
Physiological and Transcriptome Analysis Revealed the Effect of ABA on Promoting Persimmon Fruit Postharvest Deastringency



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Soluble and Resoluble Tannin Contents
2.3. Ethanol and Acetaldehyde Measurements
2.4. RNA Isolation and Transcriptomic Analysis
2.5. Weighted Gene Co-Expression Network Analysis (WGCNA)
2.6. cDNA Synthesis and RT-qPCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of ABA Treatment on Persimmon Fruit Postharvest De-Astringency
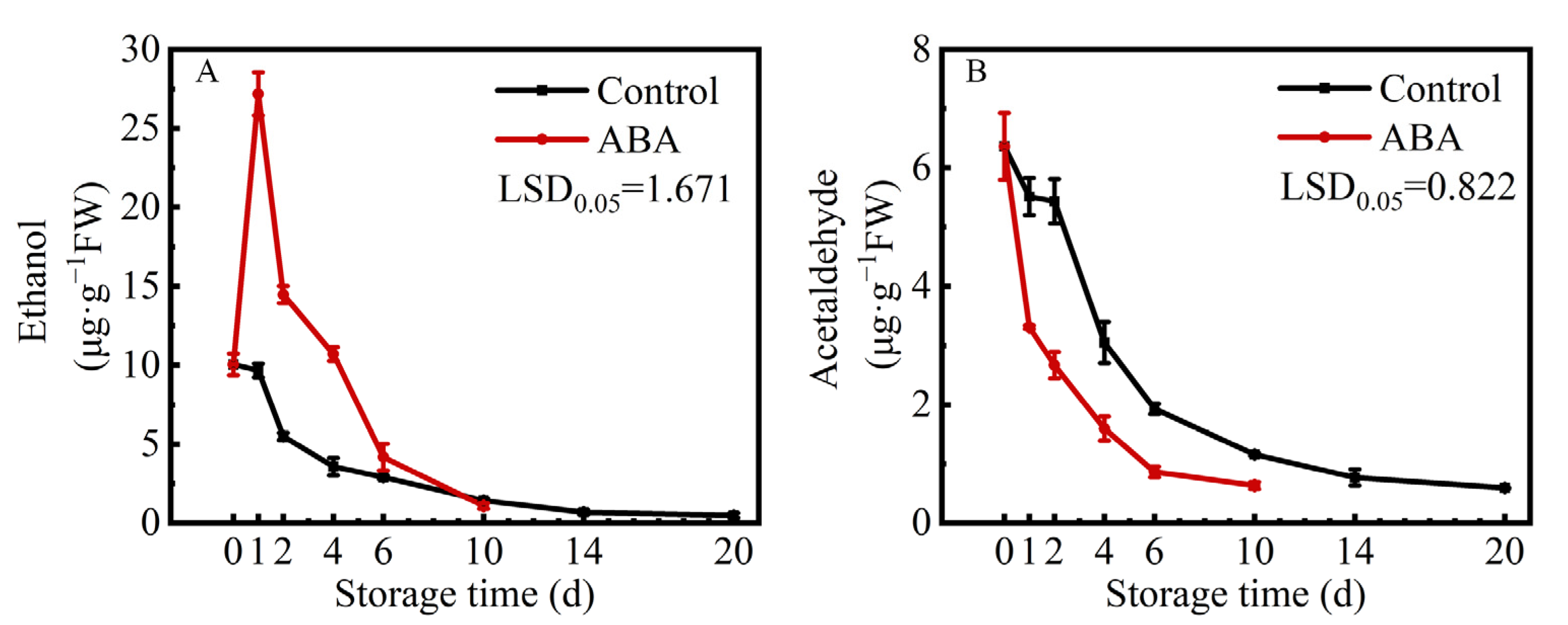
3.2. Effects of ABA Treatment on Endogenous Ethanol and Acetaldehyde Contents
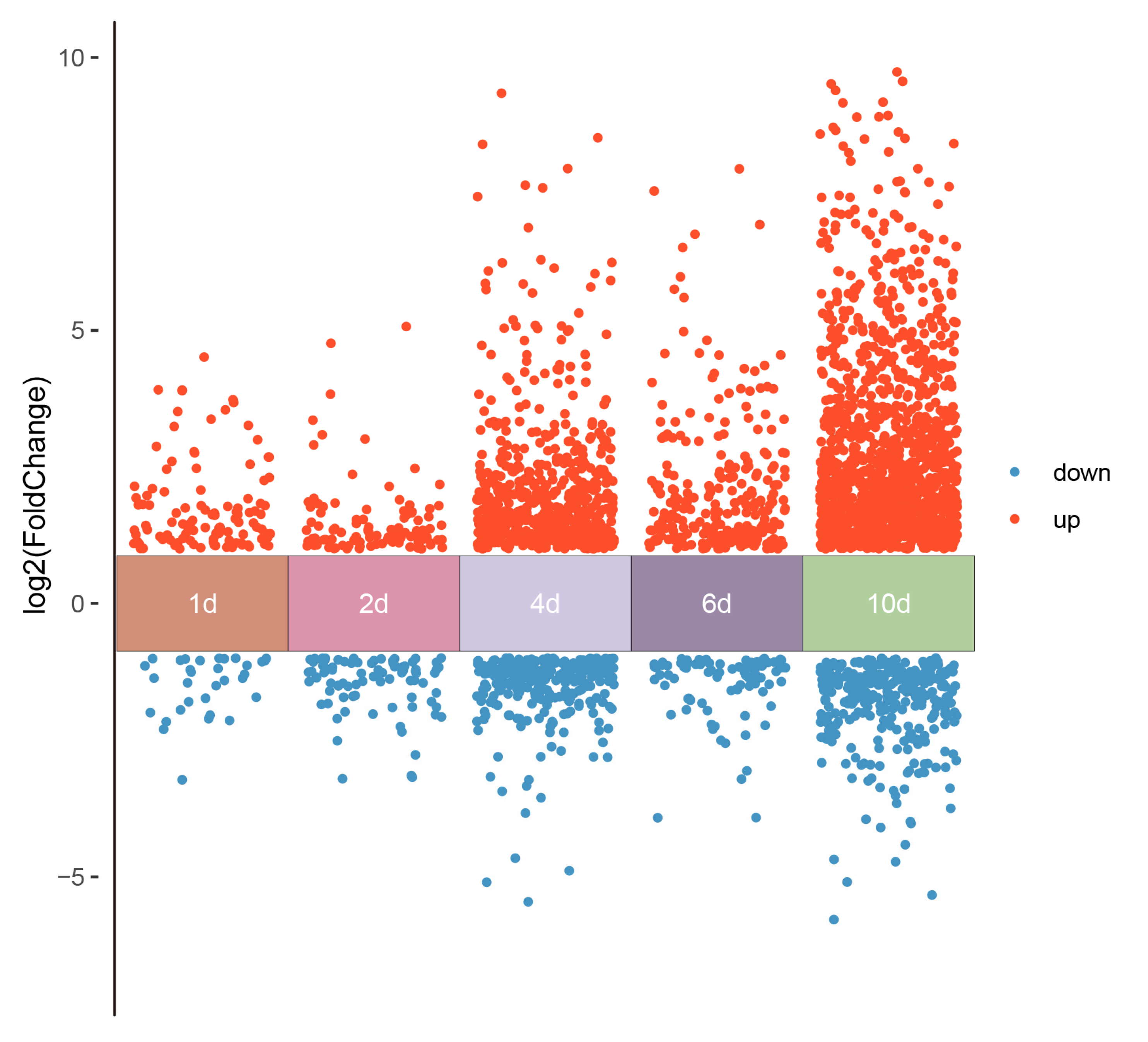
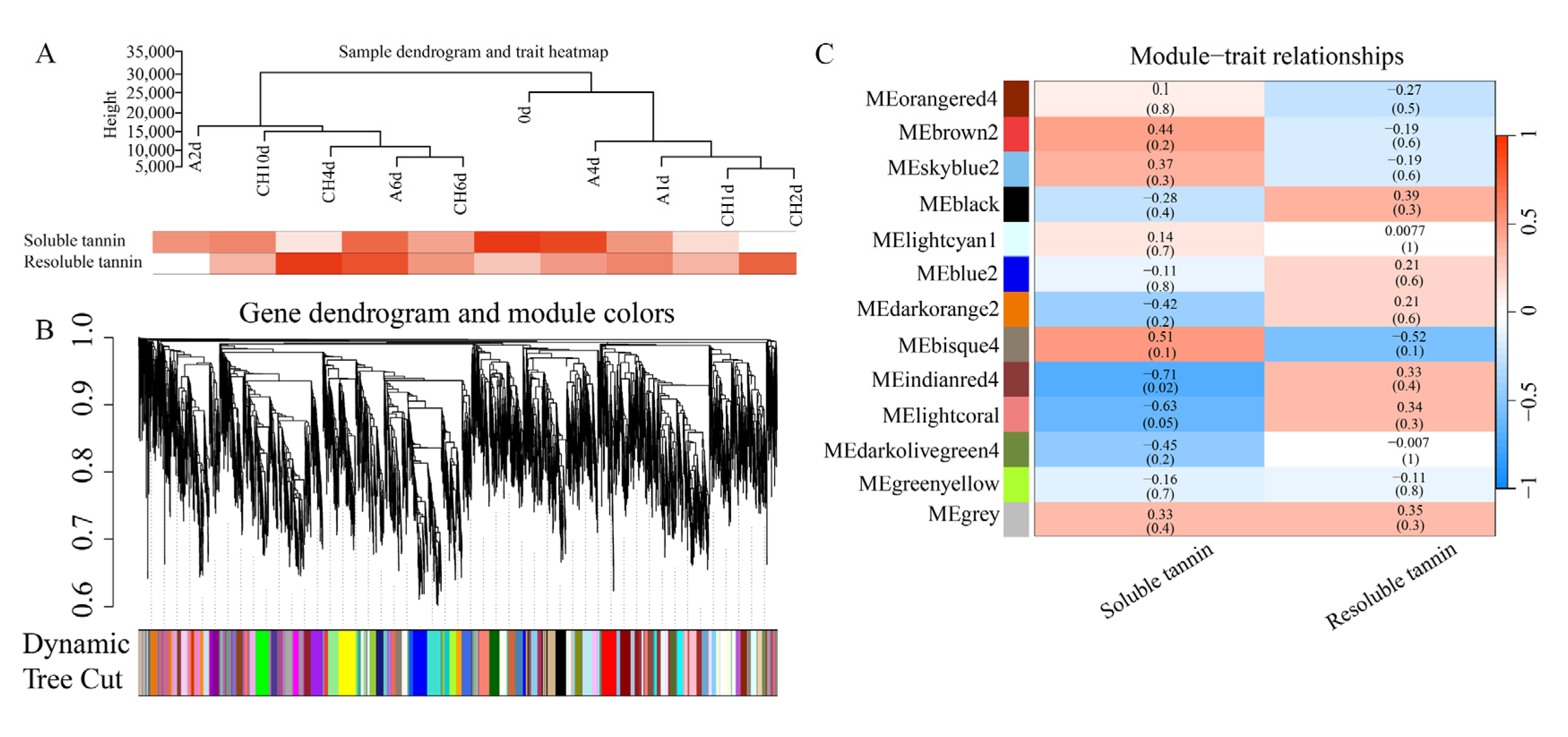
3.3. RNA-Seq and Weighted Gene Co-Expression Network Analysis
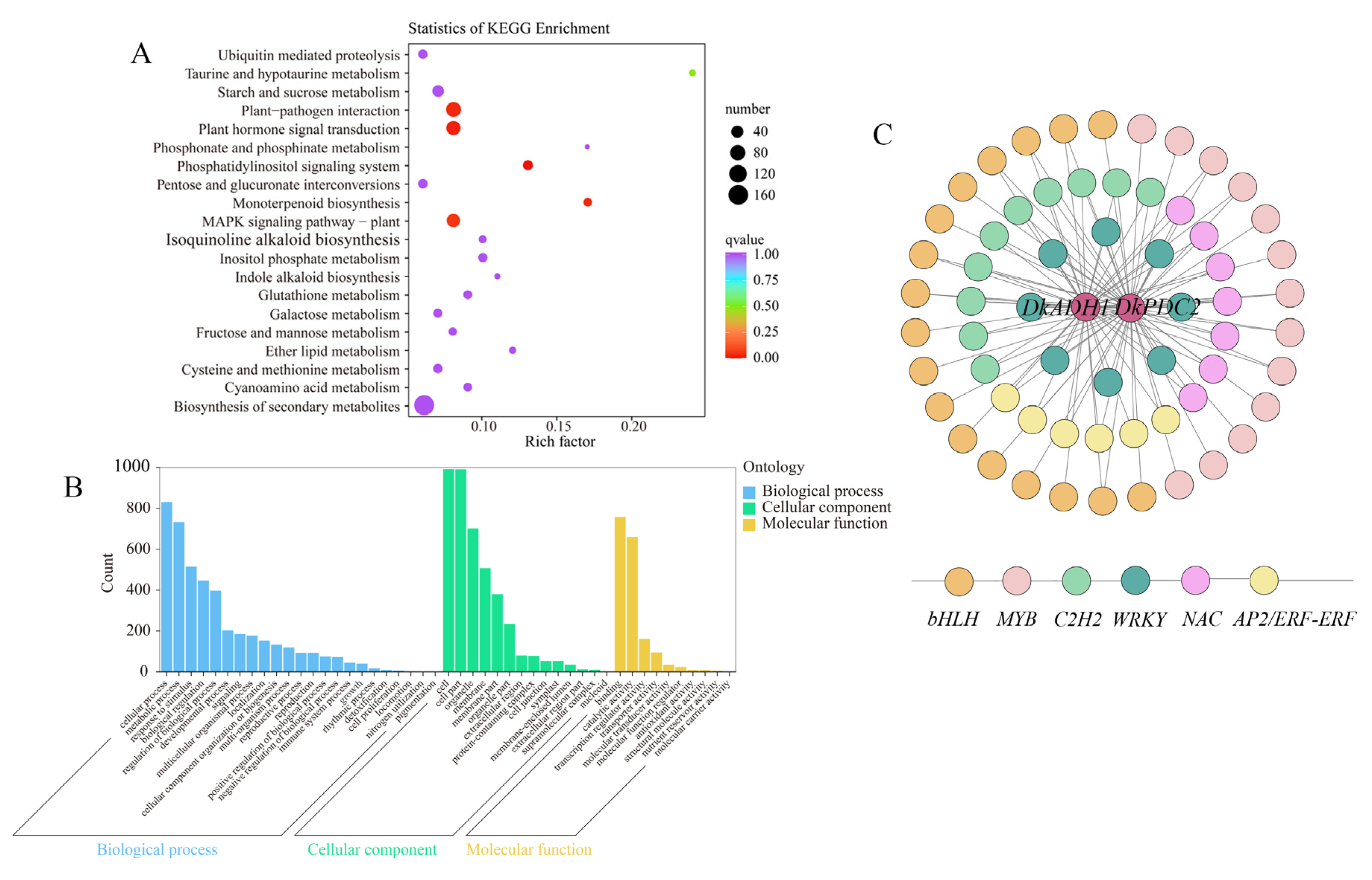
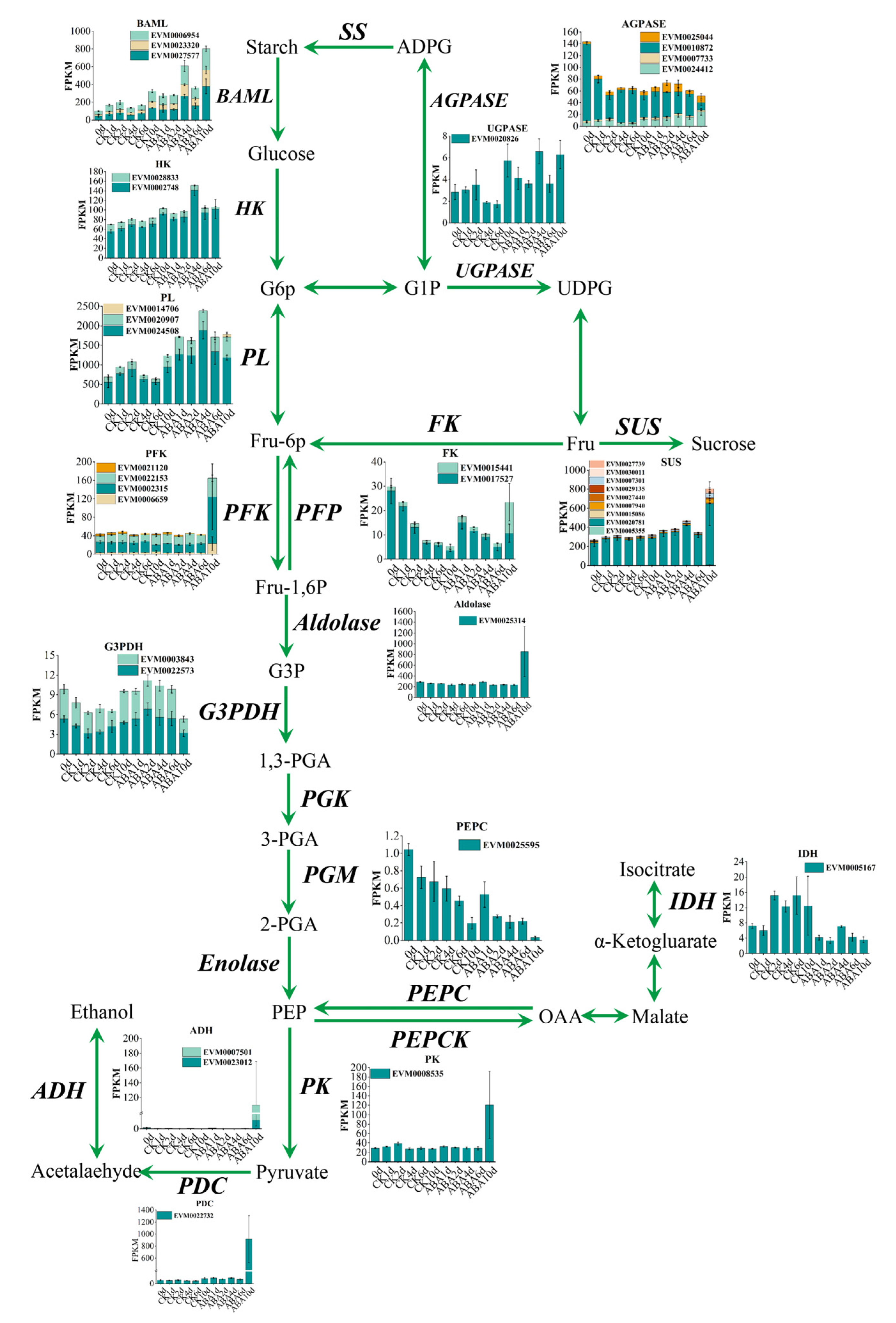
3.4. Differential Responses of Carbohydrate Metabolism Related Genes to ABA Treatment in Persimmon Fruit
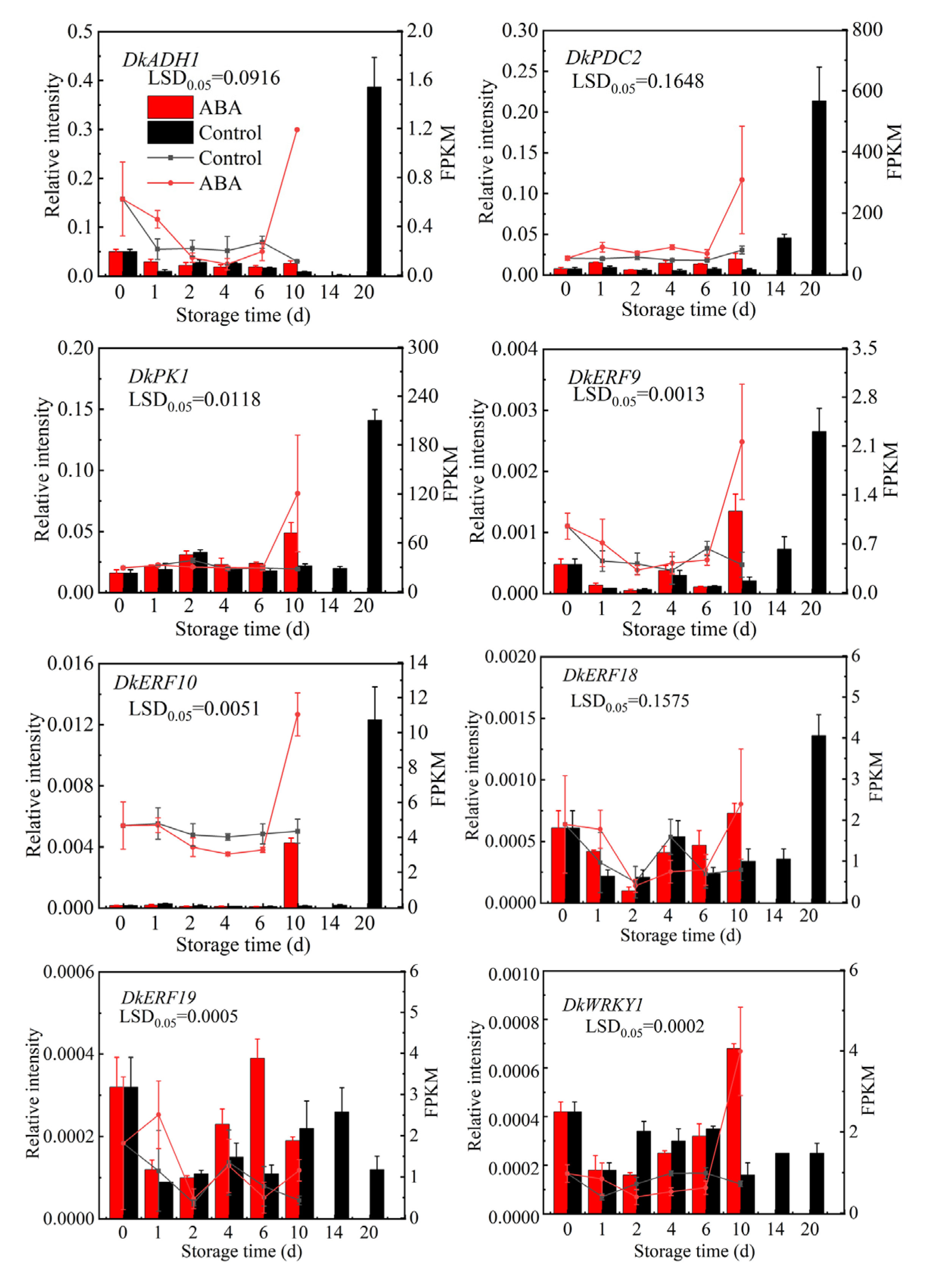
3.5. RT-qPCR Validation of RNA-Seq Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Q.-G.; Xu, Y.; Yang, Y.; Guan, C.-F.; Zhang, Q.-Y.; Huang, J.-W.; Grierson, D.; Chen, K.-S.; Gong, B.-C.; Yin, X.-R. The persimmon (Diospyros oleifera Cheng) genome provides new insights into the inheritance of astringency and ancestral evolution. Hortic. Res. 2019, 6, 138. [Google Scholar] [CrossRef]
- Min, T.; Yin, X.-R.; Shi, Y.-N.; Luo, Z.-R.; Yao, Y.-C.; Grierson, D.; Ferguson, I.B.; Chen, K.-S. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon (Diospyros kaki) fruit de-astringency. J. Exp. Bot. 2012, 63, 6393–6405. [Google Scholar] [CrossRef]
- Yin, X.-R.; Shi, Y.-N.; Min, T.; Luo, Z.-R.; Yao, Y.-C.; Xu, Q.; Ferguson, I.; Chen, K.-S. Expression of ethylene response genes during persimmon fruit astringency removal. Planta 2012, 235, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Chen, L.; Chen, W.; Mo, R.; Zhang, Q.; Du, X.; Liu, J.; Luo, Z. SSAP analysis reveals candidate genes associated with deastringency in persimmon (Diospyros kaki Thunb.) treated with 40 °C water. Tree Genet. Genomes 2015, 11, 20. [Google Scholar] [CrossRef]
- Salvador, A.; Arnal, L.; Besada, C.; Larrea, V.; Quiles, A.; Pérez-Munuera, I. Physiological and structural changes during ripening and deastringency treatment of persimmon fruit cv. ‘Rojo Brillante’. Postharvest Biol. Technol. 2007, 46, 181–188. [Google Scholar] [CrossRef]
- Wu, W.; Zhu, Q.-G.; Wang, W.-Q.; Grierson, D.; Yin, X.-R. Molecular basis of the formation and removal of fruit astringency. Food Chem. 2022, 372, 131234. [Google Scholar] [CrossRef]
- Kou, S.-M.; Jin, R.; Wu, Y.-Y.; Huang, J.-W.; Zhang, Q.-Y.; Sun, N.-J.; Yang, Y.; Guan, C.-F.; Wang, W.-Q.; Zhu, C.-Q.; et al. Transcriptome analysis revealed the roles of carbohydrate metabolism on differential acetaldehyde production capacity in persimmon fruit in response to high-CO2 treatment. J. Agric. Food Chem. 2021, 69, 836–845. [Google Scholar] [CrossRef]
- Li, S.; Chen, K.; Grierson, D. Molecular and hormonal mechanisms regulating fleshy fruit ripening. Cells 2021, 10, 1136. [Google Scholar] [CrossRef]
- Wu, W.; Wang, W.-Q.; Zhu, Q.-G.; Jin, R.; Liu, X.-F.; Grierson, D.; Yin, X.-R. Convergent and divergent regulations of ethylene and abscisic acid biosynthesis during persimmon fruit postharvest ripening. Postharvest Biol. Technol. 2022, 191, 111977. [Google Scholar] [CrossRef]
- Zhang, M.; Yuan, B.; Leng, P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef]
- Soto, A.; Ruiz, K.B.; Ravaglia, D.; Costa, G.; Torrigiani, P. ABA may promote or delay peach fruit ripening through modulation of ripening- and hormone-related gene expression depending on the developmental stage. Plant Physiol. Biochem. 2013, 64, 11–24. [Google Scholar] [CrossRef]
- Wu, Q.; Tao, X.; Ai, X.; Luo, Z.; Mao, L.; Ying, T.; Li, L. Contribution of abscisic acid to aromatic volatiles in cherry tomato (Solanum lycopersicum L.) fruit during postharvest ripening. Plant Physiol. Biochem. 2018, 130, 205–214. [Google Scholar] [CrossRef]
- Zou, J.; Li, N.; Hu, N.; Tang, N.; Cao, H.; Liu, Y.; Chen, J.; Jian, W.; Gao, Y.; Yang, J.; et al. Co-silencing of ABA receptors (SlRCAR) reveals interactions between ABA and ethylene signaling during tomato fruit ripening. Hortic. Res. 2022, 9, uhac057. [Google Scholar] [CrossRef]
- Park, D.S.; Tilahun, S.; Park, K.C.; Choi, I.Y.; Jeong, C.S. Transcriptome analysis of astringent ‘Cheongdo-Bansi’ persimmon fruit treated with ethylene for removal of astringency. Postharvest Biol. Technol. 2019, 150, 52–59. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Su, W.-Y.; Wu, W.; Zhu, Q.-G.; Yin, X.-R.; Sun, N.-J.; Liu, X.-F. Transcriptional responses of postharvest ABA treatment in persimmon fruit. Postharvest Biol. Technol. 2024, 217, 113097. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Arnal, L.; Del Río, M.A. Removing Astringency by carbon dioxide and nitrogen-enriched atmospheres in persimmon fruit cv. ‘Rojo brillante’. J. Food Sci. 2003, 68, 1516–1518. [Google Scholar] [CrossRef]
- Zhu, Q.-G.; Gong, Z.-Y.; Wang, M.-M.; Li, X.; Grierson, D.; Yin, X.-R.; Chen, K.-S. A transcription factor network responsive to high CO2/hypoxia is involved in deastringency in persimmon fruit. J. Exp. Bot. 2018, 69, 2061–2070. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.-G.; Gong, Z.-Y.; Huang, J.; Grierson, D.; Chen, K.-S.; Yin, X.-R. High-CO2/hypoxia-responsive transcription factors DkERF24 and DkWRKY1 interact and activate DkPDC2 promoter. Plant Physiol. 2019, 180, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shen, X.; Ding, Y.; Zhang, P.; Zhu, Q.; Wang, Y.; Zhang, Q.; Luo, Z.; Yang, Y.; Du, X.; et al. A comprehensive transcriptomic and metabolomic map reveals the molecular mechanism of persimmon fruit deastringency upon 40 °C warm water treatment. Postharvest Biol. Technol. 2025, 220, 113313. [Google Scholar] [CrossRef]
- Liang, B.; Zheng, Y.; Wang, J.; Zhang, W.; Fu, Y.; Kai, W.; Xu, Y.; Yuan, B.; Li, Q.; Leng, P. Overexpression of the persimmon abscisic acid β-glucosidase gene (DkBG1) alters fruit ripening in transgenic tomato. Plant J. 2020, 102, 1220–1233. [Google Scholar] [CrossRef]
- Zhu, Q.-G.; Wang, M.-M.; Gong, Z.-Y.; Fang, F.; Sun, N.-J.; Li, X.; Grierson, D.; Yin, X.-R.; Chen, K.-S. Involvement of DkTGA1 transcription factor in anaerobic response leading to persimmon fruit postharvest de-astringency. PLoS ONE 2016, 11, e015591. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Nie, J.-J.; Ren, M.-L.; Ding, Y.-D.; Xu, Y.-X.; Zhu, Q.-G. Physiological and Transcriptome Analysis Revealed the Effect of ABA on Promoting Persimmon Fruit Postharvest Deastringency. Life 2025, 15, 1027. https://doi.org/10.3390/life15071027
Zhou H, Nie J-J, Ren M-L, Ding Y-D, Xu Y-X, Zhu Q-G. Physiological and Transcriptome Analysis Revealed the Effect of ABA on Promoting Persimmon Fruit Postharvest Deastringency. Life. 2025; 15(7):1027. https://doi.org/10.3390/life15071027
Chicago/Turabian StyleZhou, Han, Jiao-Jiao Nie, Meng-Lin Ren, Yu-Duan Ding, Ya-Xiu Xu, and Qing-Gang Zhu. 2025. "Physiological and Transcriptome Analysis Revealed the Effect of ABA on Promoting Persimmon Fruit Postharvest Deastringency" Life 15, no. 7: 1027. https://doi.org/10.3390/life15071027
APA StyleZhou, H., Nie, J.-J., Ren, M.-L., Ding, Y.-D., Xu, Y.-X., & Zhu, Q.-G. (2025). Physiological and Transcriptome Analysis Revealed the Effect of ABA on Promoting Persimmon Fruit Postharvest Deastringency. Life, 15(7), 1027. https://doi.org/10.3390/life15071027