

Phenotypical and Molecular Characterization of Acinetobacter baumannii Isolated from Hospitalized Patients During the COVID-19 Pandemic in Brazil
 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Identification by Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry (MALDI-TOF MS) Proteomic Characterization
2.3. Identification by 16S rRNA Gene Sequencing Analysis
2.4. Typing by FTIR Spectroscopy Analysis
2.5. Evaluation of Antimicrobial Susceptibility Profile
2.6. Evaluation of Biofilm Formation and Its Tolerance to Disinfectants
3. Results
3.1. Identification and Typing
3.2. Antimicrobial Susceptibility Profile
3.3. Biofilm Formation and Its Tolerance to Disinfectants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1

{kind=link}
{kind=link}
{kind=link}
| Strain ID | NCBI a Access Number | Base Pair Length (nt) | Identification (%) |
|---|---|---|---|
| AC001/21 | OQ955834 | 728 | A. baumannii (99.59) |
| AC002/21 | OQ955835 | 842 | A. baumannii (99.88) |
| AC003/21 | OQ955836 | 860 | A. baumannii (99.88) |
| AC004/21 | OQ955837 | 704 | A. baumannii (97.28) |
| AC005/21 | OQ955838 | 851 | A. baumannii (99.88) |
| AC006/21 | OQ955839 | 974 | A. baumannii (99.90) |
| AC007/21 | OQ955840 | 858 | A. baumannii (99.88) |
| AC008/21 | OQ955841 | 1.030 | A. baumannii (99.90) |
| AC009/21 | OQ955842 | 984 | A. baumannii (99.90) |
| AC010/21 | OQ955843 | 940 | A. baumannii (99.90) |
| AC012/21 | OQ955844 | 874 | A. baumannii (99.88) |
| AC013/21 | OQ955845 | 980 | A. baumannii (99.90) |
| AC014/21 | OQ955846 | 943 | A. baumannii (99.89) |
| AC015/21 | OQ955847 | 922 | A. baumannii (99.89) |
| AC016/21 | OQ955848 | 861 | A. baumannii (99.88) |
| AC017/21 | OQ955849 | 986 | A. baumannii (99.90) |
| AC018/22 | OQ955850 | 1.036 | A. baumannii (99.90) |
| AC019/22 | OQ955851 | 945 | A. baumannii (99.89) |
| AC020/22 | OQ955852 | 984 | A. baumannii (99.89) |
| AC021/22 | OQ955853 | 860 | A. baumannii (99.88) |
| AC022/22 | OQ955854 | 884 | A. baumannii (99.77) |
| AC023/22 | OQ955855 | 845 | A. baumannii (99.88) |
| AC024/22 | OQ968344 | 1.036 | A. baumannii (99.71) |
| AC025/22 | OQ968345 | 1.034 | A. baumannii (99.90) |
| AC026.22 | OQ968346 | 1.037 | A. baumannii (99.90) |
| AC027/22 | OQ968347 | 873 | A. baumannii (99.88) |
| AC028/22 | OQ968348 | 945 | A. baumannii (99.68) |
| AC029/22 | OQ968349 | 1.034 | A. baumannii (99.90) |
| AC030/22 | OQ968350 | 1.043 | A. baumannii (99.61) |
| AC031/22 | OQ968351 | 1.043 | A. baumannii (99.90) |
| AC032/22 | OQ968352 | 1.033 | A. baumannii (99.90) |
| AC033/22 | OQ970134 | 1.031 | A. baumannii (99.90) |
| AC034/22 | OQ970135 | 1.034 | A. baumannii (99.90) |
| AC035/22 | OQ970136 | 1.042 | A. baumannii (99.90) |
| AC036/22 | OQ970137 | 453 | A. baumannii (99.77) |
| AC037/22 | OQ970138 | 1.031 | A. baumannii (99.90) |
| AC038/22 | OQ970139 | 1.032 | A. baumannii (99.90) |
| AC039/22 | OQ970140 | 1.033 | A. baumannii (99.90) |
| AC040/22 | OQ970141 | 1.032 | A. baumannii (99.90) |
| AC041/22 | OQ970142 | 1.020 | A. baumannii (90.90) |
| AC042/22 | OQ970143 | 1.035 | A. baumannii (90.90) |
| AC043/22 | OQ970144 | 1.034 | A. baumannii (90.90) |
| AC044/22 | OQ970145 | 1.003 | A. baumannii (90.90) |
| AC045/22 | OQ970146 | 1.034 | A. baumannii (90.90) |
| AC046/22 | PP373676 | 1.306 | A. baumannii (90.90) |
| AC047/22 | OQ970147 | 1.006 | A. baumannii (100) |
| AC048/22 | OQ970148 | 1.003 | A. baumannii (100) |
Appendix A.2
| Strains | AMS | TZP | FOX | CAZ | CRO | CEF | IPM | MEM | GEN | CIP | TIG | SXT | AMI | Magiorakos et al. [1] Classification |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AC001.21 | I | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC002.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC003.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC004.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC005.21 | R | R | R | R | R | R | R | R | R | R | S | R | S | XDR |
| AC006.21 | R | R | R | R | I | R | R | R | R | R | R | R | R | XDR |
| AC007.21 | R | R | R | R | I | R | R | R | R | R | R | R | R | XDR |
| AC008.21 | R | R | R | R | I | R | R | R | R | R | R | R | R | XDR |
| AC010.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC012.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC013.21 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC014.21 | S | R | R | R | I | R | R | R | R | R | R | R | R | XDR |
| AC015.21 | S | S | R | S | I | S | S | S | S | I | S | S | S | N-MDR |
| AC016.21 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC017.21 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC018.22 | R | R | R | R | R | R | R | R | R | R | I | R | S | XDR |
| AC019.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC020.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC021.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC022.22 | S | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC023.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC024.22 | R | R | R | R | R | R | R | R | S | R | I | S | R | MDR |
| AC025.22 | R | R | R | R | R | R | R | R | S | R | S | R | S | MDR |
| AC026.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC027.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC028.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC029.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC030.22 | R | R | R | R | R | R | R | R | R | R | R | S | S | XDR |
| AC031.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC032.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC033.22 | R | R | R | R | R | R | R | R | R | R | R | R | R | XDR |
| AC034.22 | R | R | R | R | R | R | R | R | R | R | R | R | R | XDR |
| AC035.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC036.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC037.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC038.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC039.22 | R | R | R | R | R | R | R | R | R | R | R | R | R | XDR |
| AC040.22 | R | R | R | R | R | R | R | R | R | R | R | R | R | XDR |
| AC041.22 | I | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC042.22 | R | R | R | R | R | R | R | R | R | R | R | R | R | XDR |
| AC043.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC044.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC045.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
| AC046.22 | R | R | R | R | R | R | R | R | R | R | S | R | R | XDR |
| AC047.22 | S | S | I | S | S | S | R | S | R | R | S | R | S | MDR |
| AC048.22 | R | R | R | R | R | R | R | R | R | R | I | R | R | XDR |
References
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.A.; Ozer, E.; Bolon, M.K.; Hauser, A.R. Influence of ACB complex genospecies on clinical outcomes in a U.S. hospital with high rates of multidrug resistance. J. Infect. 2015, 70, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55–90. [Google Scholar] [CrossRef]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef] [PubMed]
- Alenazi, T.A.; Bin Shaman, M.S.; Suliman, D.M.; Alanazi, T.A.; Altawalbeh, S.M.; Alshareef, H.; Lahreche, D.I.; Al-Azzam, S.; Araydah, M.; Karasneh, R.; et al. The Impact of Multidrug-Resistant Acinetobacter baumannii Infection in Critically Ill Patients with or without COVID-19 Infection. Healthcare 2023, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- FAO (United Nations Food and Agriculture Organization). The FAO Action Plan on Antimicrobial Resistance 2016–2020; FAO: Rome, Italy, 2016. [Google Scholar]
- Ergonul, O.; Tokca, G.; Keske, Ş.; Donmez, E.; Madran, B.; Kömür, A.; Gönen, M.; Can, F. Elimination of healthcare-associated Acinetobacter baumannii infection in a highly endemic region. Int. J. Infect. Dis. 2022, 114, 11–14. [Google Scholar] [CrossRef]
- WHO. World Health Organization (WHO). WHO Bacterial Priority Pathogens List. WHO. 2024. Available online: https://www.who.int/publications/i/item/9789240093461 (accessed on 20 February 2025).
- Yang, C.-H.; Su, P.-W.; Moi, S.-H.; Chuang, L.-Y. Biofilm Formation in Acinetobacter baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef]
- Choudhary, M.; Shrivastava, R.; Vashistt, J. Acinetobacter baumannii Biofilm Formation: Association with Antimicrobial Resistance and Prolonged Survival under Desiccation. Curr. Microbiol. 2022, 79, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Chowdhury, G.; Mukhopadhyay, A.K.; Dutta, S.; Basu, S. Convergence of Biofilm Formation and Antibiotic Resistance in Acinetobacter baumannii Infection. Front. Med. 2022, 9, 793615. [Google Scholar] [CrossRef]
- Oho, M.; Nagasawa, Z.; Funashima, Y.; Ueda, O.; Watamabe, S.; Cui, L.; Miyamoto, H.; Sueoka, E. Correlation of Strain Classification with IR Biotyper and Molecular Epidemiological Method of Pseudomonas aeruginosa. Rinsho Biseibutshu Jinsoku Shindan Kenkyukai Shi 2021, 31, 29–40. [Google Scholar] [PubMed]
- Martak, D.; Valot, B.; Sauget, M.; Cholley, P.; Thouverez, M.; Bertrand, X.; Hocquet, D. Fourier-Transform InfraRed Spectroscopy Can Quickly Type Gram-Negative Bacilli Responsible for Hospital Outbreaks. Front. Microbiol. 2019, 10, 1440. [Google Scholar] [CrossRef]
- Guerrero-Lozano, I.; Galán-Sánchez, F.; Rodríguez-Iglesias, M. Fourier transform infrared spectroscopy as a new tool for surveillance in local stewardship antimicrobial program: A retrospective study in a nosocomial Acinetobacter baumannii outbreak. Braz. J. Microbiol. 2022, 53, 1349–1353. [Google Scholar] [CrossRef]
- Moubareck, C.A.; Halat, D.H. Insights into Acinetobacter baumannii: A review of microbiological, virulence, and resistance traits in a threatening nosocomial pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Čiginskienė, A.; Dambrauskienė, A.; Rello, J.; Adukauskienė, D. Ventilator-associated pneumonia due to drug-resistant Acinetobacter baumannii: Risk factors and mortality relation with resistance profiles, and independent predictors of in-hospital mortality. Medicina 2019, 55, 49. [Google Scholar] [CrossRef]
- Rangel, K.; Chagas, T.P.G.; De-Simone, S.G. Acinetobacter baumannii Infections in Times of COVID-19 Pandemic. Pathogens 2021, 10, 1006. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, I.; Lupia, T.; Shbaklo, N.; Bianchi, A.; Concialdi, E.; Penna, M.; Corcione, S.; De Rosa, F.G. Prognostic evaluation of Acinetobacter baumannii ventilator-associated pneumonia in COVID-19. Infez Med. 2022, 30, 570–576. [Google Scholar] [PubMed]
- Russo, A.; Gavaruzzi, F.; Ceccarelli, G.; Borrazzo, C.; Oliva, A.; Alessandri, F.; Magnanimi, E.; Pugliese, F.; Venditti, M. Multidrug-resistant Acinetobacter baumannii infections in COVID-19 patients hospitalized in intensive care unit. Infection 2021, 50, 83–92. [Google Scholar] [CrossRef]
- Novović, K.; Nedeljković, S.K.; Poledica, M.; Nikolić, G.; Grujić, B.; Jovčić, B.; Kojić, M.; Filipić, B. Virulence potential of multidrug-resistant Acinetobacter baumannii isolates from COVID-19 patients on mechanical ventilation: The first report from Serbia. Front. Microbiol. 2023, 14, 1094184. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Tao, S.; Ying, H.; Fang, Q.; Kong, J.; Guo, F.; Yang, Y.; Cao, P.; Zhou, Y.; Jin, W.; et al. Molecular pathogen profiling of COVID-19 coinfections. BMC Infect. Dis. 2024, 24, 1451. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; CLSI: Berwyn, PA, USA, 2020. [Google Scholar]
- Vasconcellos, L.; Silva, S.V.; da Costa, L.V.; da Silva Lage de Miranda, R.V.; Reis, C.M.F.D.; da Silva Braga, L.M.P.; Silva, C.; Conceição, G.; Mattoso, J.; Silva, I.B.; et al. Phenotypical and molecular characterization of Acinetobacter spp. isolated from a pharmaceutical facility. Lett. Appl. Microbiol. 2023, 76, ovad101. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Chezzi, C. MALDI-TOF MS: A Reliable Tool in the Real Life of the Clinical Microbiology Laboratory. Microorganisms 2024, 12, 322. [Google Scholar] [CrossRef] [PubMed]
- Toh, B.E.; Paterson, D.L.; Kamolvit, W.; Zowawi, H.; Kvaskoff, D.; Sidjabat, H.; Wailan, A.; Peleg, A.Y.; Huber, C.A. Species identification within Acinetobacter calcoaceticus–baumannii complex using MALDI-TOF MS. J. Microbiol. Methods 2015, 118, 128–132. [Google Scholar] [CrossRef] [PubMed]
- de Souza, P.A.; Santos, M.C.S.d.; de Miranda, R.V.d.S.L.; da Costa, L.V.; da Silva, R.P.P.; de Miranda, C.A.C.; da Silva, A.P.R.; Forsythe, S.J.; Bôas, M.H.S.V.; Brandão, M.L.L. Evaluation of antimicrobial resistance patterns of Pseudomonas aeruginosa strains isolated among COVID-19 patients in Brazil typed by Fourier-transform infrared spectroscopy. Life 2024, 14, 1079. [Google Scholar] [CrossRef]
- Lombardo, D.; Cordovana, M.; Deidda, F.; Pane, M.; Ambretti, S. Application of Fourier Transform Infrared Spectroscopy for Real-Time Typing of Acinetobacter baumannii Outbreak in Intensive Care Unit. Futur. Microbiol. 2021, 16, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-J.; Xiao, Z.-G.; Lv, X.-J.; Huang, H.-T.; Liao, C.; Hui, C.-Y.; Xu, Y.; Li, H.-F. Drug-resistant Acinetobacter baumannii: From molecular mechanisms to potential therapeutics (Review). Exp. Ther. Med. 2023, 25, 209. [Google Scholar] [CrossRef]
- Falagas, M.E.; Vardakas, K.Z.; Roussos, N.S. Trimethoprim/sulfamethoxazole for Acinetobacter spp.: A review of current microbiological and clinical evidence. Int. J. Antimicrob Agents 2015, 46, 231–241. [Google Scholar] [CrossRef]
- Al-Tamimi, M.; Albalawi, H.; Alkhawaldeh, M.; Alazzam, A.; Ramadan, H.; Altalalwah, M.; Alma’aitah, A.; Al Balawi, D.A.; Shalabi, S.; Abu-Raideh, J.; et al. Multidrug-resistant Acinetobacter baumannii in Jordan. Microorganisms 2022, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Dehbanipour, R.; Ghalavand, Z. Acinetobacter baumannii: Pathogenesis, virulence factors, novel therapeutic options and mechanisms of resistance to antimicrobial agents with emphasis on tigecycline. J. Clin. Pharm. Ther. 2022, 47, 1875–1884. [Google Scholar] [CrossRef]
- Yaghoubi, S.; Zekiy, A.O.; Krutova, M.; Gholami, M.; Kouhsari, E.; Sholeh, M.; Ghafouri, Z.; Maleki, F. Tigecycline antibacterial activity, clinical effectiveness, and mechanisms and epidemiology of resistance: Narrative review. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1003–1022. [Google Scholar] [CrossRef]
- Hua, X.; He, J.; Wang, J.; Zhang, L.; Zhang, L.; Xu, Q.; Shi, K.; Leptihn, S.; Shi, Y.; Fu, X.; et al. Novel tigecycline resistance mechanisms in Acinetobacter baumannii mediated by mutations in adeS, rpoB and rrf. Emerg Microbes Infect 2021, 10, 1404–1417. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.E.; Grim, S.A.; Aldeza, C.A.; Janda, W.M.; Clark, N.M. Rapid Development of Acinetobacter baumannii Resistance to Tigecycline. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2007, 27, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- FDA. Drug safety communication: Increased risk of death with Tygacil (tigecycline) compared to other antibiotics used to treat similar infections (1 September 2010). In FDA Drug Safety Communication: FDA Warns of Increased Risk of Death with IV Antibacterial Tygacil (Tigecycline) and Approves New Boxed Warning; US Food and Drug Administration: Silver Spring, MD, USA, 2017. [Google Scholar]
- Shi, J.; Sun, T.; Cui, Y.; Wang, C.; Wang, F.; Zhou, Y.; Miao, H.; Shan, Y.; Zhang, Y. Multidrug resistant and extensively drug resistant Acinetobacter baumannii hospital infection associated with high mortality: A retrospective study in the pediatric intensive care unit. BMC Infect. Dis. 2020, 20, 597. [Google Scholar] [CrossRef] [PubMed]
- Loyola-Cruz, M.Á.; Durán-Manuel, E.M.; Cruz-Cruz, C.; Márquez-Valdelamar, L.M.; Bravata-Alcántara, J.C.; Cortés-Ortíz, I.A.; Cureño-Díaz, M.A.; Ibáñez-Cervantes, G.; Fernández-Sánchez, V.; Castro-Escarpulli, G.; et al. ESKAPE bacteria characterization reveals the presence of Acinetobacter baumannii and Pseudomonas aeruginosa outbreaks in COVID-19/VAP patients. Am. J. Infect Control 2023, 51, 729–737. [Google Scholar] [CrossRef]

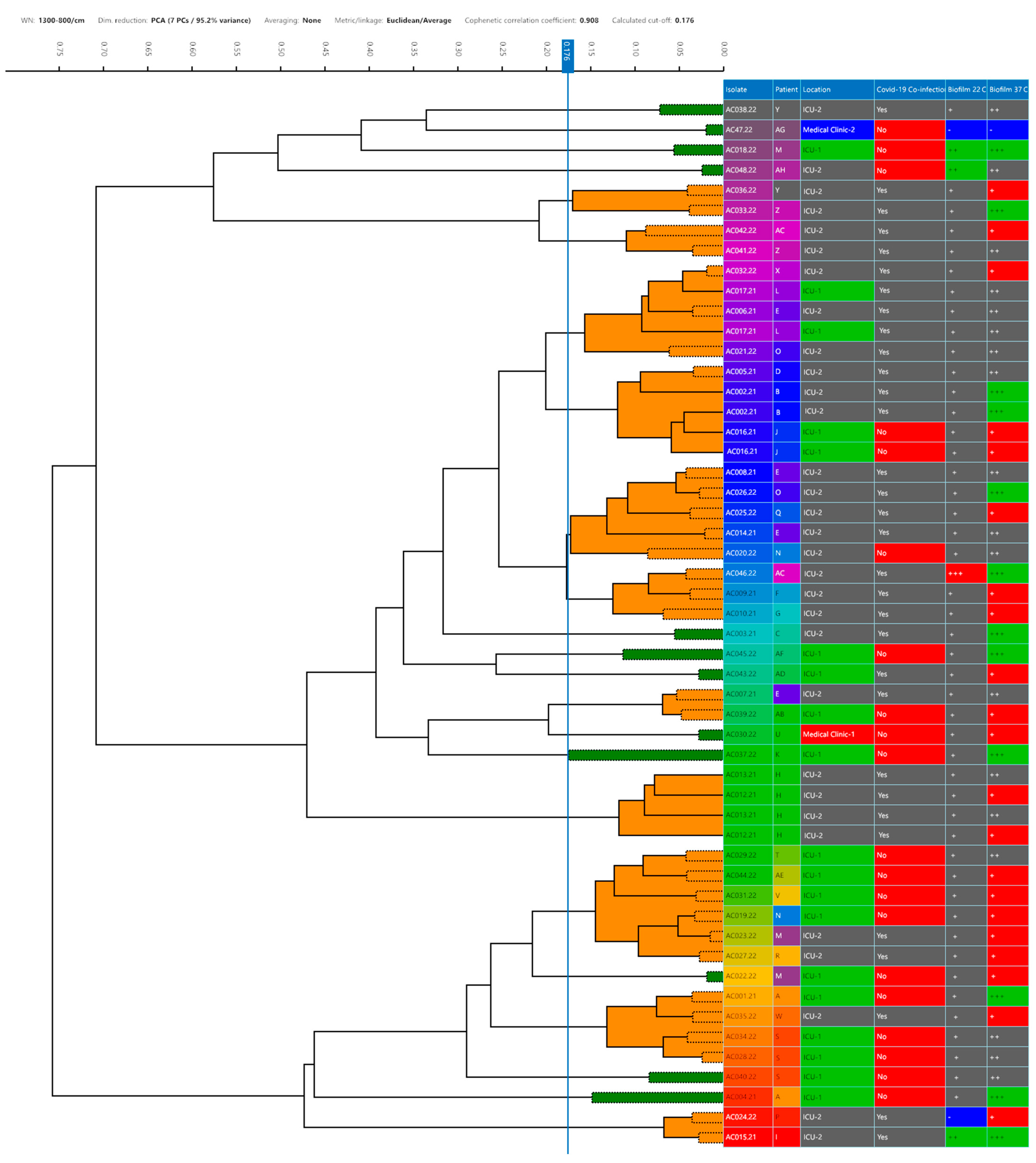
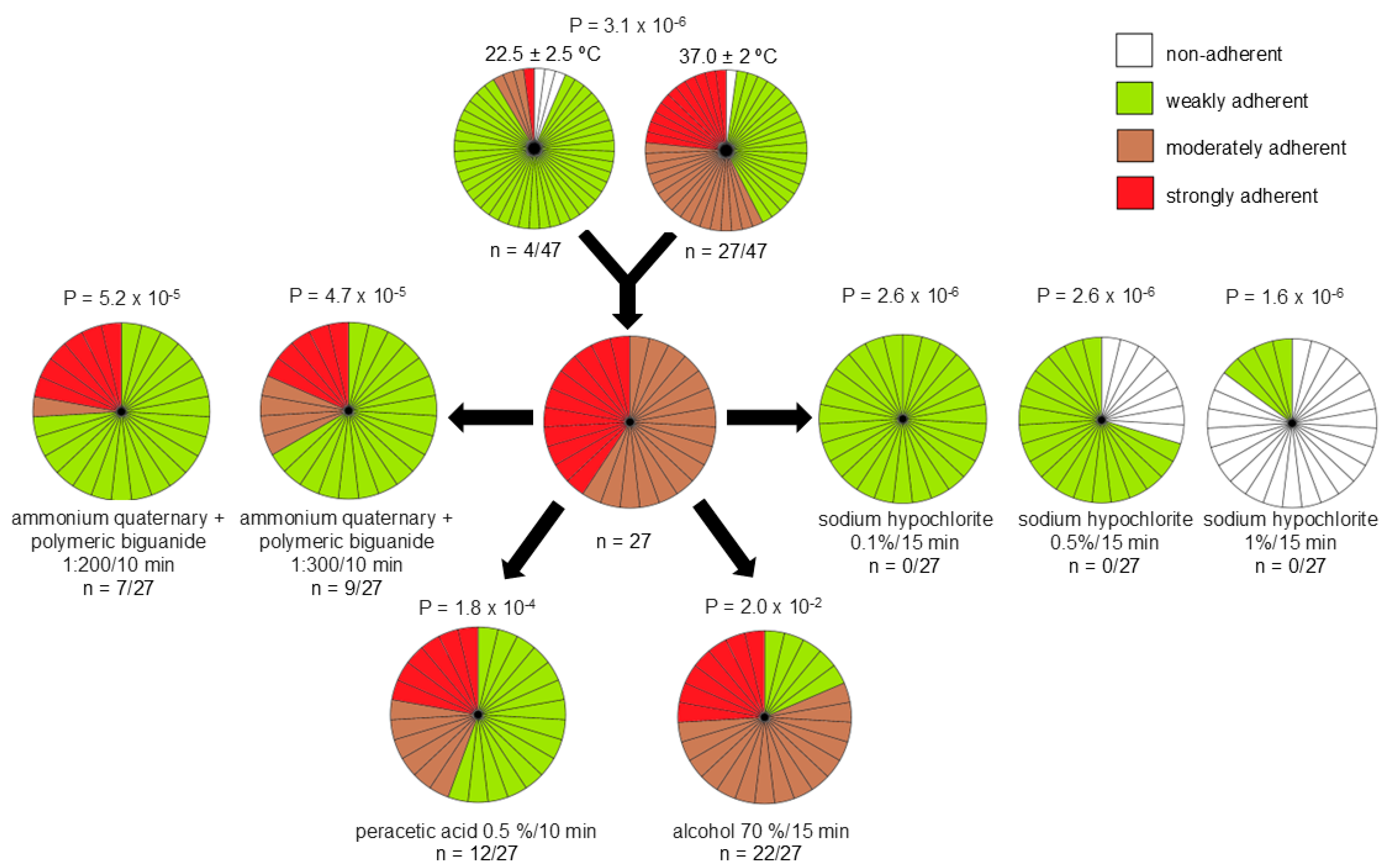
| Strain (CBAS a ID) | Source | Patient ID | Hospital Department | COVID-19 Co-Infection | Isolation Date | VITEK® 2 Bionumber (Profile) | MALDI-TOF MS b | |
|---|---|---|---|---|---|---|---|---|
| VITEK MS® (%) | MALDI Biotyper® (Score) | |||||||
| AC001/21 (936) | Tracheal swab | A | ICU c-1 | no | 21 June | 0241010103500250 (I) | A. baumannii (98.4) | A. baumannii (2.43) |
| AC002/21(937) | Tracheal swab | B | ICU-2 | yes | 21 June | 0201010103500210 (II) | A. baumannii (99.9) | A. baumannii (2.33) |
| AC003/21 (938) | Tracheal swab | C | ICU-2 | yes | 21 June | 0241010103500352 (III) | A. baumannii (99.9) | A. baumannii (2.43) |
| AC004/21 (939) | Tracheal swab | A | ICU-1 | no | 21 June | 0241010103500210 (IV) | A. baumannii (95.0) | A. baumannii (2.31) |
| AC005/21 (940) | Blood | D | ICU-2 | yes | 21 June | 0201010103500352 (V) | A. baumannii (99.9) | A. baumannii (2.36) |
| AC006/21 (941) | Blood | E | ICU-2 | yes | 21 June | 0201010103500312 (VI) | A. baumannii (99.9) | A. baumannii (2.28) |
| AC007/21 (942) | Blood | E | ICU-2 | yes | 21 June | 0241010103500352 (III) | A. baumannii (99.9) | A. baumannii (2.04) |
| AC008/21 (943) | Blood | E | ICU-2 | yes | 21 June | 0241010103500310 (VII) | A. baumannii (81.7) | A. baumannii (2.25) |
| AC009/21 (944) | Urine | F | ICU-2 | yes | 21 July | 0201010103500210 (II) | A. baumannii (99.9) | A. baumannii (2.33) |
| AC010/21 (945) | Tracheal secretion | G | ICU-2 | Yes | 21 July | 0201010103500210 (II) | A. baumannii (99.9) | A. baumannii (2.38) |
| AC012/21 (946) | Urine | H | ICU-2 | Yes | 21 July | 0241010103500310 (VII) | A. baumannii (99.9) | A. baumannii (2.21) |
| AC013/21 (947) | Tracheal secretion | H | ICU-2 | Yes | 21 July | 0201010103500352 (V) | A. baumannii (99.9) | A. baumannii (2.22) |
| AC014/21 (948) | Blood | E | ICU-2 | Yes | 21 July | 0241010003500312 (VIII) | A. baumannii (99.9) | A. baumannii (2.32) |
| AC015/21 (949) | Blood | I | ICU-2 | Yes | 21 July | 0201010103500312 (VI) | A. baumannii (97.0) | A. baumannii (2.29) |
| AC016/21 (950) | Blood | J | ICU-1 | No | 21 July | 0241010103500310 (VII) | A. baumannii (99.9) | A. baumannii (2.26) |
| AC017/21 (951) | Blood | L | ICU-2 | Yes | 21 September | 0241010103500210 (IV) | A. baumannii (99.9) | A. baumannii (2.15) |
| AC018/22 (952) | Rectal swab | M | ICU-1 | No | 22 January | 0241010103500250 (I) | A. baumannii (81.3) | A. baumannii (2.25) |
| AC019/22 (953) | Rectal swab | N | ICU-1 | No | 22 January | 0241010003500210 (IX) | A. baumannii (83.0) | A. baumannii (2.37) |
| AC020/22 (954) | Tracheal secretion | N | ICU-1 | No | 22 February | 0241010103500212 (X) | A. baumannii (99.4) | A. baumannii (2.23) |
| AC021/22 (955) | Tracheal secretion | O | ICU-2 | Yes | 22 February | 0241010103500352 (III) | A. baumannii (99.9) | A. baumannii (2.33) |
| AC022/22 (956) | Urine | M | ICU-1 | No | 22 January | 0241010103500210 (IV) | A. baumannii (99.9) | A. baumannii (2.36) |
| AC023/22 (957) | Tracheal secretion | M | ICU-2 | Yes | 22 February | 0241010103500210 (IV) | A. baumannii (99.9) | A. baumannii (2.02) |
| AC024/22 (958) | Tracheal secretion | P | ICU-2 | Yes | 22 February | 0241010103500312 (XI) | A. baumannii (99.9) | A. baumannii (2.36) |
| AC025/22 (959) | Tracheal secretion | Q | ICU-2 | Yes | 22 February | 0241010003500310 (XII) | A. baumannii (99.9) | A. baumannii (2.32) |
| AC026/22 (960) | Blood | O | ICU-2 | Yes | 22 February | 0241010103500310 (VII) | A. baumannii (98.0) | A. baumannii (2.41) |
| AC027/22 (961) | Blood | R | ICU-2 | Yes | 22 February | 0201010103500210 (II) | A. baumannii (99.9) | A. baumannii (2.00) |
| AC028/22 (962) | Tracheal secretion | S | ICU-1 | no | 22 February | 0201010003500210 (XIII) | A. baumannii (99.9) | A. baumannii (2.31) |
| AC029/22 (963) | Tracheal secretion | T | ICU-1 | no | 22 February | 0241010103500310 (VII) | A. baumannii (99.9) | A. baumannii (2.24) |
| AC030/22 (964) | Tracheal secretion | U | Medical clinic-1 | no | 22 February | 0241010003500310 (XII) | A. baumannii (99.9) | A. baumannii (2.41) |
| AC031/22 (965) | Blood | V | ICU-1 | no | 22 February | 0241010103500210 (IV) | A. baumannii (88.8) | A. baumannii (2.29) |
| AC032/22 (966) | Rectal swab | X | ICU-2 | yes | 22 February | 0243051103500352 (XIV) | A. baumannii (99.5) | A. baumannii (2.46) |
| AC033/22 (967) | Rectal swab | Z | ICU-2 | yes | 22 February | 0201010003500210 (XIII) | A. baumannii (99.9) | A. baumannii (2.13) |
| AC034/22 (968) | Tracheal secretion | S | ICU-1 | no | 22 February | 0241010103500210 (IV) | A. baumannii (99.9) | A. baumannii (2.44) |
| AC035/22 (969) | Rectal swab | W | ICU-2 | yes | 22 February | 0241010003500210 (IX) | A. baumannii (96.2) | A. baumannii (2.20) |
| AC036/22 (970) | Urine | Y | ICU-2 | yes | 22 February | 0201010001500210 (XV) | A. baumannii (99.9) | A. baumannii (2.36) |
| AC037/22 (971) | Tracheal secretion | K | ICU-1 | no | 22 March | 0241010103500352 (III) | A. baumannii (96.4) | A. baumannii (2.40) |
| AC038/22 (672) | Tracheal secretion | Y | ICU-2 | yes | 22 March | 0241010103500310 (VII) | A. baumannii (99.9) | A. baumannii (2.45) |
| AC039/22 (973) | Tracheal secretion | AB | ICU-1 | no | 22 February | 0241010103500310 (VII) | A. baumannii (95.4) | A. baumannii (2.07) |
| AC040/22 (974) | Tracheal secretion | S | ICU-1 | no | 22 February | 0241010103500210 (IV) | A. baumannii (91.9) | A. baumannii (2.01) |
| AC041/22 (975) | Urine | Z | ICU-2 | yes | 22 March | 0241010103500210 (IV) | A. baumannii (88.8) | A. baumannii (2.49) |
| AC042/22 (976) | Rectal swab | AC | ICU-2 | yes | 22 March | 0241010003500210 (IX) | A. baumannii (99.9) | A. baumannii (2.17) |
| AC043/22 (977) | Rectal swab | AD | ICU-2 | yes | 22 March | 0241010103500310 (VII) | A. baumannii (92.0) | A. baumannii (2.25) |
| AC044/22 (978) | Rectal swab | AE | ICU-1 | no | 22 March | 0201010003500210 (XIII) | A. baumannii (99.9) | A. baumannii (2.44) |
| AC045/22 (979) | Rectal swab | AF | ICU-1 | no | 22 March | 0201010103500312 (VI) | A. baumannii (91.8) | A. baumannii (2.30) |
| AC046/22 (1030) | Tracheal secretion | AC | ICU-2 | yes | 22 March | 0201010103500210 (II) | A. baumannii (92.0) | A. baumannii (2.20) |
| AC047.22 (980) | Urine | AG | Medical clinic-2 | no | 22 March | 0241411103500353 (XVI) | A. baumannii (99.9) | A. baumannii (2.25) |
| AC048.22 (981) | Rectal swab | AH | ICU-2 | no | 22 March | 0241010103500310 (VII) | A. baumannii (99.9) | A. baumannii (2.05) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, P.A.; Santos, M.C.N.d.; Miranda, R.V.d.S.L.d.; da Costa, L.V.; da Silva, R.P.P.; Miranda, C.A.C.d.; Conceição, G.M.S.d.; Forsythe, S.J.; Villas Bôas, M.H.S.; Brandão, M.L.L. Phenotypical and Molecular Characterization of Acinetobacter baumannii Isolated from Hospitalized Patients During the COVID-19 Pandemic in Brazil. Life 2025, 15, 623. https://doi.org/10.3390/life15040623
de Souza PA, Santos MCNd, Miranda RVdSLd, da Costa LV, da Silva RPP, Miranda CACd, Conceição GMSd, Forsythe SJ, Villas Bôas MHS, Brandão MLL. Phenotypical and Molecular Characterization of Acinetobacter baumannii Isolated from Hospitalized Patients During the COVID-19 Pandemic in Brazil. Life. 2025; 15(4):623. https://doi.org/10.3390/life15040623
Chicago/Turabian Stylede Souza, Paula Araujo, Milena Cristina Nunes dos Santos, Rebeca Vitória da Silva Lage de Miranda, Luciana Veloso da Costa, Raphael Paiva Paschoal da Silva, Cátia Aparecida Chaia de Miranda, Greice Maria Silva da Conceição, Stephen James Forsythe, Maria Helena Simões Villas Bôas, and Marcelo Luiz Lima Brandão. 2025. "Phenotypical and Molecular Characterization of Acinetobacter baumannii Isolated from Hospitalized Patients During the COVID-19 Pandemic in Brazil" Life 15, no. 4: 623. https://doi.org/10.3390/life15040623
APA Stylede Souza, P. A., Santos, M. C. N. d., Miranda, R. V. d. S. L. d., da Costa, L. V., da Silva, R. P. P., Miranda, C. A. C. d., Conceição, G. M. S. d., Forsythe, S. J., Villas Bôas, M. H. S., & Brandão, M. L. L. (2025). Phenotypical and Molecular Characterization of Acinetobacter baumannii Isolated from Hospitalized Patients During the COVID-19 Pandemic in Brazil. Life, 15(4), 623. https://doi.org/10.3390/life15040623