Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Physical Environment on ISS, the Earth, Moon and Mars
2.2. Main Conclusion from Literature Review
2.2.1. Gravity
2.2.2. Radiation
2.2.3. Magnetic Fields
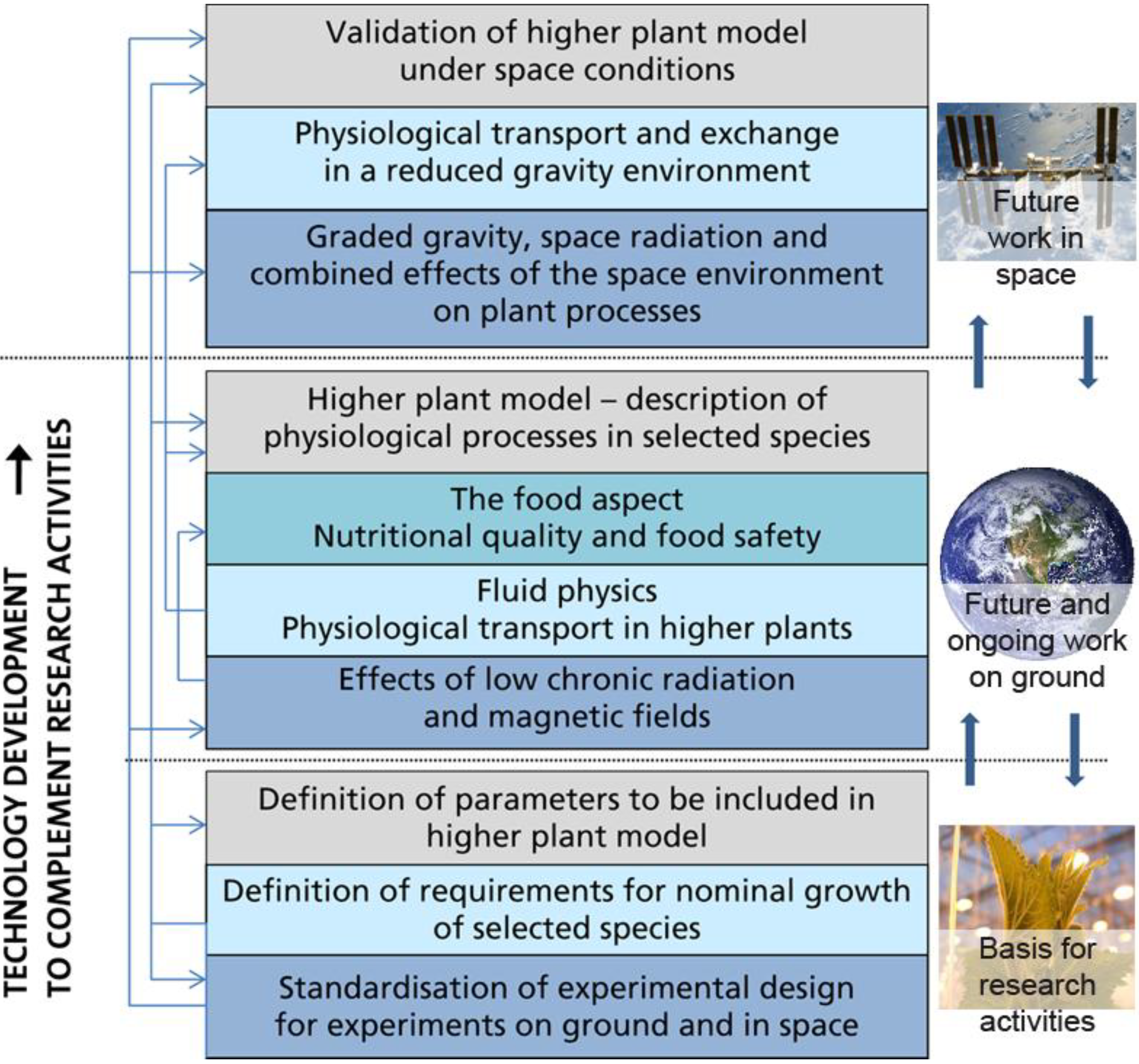
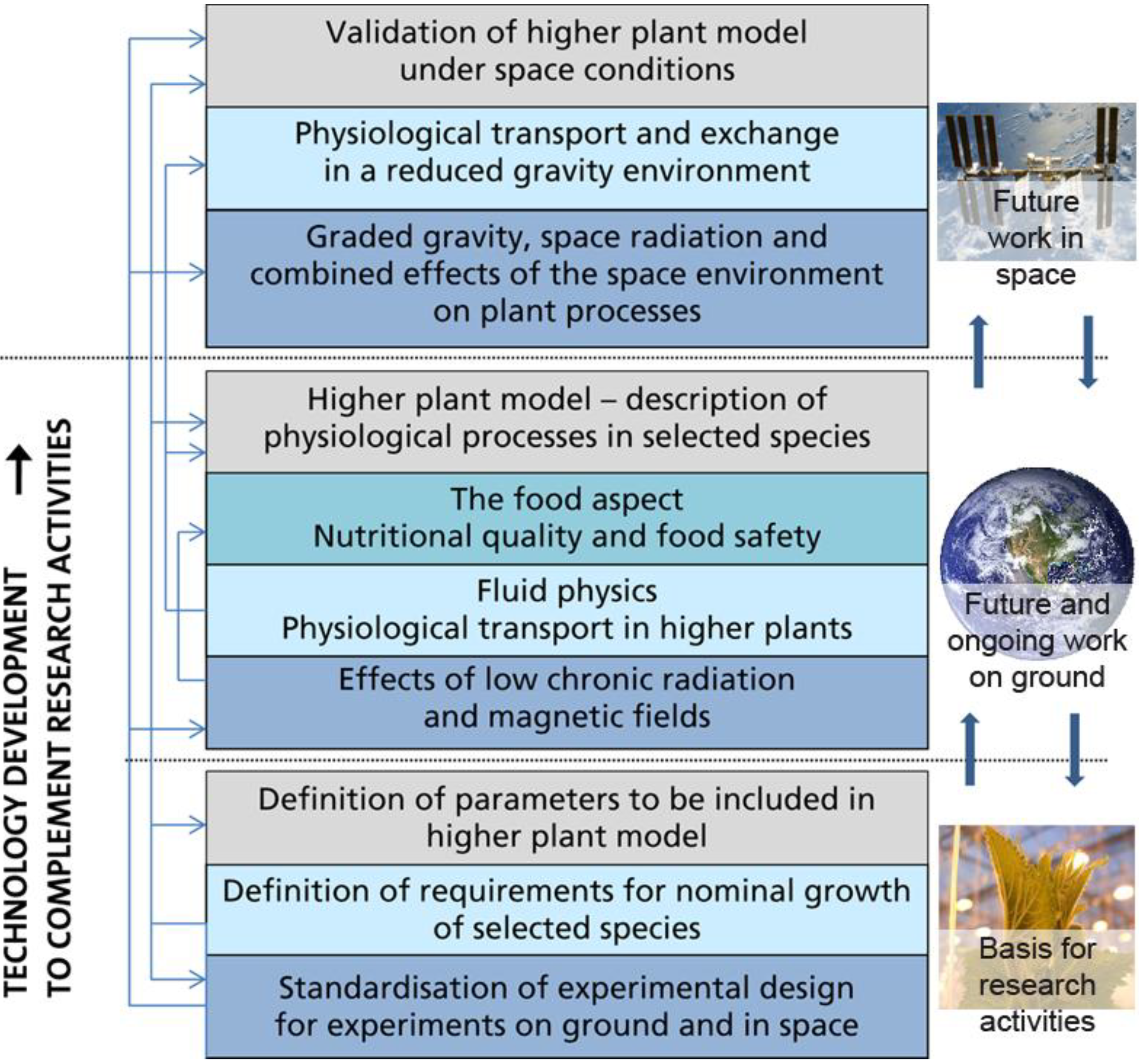
3. Directions and Requirements for Future Research
3.1. Higher Plant Model
3.2. Ground Based Experiments
3.3. Space Experiments
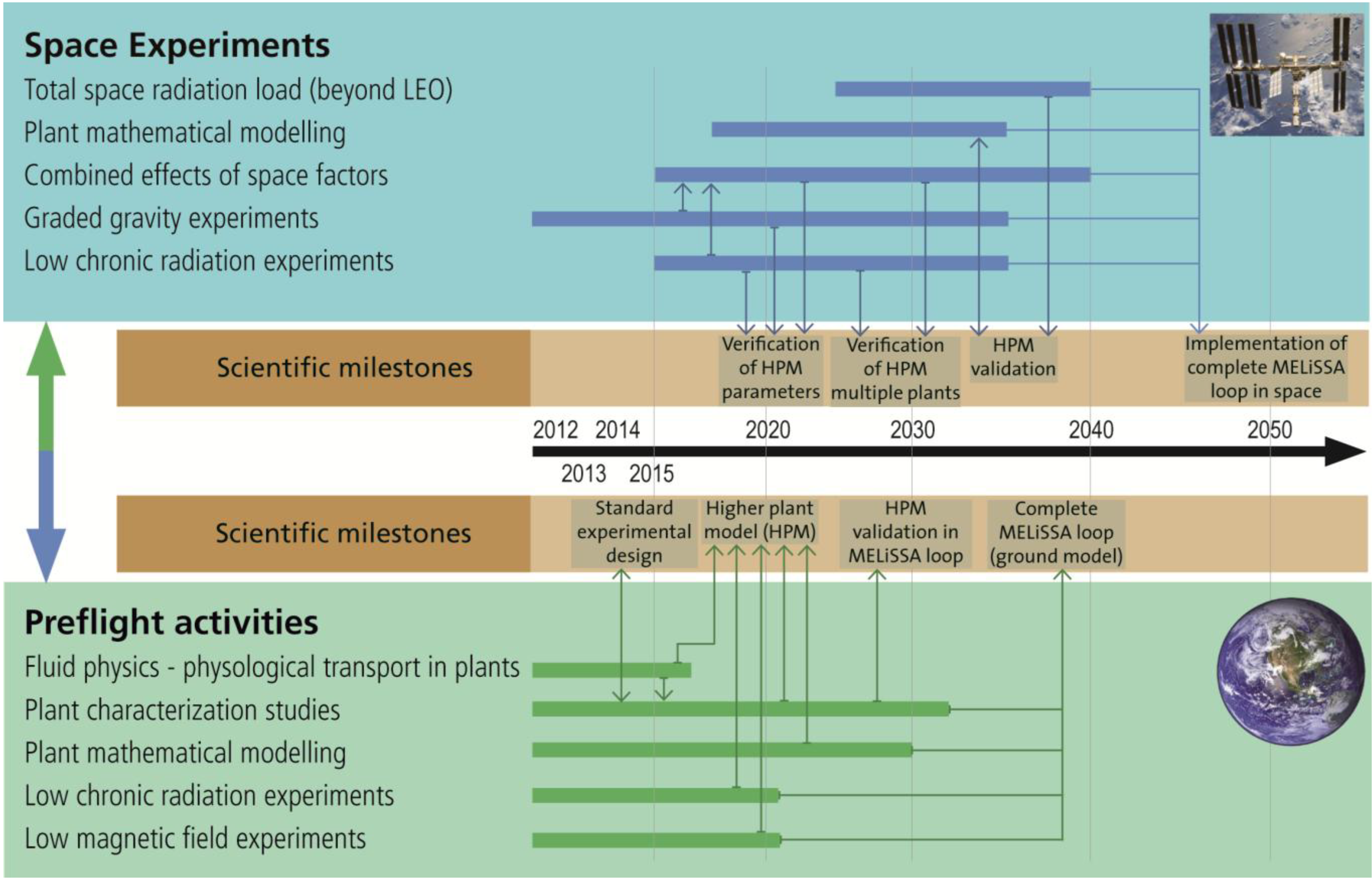
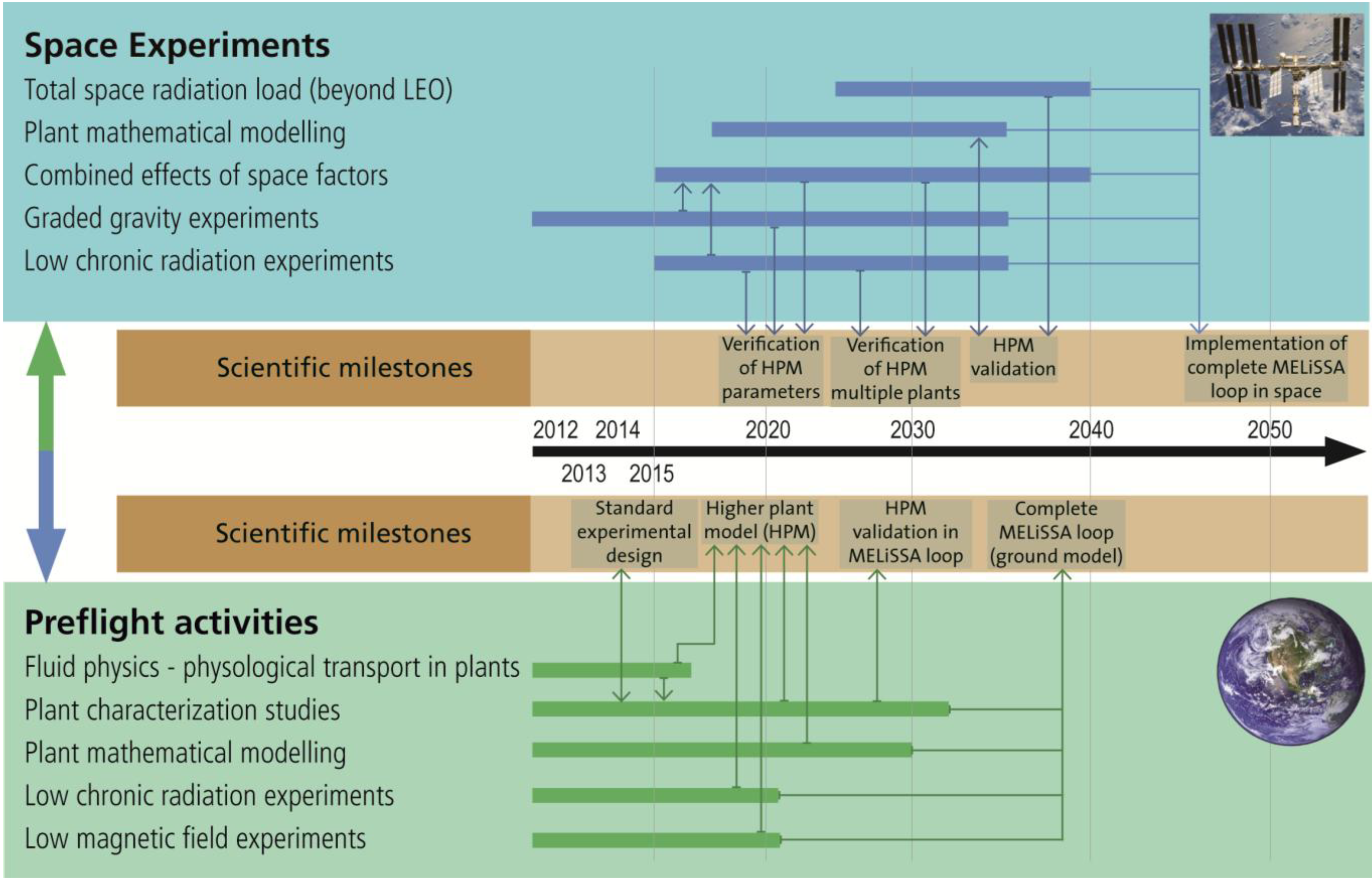
3.4. Roadmap
4. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| BDTC | Buoyancy Driven Thermal Convection |
| CRLSS | Closed Regenerative Life Support System |
| EMCS | European Modular Cultivation System |
| GCR | Higher Plant Model |
| HPM | Galactic Cosmic Rays |
| ISS | International Space Station |
| LEO | Low Earth Orbit |
| LiRHiPliSME | Literature Review of Higher Plants in Space for MELiSSA |
| MELiSSA | Micro-Ecological Life Support System Alternative |
| RPM | Random Positioning Machine |
| SAA | South Atlantic Anomaly |
| SPE | Solar Particle Event |
Conflicts of Interest
References
- Godia, F.; Albiol, J.; Montesinos, J.L.; Perez, J.; Creus, N.; Cabello, F.; Mengual, X.; Montras, A.; Lasseur, C. MELiSSA: A loop of interconnected bioreactors to develop life support in space. J. Biotechnol. 2002, 99, 319–330. [Google Scholar] [CrossRef]
- Godia, F.; Albiol, J.; Perez, J.; Creus, N.; Cabello, F.; Montras, A.; Masot, A.; Lasseur, C. The MELiSSA pilot plant facility as an integration test-bed for advanced life support systems. Adv. Space Res. 2004, 34, 1483–1493. [Google Scholar] [CrossRef]
- Hendrickx, L.; de Wever, H.; Hermans, V.; Mastroleo, F.; Morin, N.; Wilmotte, A.; Janssen, P.; Mergeay, M. Microbial ecology of the closed artificial ecosystem MELiSSA (micro-ecological life support system alternative): Reinventing and compartmentalizing the earth’s food and oxygen regeneration system for long-haul space exploration missions. Res. Microbiol. 2006, 157, 77–86. [Google Scholar]
- Paradiso, R.; de Micco, V.; Buonomo, R.; Aronne, G.; Barbieri, G.; de Pascale, S. Soilless cultivation of soybean for bioregenerative life-support systems: A literature review and the experience of the MELiSSA project—Food characterisation phase I. Plant Biol. 2014, 16, 69–78. [Google Scholar] [CrossRef]
- Kiss, J.Z. Plant biology in reduced gravity on the Moon and Mars. Plant Biol. 2014, 16, 12–17. [Google Scholar] [CrossRef]
- Wheeler, R.M.; Stutte, G.W.; Sobarrao, G.V.; Yorio, N.C. Plant growth and human life support for space travel. In Handbook of Plant and Crop Physiology; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA; Basel, Switzerland, 2001; pp. 925–941. [Google Scholar]
- Ferl, R.; Wheeler, R.; Levine, H.G.; Paul, A.L. Plants in space. Curr. Opin. Plant Biol. 2002, 5, 258–263. [Google Scholar] [CrossRef]
- Ivanova, T.N.; Bercovich, Y.A.; Mashinskiy, A.L.; Meleshko, G.I. The 1st space vegetables have been grown in the svet greenhouse using controlled environmental-conditions. Acta Astronaut. 1993, 29, 639–644. [Google Scholar] [CrossRef]
- Link, B.M.; Durst, S.J.; Zhou, W.; Stankovic, B. Seed-to-seed growth of arabidopsis thaliana on the international space station. Adv. Space Res. 2003, 31, 2237–2243. [Google Scholar] [CrossRef]
- Musgrave, M.E.; Kuang, A.X.; Matthews, S.W. Plant reproduction during spaceflight: Importance of the gaseous environment. Planta 1997, 203, S177–S184. [Google Scholar] [CrossRef]
- Sychev, V.N.; Shepelev, E.Y.; Meleshko, G.I.; Gurieva, T.S.; Levinskikh, M.A.; Podolsky, I.G.; Dadasheva, O.A.; Popov, V.V. Main characteristics of biological components of developing life support system observed during the experiments aboard orbital complex mir. Adv. Space Res. 2001, 27, 1529–1534. [Google Scholar] [CrossRef]
- Sychev, V.N.; Levinskikh, M.A.; Podolsky, I.G. Biological component of life support systems for a crew in long-duration space expeditions. Acta Astronaut. 2008, 63, 1119–1125. [Google Scholar] [CrossRef]
- Stanković, B. A plant space odyssey. Trends Plant Sci. 2001, 6, 591–593. [Google Scholar] [CrossRef]
- Ward, C.H.; Wilks, S.S.; Craft, H.L. Effects of prolonged near weightlessness on growth and gas exchange of photosynthetic plants. Dev. Ind. Microbiol. 1970, 11, 276–295. [Google Scholar]
- Johnson, S.P.; Tibbitts, T.W. Liminal angle of a plagiogeotropic organ under weightlessness. Bioscience 1968, 18, 655–661. [Google Scholar] [CrossRef]
- Merkys, A.J.; Laurinavicius, R.S.; Svegzdiene, D.V. Plant growth, development and embryogenesis during salyut-7 flight. Adv. Space Res. 1984, 4, 55–63. [Google Scholar]
- Wolff, S.A.; Coelho, L.H.; Zabrodina, M.; Brinckmann, E.; Kittang, A.I. Plant mineral nutrition, gas exchange and photosynthesis in space: A review. Adv. Space Res. 2013, 51, 465–475. [Google Scholar] [CrossRef]
- Karoliussen, I.B.E.; Kittang, A.-I. Will plants grow on Moon or Mars? Curr. Biotechnol. 2013, 2, 235–243. [Google Scholar] [CrossRef]
- Acuña, M.H. The magnetic field of mars. Lead. Edge 2003, 22, 769–771. [Google Scholar] [CrossRef]
- Acuña, M.H.; Connerney, J.E.P.; Wasilewski, P.; Lin, R.P.; Anderson, K.A.; Carlson, C.W.; McFadden, J.; Curtis, D.W.; Mitchell, D.; Reme, H.; et al. Magnetic field and plasma observations at mars: Initial results of the Mars global surveyor mission. Science 1998, 279, 1676–1680. [Google Scholar] [CrossRef]
- Mitchell, D.L.; Halekas, J.S.; Lin, R.P.; Frey, S.; Hood, L.L.; Acuna, M.H.; Binder, A. Global mapping of lunar crustal magnetic fields by lunar prospector. Icarus 2008, 194, 401–409. [Google Scholar] [CrossRef]
- Purucker, M.E. A global model of the internal magnetic field of the Moon based on lunar prospector magnetometer observations. Icarus 2008, 197, 19–23. [Google Scholar] [CrossRef]
- Rapp, D. Radiation effects and shielding requirements in human missions to the Moon and Mars. MARS Int. J. Mars Sci. Exp. 2006, 2, 46–71. [Google Scholar]
- Yang, C.H.; Craise, L.M.; Durante, M.; Mei, M. Heavy-ion-induced genetic changes and evolution processes. Life Sci. Space Res. 1994, 14, 373–382. [Google Scholar]
- Berger, T. Radiation dosimetry onboard the international space station iss. Z Med. Phys. 2008, 18, 265–275. [Google Scholar] [CrossRef]
- Musgrave, M.E. Growing plants in space. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2. No. 065. [Google Scholar]
- Sychev, V.N.; Levinskikh, M.A.; Gostimsky, S.A.; Bingham, G.E.; Podolsky, I.G. Spaceflight effects on consecutive generations of peas grown onboard the russian segment of the international space station. Acta Astronaut. 2007, 60, 426–432. [Google Scholar] [CrossRef]
- Wolverton, C.; Kiss, J.Z. An update on plant space biology. Gravit. Space Biol. 2011, 22. No. 2. [Google Scholar]
- De Micco, V.; de Pascale, S.; Paradiso, R.; Aronne, G. Microgravity effects on different stages of higher plant life cycle and completion of the seed-to-seed cycle. Plant Biol. 2014, 16, 31–38. [Google Scholar] [CrossRef]
- Kordyum, E.L. Plant cell gravisensitivity and adaptation to microgravity. Plant Biol. 2014, 16, 79–90. [Google Scholar] [CrossRef]
- Porterfield, D.M. The biophysical limitations in physiological transport and exchange in plants grown in microgravity. J. Plant Growth Regul. 2002, 21, 177–190. [Google Scholar] [CrossRef]
- Kitaya, Y.; Kawai, M.; Tsuruyama, J.; Takahashi, H.; Tani, A.; Goto, E.; Saito, T.; Kiyota, M. The effect of gravity on surface temperatures of plant leaves. Plant Cell Environ. 2003, 26, 497–503. [Google Scholar] [CrossRef]
- Liao, J.; Liu, G.; Monje, O.; Stutte, G.W.; Porterfield, D.M. Induction of hypoxic root metabolism results from physical limitations in O2 bioavailability in microgravity. Adv. Space Res. 2004, 34, 1579–1584. [Google Scholar] [CrossRef]
- Briarty, L.G.; Maher, E.P. Reserve utilization in seeds of arabidopsis thaliana germinating in microgravity. Int. J. Plant Sci. 2004, 165, 545–551. [Google Scholar] [CrossRef]
- Porterfield, D.M.; Barta, D.J.; Ming, D.W.; Morrow, R.C.; Musgrave, M.E. Astroculture (tm) root metabolism and cytochemical analysis. Adv. Space Res. 2000, 26, 315–318. [Google Scholar] [CrossRef]
- Nechitailo, G.; Gordeev, A. Effect of artificial electric fields on plants grown under microgravity conditions. Adv. Space Res. 2001, 28, 629–631. [Google Scholar] [CrossRef]
- Aliyev, A.A.; Abilov, Z.K.; Mashinskiy, A.L.; Ganiyeva, R.A.; Ragimova, G.K. The ultrastructure and physiological characteristics of the photosynthesis system of shoots of garden pea grown for 29 days on the “salyut-7”space station. USSR Space Life Sci. Dig. 1987, 10, 6. [Google Scholar]
- Belyavskaya, N.A. Free and membrane-bound calcium in microgravity and microgravity effects at the membrane level. Adv. Space Res. 1995, 17, 169–177. [Google Scholar] [CrossRef]
- Kordyum, E.L.; Belyavskaya, N.A.; Nedukha, E.M.; Palladina, T.A.; Tarasenko, V.A. The role of calcium ions in cytological effects of hypogravity. Adv. Space Res. 1984, 4, 23–26. [Google Scholar]
- Levine, H.G.; Krikorian, A.D. Changes in plant medium composition after a spaceflight experiment: Potassium levels are of special interest. Adv. Space Res. 2008, 42, 1060–1065. [Google Scholar] [CrossRef]
- Heyenga, A.G.; Forsman, A.; Stodieck, L.S.; Hoehn, A.; Kliss, M. Approaches in the determination of plant nutrient uptake and distribution in space flight conditions. Adv. Space Res. 2000, 26, 299–302. [Google Scholar] [CrossRef]
- Stutte, G.W.; Monje, O.; Goins, G.D.; Tripathy, B.C. Microgravity effects on thylakoid, single leaf, and whole canopy photosynthesis of dwarf wheat. Planta 2005, 223, 46–56. [Google Scholar]
- Tripathy, B.C.; Brown, C.S.; Levine, H.G.; Krikorian, A.D. Growth and photosynthetic responses of wheat plants crown in space. Plant Physiol. 1996, 110, 801–806. [Google Scholar]
- Kiss, J.Z.; Guisinger, M.M.; Miller, A.J.; Stackhouse, K.S. Reduced gravitropism in hypocotyls of starch-deficient mutants of arabidopsis. Plant Cell Physiol. 1997, 38, 518–525. [Google Scholar] [CrossRef]
- Kiss, J.Z. Mechanisms of the early phases of plant gravitropism. Crit. Rev. Plant Sci. 2000, 19, 551–573. [Google Scholar] [CrossRef]
- Wyatt, S.E.; Kiss, J.Z. Plant tropisms: From darwin to the international space station. Am. J. Bot. 2013, 100, 1–3. [Google Scholar] [CrossRef]
- Blancaflor, E.B.; Masson, P.H. Plant gravitropism. Unraveling the ups and downs of a complex process. Plant Physiol. 2003, 133, 1677–1690. [Google Scholar] [CrossRef]
- Perbal, G.; Driss-Ecole, D. Contributions of space experiments to the study of gravitropism. J. Plant Growth Regul. 2002, 21, 156–165. [Google Scholar] [CrossRef]
- Morita, M.T. Directional gravity sensing in gravitropism. Annu. Rev. Plant. Biol. 2010, 61, 705–720. [Google Scholar] [CrossRef]
- Toyota, M.; Gilroy, S. Gravitropism and mechanical signaling in plants. Am. J. Bot. 2013, 100, 111–125. [Google Scholar] [CrossRef]
- Driss-Ecole, D.; Legue, V.; Carnero-Diaz, E.; Perbal, G. Gravisensitivity and automorphogenesis of lentil seedling roots grown on board the international space station. Physiol. Plant. 2008, 134, 191–201. [Google Scholar] [CrossRef]
- Cassab, G.I. Other tropisms and their relationship to gravitropism. In Plant Tropisms; Wiley: New York, NY, USA, 2008; pp. 123–139. [Google Scholar]
- Millar, K.D.L.; Kumar, P.; Correll, M.J.; Mullen, J.L.; Hangarter, R.P.; Edelmann, R.E.; Kiss, J.Z. A novel phototropic response to red light is revealed in microgravity. New Phytol. 2010, 186, 648–656. [Google Scholar] [CrossRef]
- Takahashi, H.; Mizuno, H.; Kamada, M.; Fujii, N.; Higashitani, A.; Kamigaichi, S.; Aizawa, S.; Mukai, C.; Shimazu, T.; Fukui, K.; et al. A spaceflight experiment for the study of gravimorphogenesis and hydrotropism in cucumber seedlings. J. Plant Res. 1999, 112, 497–505. [Google Scholar] [CrossRef]
- Hoson, T.; Saiki, M.; Kamisaka, S.; Yamashita, M. Automorphogenesis and gravitropism of plant seedlings grown under microgravity conditions. Adv. Space Res. 2001, 27, 933–940. [Google Scholar] [CrossRef]
- Soga, K.; Wakabayashi, K.; Kamisaka, S.; Hoson, T. Stimulation of elongation growth and xyloglucan breakdown in arabidopsis hypocotyls under microgravity conditions in space. Planta 2002, 215, 1040–1046. [Google Scholar] [CrossRef]
- Nedukha, E. Effects of microgravity on the structure and function of plant cell walls. Int. Rev. Cytol. 1997, 170, 39–77. [Google Scholar] [CrossRef]
- Nevzgodina, L.V. Chromosomal aberrations as a biomarker for cosmic radiation. In Fundamentals for the Assessment of Risks from Environmental Radiation; NATO Science Series; Springer Netherlands: Houten, The Netherlands, 1999; pp. 203–208. [Google Scholar]
- Kovalchuk, O.; Arkhipov, A.; Barylyak, I.; Karachov, I.; Titov, V.; Hohn, B.; Kovalchuk, I. Plants experiencing chronic internal exposure to ionizing radiation exhibit higher frequency of homologous recombination than acutely irradiated plants. Mutat. Res. 2000, 449, 47–56. [Google Scholar] [CrossRef]
- Real, A.; Sundell-Bergman, S.; Knowles, J.F.; Woodhead, D.S.; Zinger, I. Effects of ionising radiation exposure on plants, fish and mammals: Relevant data for environmental radiation protection. J. Radiol. Prot. 2004, 24, A123–A137. [Google Scholar] [CrossRef]
- Esnault, M.A.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Kovalchuk, I.; Molinier, J.; Yao, Y.L.; Arkhipov, A.; Kovalchuk, O. Transcriptome analysis reveals fundamental differences in plant response to acute and chronic exposure to ionizing radiation. Mutat. Res. 2007, 624, 101–113. [Google Scholar] [CrossRef]
- Vandenhove, H.; Vanhoudt, N.; Cuypers, A.; van Hees, M.; Wannijn, J.; Horemans, N. Life-cycle chronic gamma exposure of arabidopsis thaliana induces growth effects but no discernable effects on oxidative stress pathways. Plant Physiol. Biochem. 2010, 48, 778–786. [Google Scholar] [CrossRef]
- Bhaskaran, S.; Swaminathan, M.S. Chromosome aberrations, changes in DNA content and frequency and spectrum of mutations induced by X-rays and neutrons in polyploids. Radiat. Bot. 1961, 1, 166–174. [Google Scholar] [CrossRef]
- Galland, P.; Pazur, A. Magnetoreception in plants. J. Plant Res. 2005, 118, 371–389. [Google Scholar] [CrossRef]
- Belyavskaya, N.A. Biological effects due to weak magnetic field on plants. Adv. Space Res. 2004, 34, 1566–1574. [Google Scholar] [CrossRef]
- Travkin, M.P. Change of bioelectric activity of setereasea purpurea under effect of constant and pulsating magnetic-field. Biofizika 1973, 18, 172–174. [Google Scholar]
- Ahmad, M.; Galland, P.; Ritz, T.; Wiltschko, R.; Wiltschko, W. Magnetic intensity affects cryptochrome-dependent responses in arabidopsis thaliana. Planta 2007, 225, 615–624. [Google Scholar] [CrossRef]
- Aladjadjiyan, A.; Zahariev, A.L. Influence of stationary magnetic field on the absorption spectra of some energy plants. J. Environ. Prot. Ecol. 2009, 10, 1032–1036. [Google Scholar]
- Brettel, K.; Setif, P. Magnetic-field effects on primary reactions in Photosystem-I. Biochim. Biophys. Acta 1987, 893, 109–114. [Google Scholar] [CrossRef]
- Hakala-Yatkin, M.; Sarvikas, P.; Paturi, P.; Mantysaari, M.; Mattila, H.; Tyystjarvi, T.; Nedbal, L.; Tyystjarvi, E. Magnetic field protects plants against high light by slowing down production of singlet oxygen. Physiol. Plant. 2011, 142, 26–34. [Google Scholar] [CrossRef]
- Jovanic, B.R.; Jovanic, R. Effect of a permanent magnetic field on the optical and physiological properties of green plant leaves. Int. J. Environ. Stud. 2002, 59, 599–606. [Google Scholar] [CrossRef]
- Yano, A.; Ohashi, Y.; Hirasaki, T.; Fuliwara, K. Effects of a 60 Hz magnetic field on photosynthetic CO2 uptake and early growth of radish seedlings. Bioelectromagnetics 2004, 25, 572–581. [Google Scholar] [CrossRef]
- Solov’yov, I.A.; Schulten, K. Reaction kinetics and mechanism of magnetic field effects in cryptochrome. J. Phys. Chem. B 2012, 116, 1089–1099. [Google Scholar] [CrossRef]
- Xu, C.X.; Yin, X.; Lv, Y.; Wu, C.Z.; Zhang, Y.X.; Song, T. A near-null magnetic field affects cryptochrome-related hypocotyl growth and flowering in arabidopsis. Adv. Space Res. 2012, 49, 834–840. [Google Scholar] [CrossRef]
- Occhipinti, A.; de Santis, A.; Maffei, M.E. Magnetoreception: An unavoidable step for plant evolution? Trends Plant Sci. 2014, 19, 1–4. [Google Scholar] [CrossRef]
- Yamashita, M.; Tomita-Yokotani, K.; Hashimoto, H.; Takai, M.; Tsushima, M.; Nakamura, T. Experimental concept for examination of biological effects of magnetic field concealed by gravity. Adv. Space Res. 2004, 34, 1575–1578. [Google Scholar] [CrossRef]
- Ruyters, G.; Braun, M. Plant biology in space: Recent accomplishments and recommendations for future research. Plant Biol. 2014, 16, 4–11. [Google Scholar] [CrossRef]
- Gielis, J.; Gerats, T. A botanical perspective on modeling plants and plant shapes in computer graphics. In proceedings of International Conference on Computing, Communications and Control Technologies, Austin, TX, USA, 14–17 August 2004; pp. 265–272.
- Hezard, P.; Sasidharan, L.S.; Creuly, C.; Dussap, C.-G. Higher plant modeling for bioregenerative life support applications: General structure of modeling. In Proceedings of the 40th International Conference on Environmental Systems, Barcelona, Spain, 11–15 July 2010.
- Holmberg, M.; Paille, C.; Lasseur, C. Preliminary modelling of mass flux at the surface of plant leaves within the MELiSSA higher plant compartments. In Proceedings of the 38th COSPAR Scientific Assembly, Bremen, Germany, 15–18 July 2010.
- Stasiak, M.; Gidzinski, D.; Jordan, M.; Dixon, M. Crop selection for advanced life support systems in the ESA MELiSSA program: Durum wheat (triticum turgidum var durum). Adv. Space Res. 2012, 49, 1684–1690. [Google Scholar] [CrossRef]
- Molders, K.; Quinet, M.; Decat, J.; Secco, B.; Duliere, E.; Pieters, S.; van der Kooij, T.; Lutts, S.; van der Straeten, D. Selection and hydroponic growth of potato cultivars for bioregenerative life support systems. Adv. Space Res. 2012, 50, 156–165. [Google Scholar]
- Van Loon, J.J. Some history and use of the random positioning machine, rpm, in gravity related research. Adv. Space Res. 2007, 39, 1161–1165. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef]
- De Micco, V.; Buonomo, R.; Paradiso, R.; de Pascale, S.; Aronne, G. Soybean cultivar selection for bioregenerative life support systems (blss)—Theoretical selection. Adv. Space Res. 2012, 49, 1415–1421. [Google Scholar] [CrossRef]
- Hoson, T.; Soga, K. New aspects of gravity responses in plant cells. Int. Rev. Cytol. 2003, 229, 209–244. [Google Scholar] [CrossRef]
- Hoson, T.; Soga, K.; Mori, R.; Saiki, M.; Nakamura, Y.; Wakabayashi, K.; Kamisaka, S. Cell wall changes involved in the automorphic curvature of rice coleoptiles under microgravity conditions in space. J. Plant Res. 2004, 117, 449–455. [Google Scholar] [CrossRef]
- Hoson, T.; Soga, K.; Wakabayashi, K.; Kamisaka, S.; Tanimoto, E. Growth and cell wall changes in rice roots during spaceflight. Plant Soil. 2003, 255, 19–26. [Google Scholar] [CrossRef]
- Musgrave, M.E.; Kuang, A.; Tuominen, L.K.; Levine, L.H.; Morrow, R.C. Seed storage reserves and glucosinolates in brassica rapa l. Grown on the international space station. J. Am. Soc. Hortic. Sci. 2005, 130, 848–856. [Google Scholar]
- Allen, J.; Bisbee, P.A.; Darnell, R.L.; Kuang, A.; Levine, L.H.; Musgrave, M.E.; van Loon, J.J. Gravity control of growth form in brassica rapa and arabidopsis thaliana (brassicaceae): Consequences for secondary metabolism. Am. J. Bot. 2009, 96, 652–660. [Google Scholar] [CrossRef]
- Tuominen, L.K.; Levine, L.H.; Musgrave, M.E. Plant Secondary Metabolism in Altered Gravity; Humana Press: New York, NY, USA, 2009; pp. 373–386. [Google Scholar]
- Brinckmann, E. Centrifuges and their application for biological experiments in space. Microgr. Sci. Technol. 2012, 24, 365–372. [Google Scholar] [CrossRef]
- Brinckmann, E. Esa hardware for plant research on the international space station. Adv. Space Res. 2005, 36, 1162–1166. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Edelmann, R.E.; Wood, P.C. Gravitropism of hypocotyls of wild-type and starch-deficient arabidopsis seedlings in spaceflight studies. Planta 1999, 209, 96–103. [Google Scholar]
- Kittang, A.I.; Iversen, T.H.; Fossum, K.R.; Mazars, C.; Carnero-Diaz, E.; Boucheron-Dubuisson, E.; le Disquet, I.; Legué, V.; Herranz, R.; Pereda-Loth, V.; et al. Exploration of plant growth and development using the european modular cultivation system facility on the international space station. Plant Biol. 2014, 16, 528–538. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wolff, S.A.; Coelho, L.H.; Karoliussen, I.; Jost, A.-I.K. Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment. Life 2014, 4, 189-204. https://doi.org/10.3390/life4020189
Wolff SA, Coelho LH, Karoliussen I, Jost A-IK. Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment. Life. 2014; 4(2):189-204. https://doi.org/10.3390/life4020189
Chicago/Turabian StyleWolff, Silje A., Liz H. Coelho, Irene Karoliussen, and Ann-Iren Kittang Jost. 2014. "Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment" Life 4, no. 2: 189-204. https://doi.org/10.3390/life4020189
APA StyleWolff, S. A., Coelho, L. H., Karoliussen, I., & Jost, A.-I. K. (2014). Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment. Life, 4(2), 189-204. https://doi.org/10.3390/life4020189