-Like Unicellular Life
4.3.2. Macroscopic Multicellular Life
4.3.3. Intelligent Civilisations
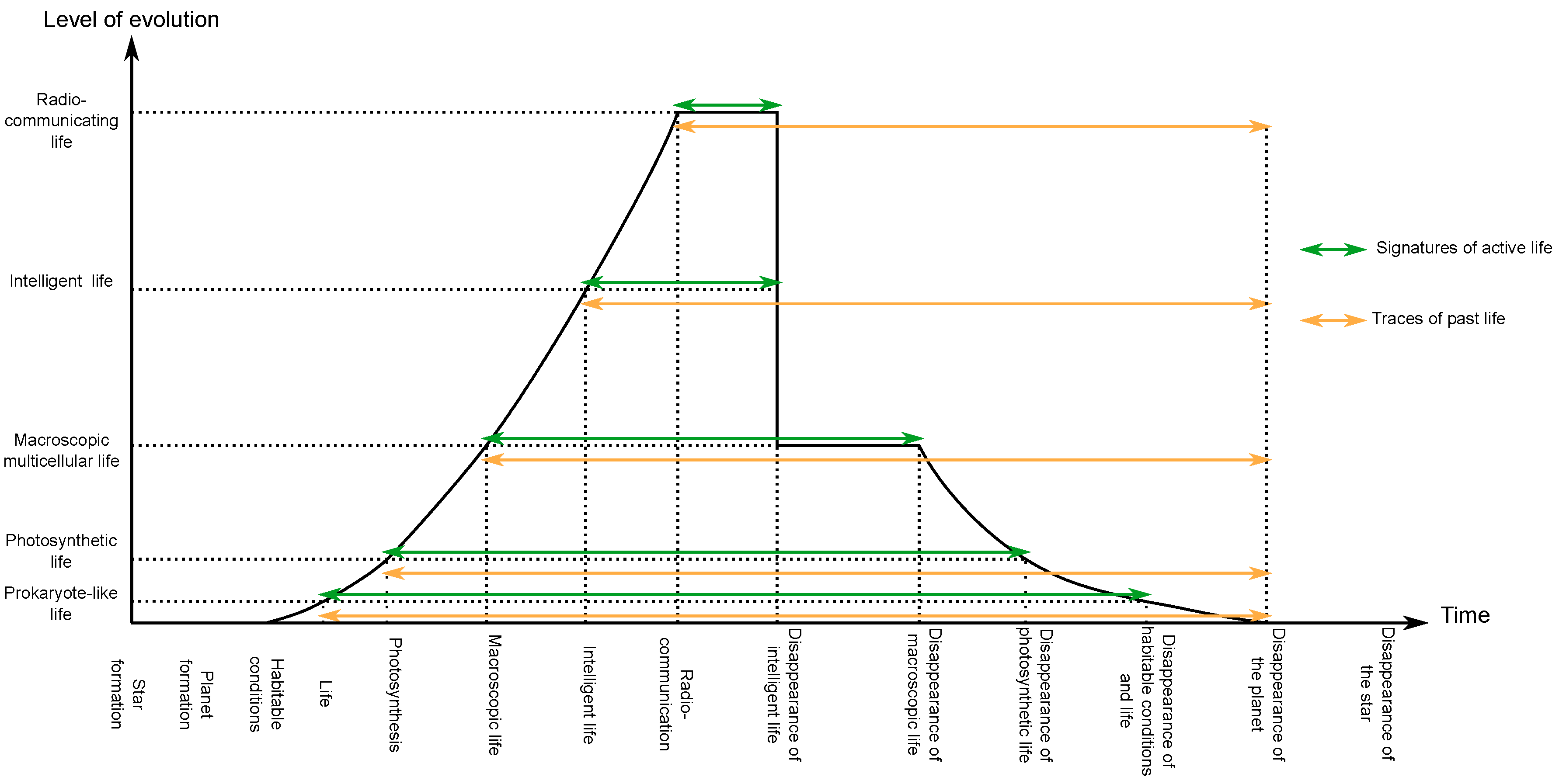
4.3.4. Considering Past Life
4.4. Summary
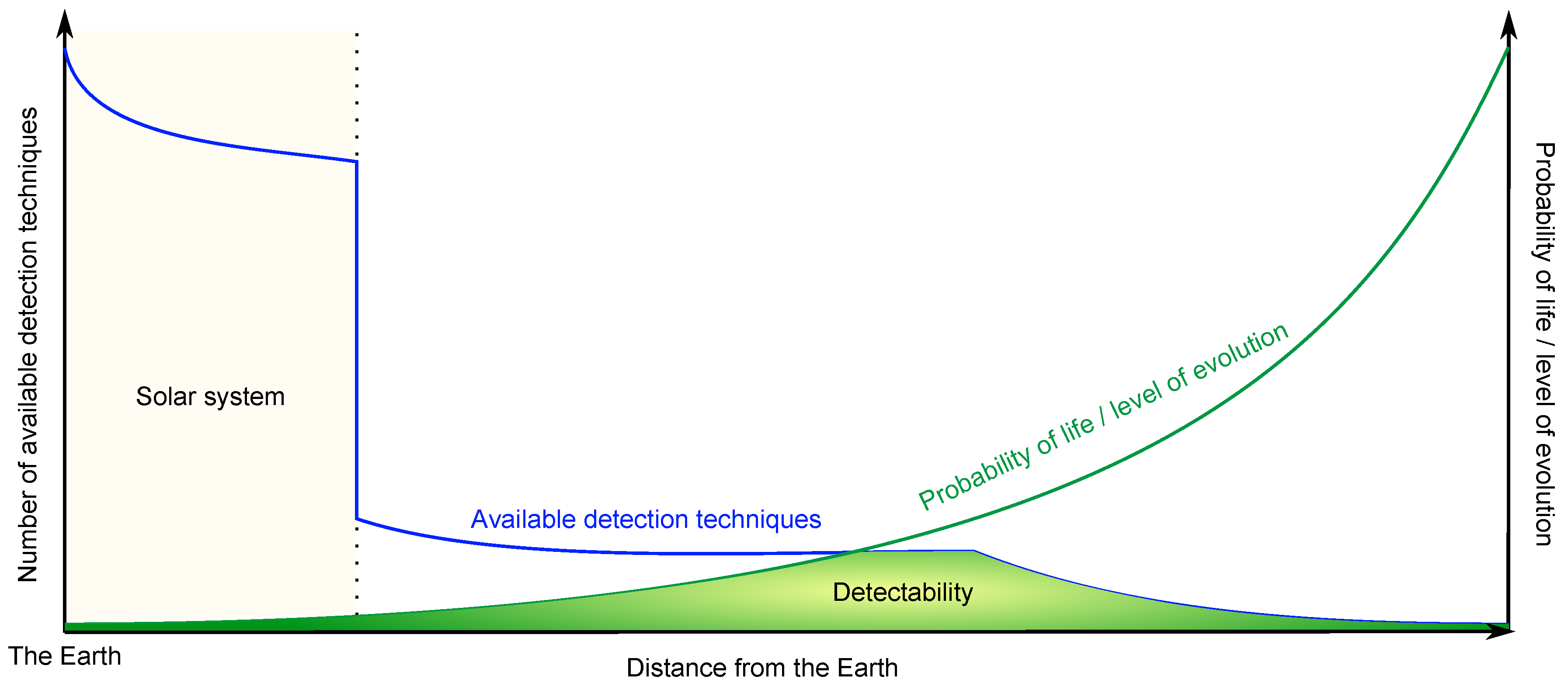
5. On the Detection of Extraterrestrial Life
5.1. Search for Extraterrestrial Prokaryote-Like Life
5.2. Search for Macroscopic Multicellular Life
5.3. Search for Extraterrestrial Civilisations
5.4. Search for Past Extraterrestrial Civilisations
5.5. Summary
6. From Complex Modeling to General Public Outreach
6.1. Describing Important Astronomical Notions
6.2. Introducing the Probabilistic Approach
6.3. From Intelligent to Microbial E.T.
6.4. Testing the Approach with Teachers
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brack, A.; Horneck, G.; Cockell, B.A.; Belisheva, N.K.; Eiroa, C.; Henning, T.; Herbst, T.; Kaltenegger, L.; Léger, A.; Liseau, R.; et al. Origin and evolution of life on terrestrial planets. Astrobiology 2010, 10, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Popa, R. Advances in astrobiology and biogeophysics. In Between Necessity and Probablility: Searching for the Definitions and Origin of Life; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Lazcano, A. Origin of life. In Encyclodedia of Astrobiology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1183–1190. [Google Scholar]
- Ruiz-Mirazo, K.; Moreno, A. Life. In Encyclodedia of Astrobiology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 919–921. [Google Scholar]
- Brack, A. Astrobiology. In The Quest for the Conditions of Life; Springer: Berlin/Heidelberg, Germany, 2001; pp. 79–88. [Google Scholar]
- Miller, S.L. Production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Brack, A. Avant la vie, la chimie prebiotique. Biofutur 2013, 345, 34–37. [Google Scholar]
- De Ronde, C.E.J.; Ebbesen, W. 3.2 b.y. of organic compound formation near sea-floor hot springs. Oceanogr. Lit. Rev. 1996, 24, 791–794. [Google Scholar] [CrossRef]
- Dickens, J.; Irvine, W.; Nummelin, A.; Mollendal, H.; Saito, S.; Thorwirth, S.; Hjalmarson, A.; Ohishi, M. Searches for new interstellar molecules, including a tentative detection of aziridine and a possible detection of propenal. Spectrochim. Acta Part A 2001, 57, 643–660. [Google Scholar] [CrossRef]
- Danger, G.; Orthous-Daunay, F.-R.; de Marcellus, P.; Modica, P.; Vuitton, V.; Duvernay, F.; Flandinet, L.; d’Hendecourt, L.L.S.; Thissen, R.; Chiavassa, T. Characterization of laboratory analogs of interstellar cometary organic residues using very high resolution mass spectrometry. Geochim. Cosmochim. Acta 2013, 118, 184–201. [Google Scholar] [CrossRef]
- Meinert, C.; Myrgorodska, I.; de Marcellus, P.; Buhse, T.; Nahon, L.; Hoffmann, S.V.; d’Hendecourt, L.L.S.; Meierhenrich, U.J. Ribose and related sugars from ultraviolet irradiation of interstellar ice analogs. Science 2016, 352, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Altwegg, K.; Balsiger, H.; Bar-Nun, A.; Berthelier, J.-J.; Bieler, A.; Bochsler, P.; Briois, C.; Calmonte, U.; Combi, M.R.; Cottin, H.; et al. Prebiotic chemicals-amino acid and phosphorus-in the coma of comet 67P/Churyumov-Gerasimenko. Sci. Adv. 2016, 2, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pizzarello, S.; Shock, E. Carbonaceous chondrite meteorites: The chronicle of a potential evolutionary path between stars and life. Orig. Life Evol. Biosph. 2017, 47, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 7, 2763–2768. [Google Scholar] [CrossRef] [PubMed]
- Arrhenius, S. On the propagation of life in space. Quest Extraterr. Life 1903, 7, 481–485. [Google Scholar]
- Brack, A. Liquid water and the origin of life. Orig. Life Evol. Biosph. 1993, 23, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Brack, A. The importance of water for life. In Water and Planets; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Dass, A.V.; Hickman-Lewis, K.; Brack, A.; Kee, T.P.; Westall, F. Stochastic prebiotic chemistry within realistic geological systems. Chem. Sel. Rev. 2016, 1, 1–22. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Irwin, L.N. The prospect of alien life in exotic forms on other worlds. Naturwissenschaften 2006, 93, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Drake, F. The radio search for intelligent extraterrestrial life. In Current Aspects of Exobiology; Mamikunian, G., Briggs, M.H., Eds.; Pergamon Press: New York, NY, USA, 1965; pp. 323–345. [Google Scholar]
- Cirkovic, M.M. The temporal aspect of the drake equation and SETI. Astrobiology 2004, 4, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.D. Broadcasting but not receiving: Density dependence considerations for SETI signals. Int. J. Astrobiol. 2009, 8, 101–105. [Google Scholar] [CrossRef]
- Forgan, D.H. A numerical testbed for hypotheses of extraterrestrial life and intelligence. Int. J. Astrobiol. 2009, 8, 121–131. [Google Scholar] [CrossRef]
- Grosz, I. Number of galactic civilisations. Open Astron. J. 2013, 6, 72–76. [Google Scholar] [CrossRef]
- Maccone, C. A mathematical model for evolution and SETI. Orig. Life Evol. Biosph. 2011, 41, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Maccone, C. SETI and SHE (Statistical Equation for Habitables). Acta Astronaut. 2011, 68, 63–75. [Google Scholar] [CrossRef]
- Maccone, C. SETI, Evolution and human history merged into a mathematical model. Int. J. Astrobiol. 2013, 12, 218–245. [Google Scholar] [CrossRef]
- Maccone, C. Evolution and history in a new “mathematical SETI” model. Acta Astronaut. 2014, 93, 317–344. [Google Scholar] [CrossRef]
- Seager, S. The drake equation revisited: Interview with planet hunter sara seager, devin powell. Astrobiol. Mag. 2013. Available online: https://www.space.com/22648-drake-equation-alien-life-seager.html (accessed on 23 October 2017).
- Seager, S. Exoplanet habitability. Science 2013, 340, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Waltham, D. Star masses and star-planet distances for Earth-like habitability. Astrobiology 2017, 17, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Loizeau, D.; Foucher, F.; Bost, N.; Bertrand, M.; Vago, J.; Kminek, G. Habitability on mars from a microbial point of view. Astrobiology 2013, 13, 887–897. [Google Scholar] [CrossRef] [PubMed]
- McKay, C.P. An approach to searching for life on Mars, Europa, and Enceladus. Space Sci. Rev. 2008, 135, 49–54. [Google Scholar] [CrossRef]
- Rampelotto, P.H. Resistance of microorganisms to extreme environmental conditions and Its contribution to astrobiology. Sustainability 2010, 2, 1602–1623. [Google Scholar] [CrossRef]
- Lunine, J.I.; Waite, J.H.; Postberg, F.; Spilker, L.; Clark, K. Enceladus life finder: The search for life in a habitable moon. In Proceedings of the 2016 IEEE Aerospace Conference, Brunów, Poland, 27 June–1 July 2016; IEEE Computer Society: Washington, WA, USA, 2016. [Google Scholar]
- Thomson, D’A.W. On Growth and Form; Cambridge University Press: Cambridge, UK, 1917. [Google Scholar]
- Kasting, J. How to Find a Habitable Planet; Princeton University Press: Princeton, NJ, USA, 2012. [Google Scholar]
- Gowanlock, M.G.; Patton, D.R.; McConnell, S.M. A model of habitability within the milky way galaxy. Astrobiology 2011, 11, 855–873. [Google Scholar] [CrossRef] [PubMed]
- Cassan, A.; Kubas, D.; Beaulieu, J.-P.; Dominik, M.; Horne, K.; Greenhill, J.; Wambsganss, J.; Menzies, J.; Williams, A.; Jorgensen, U.G.; et al. One or more bound planets per milky way star from microlensing observations. Nature 2012, 481, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, J.; Schneider, J. Detecting companions to extrasolar planets using mutual events. Astron. Astrophys. 2007, 464, 1133–1138. [Google Scholar] [CrossRef]
- Jones, M.I.; Jenkins, J.S.; Bluhm, P.; Rojo, P.; Melo, C.H.F. The properties of planets around giant stars. Astron. Astrophys. 2014, 566, A113. [Google Scholar] [CrossRef]
- Holland, H.D. The oxygenation of the atmosphere and oceans. Philos. Trans. R. Soc. B 2006, 361, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Petigura, E.A.; Howard, A.W.; Marcy, G.W. Prevalence of Earth-size planets orbiting Sun-like stars. Proc. Natl. Acad. Sci. USA 2013, 110, 19273–19278. [Google Scholar] [CrossRef] [PubMed]
- Huggett, R.J. Geoecology: An Evolutionary Approach; Routledge: Abingdon, UK, 1995. [Google Scholar]
- Lammer, H.; Bredehöft, J.H.; Coustenis, A.; Khodachenko, M.L.; Kaltenegger, L.; Grasset, O.; Prieur, D.; Raulin, F.; Ehrenfreund, P.; Yamauchi, M.; et al. What makes a planet habitable? Astron. Astrophys. Rev. 2009, 17, 181–249. [Google Scholar] [CrossRef]
- Spiegel, D.S.; Turner, E.L. Bayesian analysis of the astrobiological implications of life’s early emergence on Earth. Proc. Natl. Acad. Sci. USA 2012, 109, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S. Vacant habitats in the universe. Trends Ecol. Evol. 2011, 26, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S. Habitable worlds with no signs of life. Philos. Trans. A Math. Phys. Eng. Sci. 2014, 372. [Google Scholar] [CrossRef] [PubMed]
- Mojzsis, S.J.; Harrison, T.M.; Pidgeon, R.T. Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s surface 4300 Myr ago. Nature 2001, 409, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Schröder, K.-P.; Connon Smith, R. Distant future of the Sun and Earth revisited. Mon. Not. R. Astron. Soc. 2008, 386, 155–163. [Google Scholar] [CrossRef]
- El Albani, A.E.; Bengtson, S.; Canfield, D.E.; Bekker, A.; Macchiarelli, R.; Mazurier, A.; Hammarlund, E.U.; Boulvais, P.; Dupuy, J.-J.; Fontaine, C.; et al. Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago. Nature 2010, 466, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, D. Lessons from the past: Evolutionary impacts of mass extinctions. Proc. Natl. Acad. Sci. USA 2001, 98, 5393–5398. [Google Scholar] [CrossRef] [PubMed]
- Hull, P. Life in the aftermath of mass extinctions. Curr. Biol. 2015, 25, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Foucher, F.; Bost, N.; Bertrand, M.; Loizeau, D.; Vago, J.L.; Kminek, G.; Gaboyer, F.; Campbell, K.A.; Bréhéret, J.G.; et al. Biosignatures on Mars: What, where and how? implications for the search for Martian life. Astrobiology 2015, 15, 998–1029. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, J.L.; Rauer, H.; Selsis, F.; Kaltenegger, L.; Beichman, C.; Danchi, W.; Eiroa, C.; Fridlund, M.; Henning, T.; Herbst, T.; et al. Co-evolution of astmospheres, life, and climate. Astrobiology 2010, 10, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Léger, A.; Fontecave, M.; Labeyrie, A.; Samuel, B.; Demangeon, O.; Valencia, D. Is the presence of oxygen on an exoplanet a reliable biosignature? Astrobiology 2011, 11, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Segura, A.; Meadows, V.S.; Kasting, J.F.; Crisp, D.; Cohen, M. Abiotic formation of O2 and O3 in high-CO2 terrestrial atmospheres. Astron. Astrophys. 2007, 472, 665–679. [Google Scholar] [CrossRef]
- Buick, R. When did oxygenic photosynthesis evolve? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2731–2743. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W. The paleobiological record of photosynthesis. Photosynth. Res. 2011, 107, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Mandell, A.M.; Haynes, K.; Sinukoff, E.; Madhusudhan, N.; Burrows, A.; Deming, D. Exoplanet transit spectroscopy using WFC3: WASP-12 b, WASP-17 b, and WASP-19 b. Astrophys. J. 2013, 779, 1–18. [Google Scholar] [CrossRef]
- Des Marais, D.J. Exploring mars for evidence of habitable environments and life. Proc. Am. Philos. Soc. 2010, 154, 402–421. [Google Scholar]
- Seager, S.; Turner, E.L.; Schafer, J.; Ford, E.B. Vegetation’s red edge: A possible spectroscopic biosignature of extraterrestrial plants. Astrobiology 2005, 5, 372–390. [Google Scholar] [CrossRef] [PubMed]
- Kaltenegger, L.; Selsis, F. Characterizing habitable extrasolar planets using spectral fingerprints. C. R. Palevol 2009, 2009, 1–14. [Google Scholar] [CrossRef]
- Cocconi, G.; Morrison, P. Searching for interstellar communications. Nature 1959, 184, 844–846. [Google Scholar] [CrossRef]
- Schwartz, R.N.; Townies, C.H. Interstellar and interplanetary communication by optical Masers. Nature 1961, 190, 205–208. [Google Scholar] [CrossRef]
- Lin, H.W.; Abad, G.G.; Loeb, A. Detecting industrial pollution in the atmospheres of Earth-like exoplanets. Astrophys. J. Lett. 2014, 792, L7. [Google Scholar] [CrossRef]
- Osamnov, Z. Are the dyson rings around pulsars detectable? Int. J. Astrobio. 2017, in press. [Google Scholar] [CrossRef]
- Cabrol, N.A. Alien mindscapes—A perspective on the search for extraterrestrial intelligence. Astrobiology 2016, 16, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.; Forgan, D.; O’Malley James, J. Observational signatures of self-destructive civilisations. Int. J. Astrobiol. 2016, 15, 333–344. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Type of Life | (%) | (ly) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Prokaryote-like life | 1.00 | 1.00 | 0.22 | 0.17 | 1.00 | 0.90 | 3.36% | 6.73 × 109 | 19 |
| Macroscopic multicellular life | 0.10 | 0.22 | 0.75 | 0.94 | 0.28 | 0.16 | 0.07% | 1.39 × 108 | 70 |
| Present civilisation | 0.10 | 0.22 | 0.75 | 0.94 | 0.19 | 0.00002 | 0.000006% | 11,788 | 1586 |
| Past macroscopic multicellular life | 0.10 | 0.22 | 0.75 | 0.94 | 0.28 | 0.56 | 0.24% | 4.86 × 108 | 46 |
| Past civilisation | 0.10 | 0.22 | 0.75 | 0.94 | 0.19 | 0.50 | 0.15% | 2.95 × 108 | 54 |
| Type of Life | (%) | (ly) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Extant extraterrestrial | ||||||||||
| radio-communicating | 0.10 | 0.22 | 0.75 | 0.94 | 0.19 | 1 × 108 | N.A. | 2.9 | 6 | 19,979 |
| life | ||||||||||
| Past extraterrestrial | ||||||||||
| self-destructing | 0.10 | 0.22 | 0.75 | 0.94 | 0.19 | 1 × 105 | N.A. | 2.9 × 10−6% | 5894 | 1998 |
| civilisation | ||||||||||
| Vegetation | 0.10 | 0.22 | 0.75 | 0.94 | 0.23 | 0.14 | N.A. | 0.05% | 10 × 107 | 78 |
| Photosynthetic life | 0.10 | 0.22 | 0.75 | 0.94 | 0.64 | 0.34 | N.A. | 0.34% | 6.75 × 108 | 41 |
| Extraterrestrial | ||||||||||
| microbial life | 1.00 | 1.00 | 0.22 | 0.17 | 1.00 | 0.90 | 0.337 | 0.11% | N.A. | N.A. |
| in the Solar System | ||||||||||
| Microfossils on Mars | 1.00 | 0.22 | 0.75 | 0.94 | 1.00 | 0.95 | 0.337 | 0.05% | N.A. | N.A. |
| If the Sun Had the Size of | True Distance | If the Sun Had the Size of | ||
|---|---|---|---|---|
| Diameter in km | a Tennis Ball, the Diameter | from the Sun | a Tennis Ball, the Distance | |
| Would Be in mm | in km | Would Be in m | ||
| Sun | 1,395,200 | 65 | 0 | 0.0 |
| Mercury | 4900 | 0.23 | 58,000,000 | 2.7 |
| Venus | 12,000 | 0.56 | 108,000,000 | 5.0 |
| The Earth | 12,800 | 0.60 | 150,000,000 | 7.0 |
| The Moon | 3474 | 0.16 | 150,000,000 | 7.0 |
| Mars | 6800 | 0.32 | 228,000,000 | 10.6 |
| Jupiter | 140,000 | 6.52 | 780,000,000 | 36.3 |
| Saturn | 120,000 | 5.59 | 1,430,000,000 | 66.6 |
| Uranus | 52,000 | 2.42 | 2,880,000,000 | 134.2 |
| Neptune | 50,000 | 2.33 | 4,497,000,000 | 209.5 |
| Proxima Centauri | 200,000 | 9.32 | 42,430,000,000,000 | 1977 km |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foucher, F.; Hickman-Lewis, K.; Westall, F.; Brack, A. A Statistical Approach to Illustrate the Challenge of Astrobiology for Public Outreach. Life 2017, 7, 40. https://doi.org/10.3390/life7040040
Foucher F, Hickman-Lewis K, Westall F, Brack A. A Statistical Approach to Illustrate the Challenge of Astrobiology for Public Outreach. Life. 2017; 7(4):40. https://doi.org/10.3390/life7040040
Chicago/Turabian StyleFoucher, Frédéric, Keyron Hickman-Lewis, Frances Westall, and André Brack. 2017. "A Statistical Approach to Illustrate the Challenge of Astrobiology for Public Outreach" Life 7, no. 4: 40. https://doi.org/10.3390/life7040040
APA StyleFoucher, F., Hickman-Lewis, K., Westall, F., & Brack, A. (2017). A Statistical Approach to Illustrate the Challenge of Astrobiology for Public Outreach. Life, 7(4), 40. https://doi.org/10.3390/life7040040