

Evaluating the Impact of rs4025935, rs71748309, rs699947, and rs4646994 Genetic Determinants on Polycystic Ovary Syndrome Predisposition—A Case-Control Study



Abstract
1. Introduction
2. Methodology
2.1. Study Participants and Criteria
2.2. Biochemical Serum Profile: For the First Phase, We Determined Biochemical Serum Profiles of the Patients, Such as HbA1c, Fasting Glucose, Insulin, Serum Lipids, and Hormones
2.3. Extraction and Qualitative Assessment of Genomic DNA
2.4. Genotyping of VEGF rs4646994 I/D, VEGF rs699947 C/A, GSTM1 rs4025935, and GSTT1 rs71748309 Genes
2.5. Preparation of PCR Cocktail
2.6. PCR Program
2.7. PCR Product Visualization in Gels
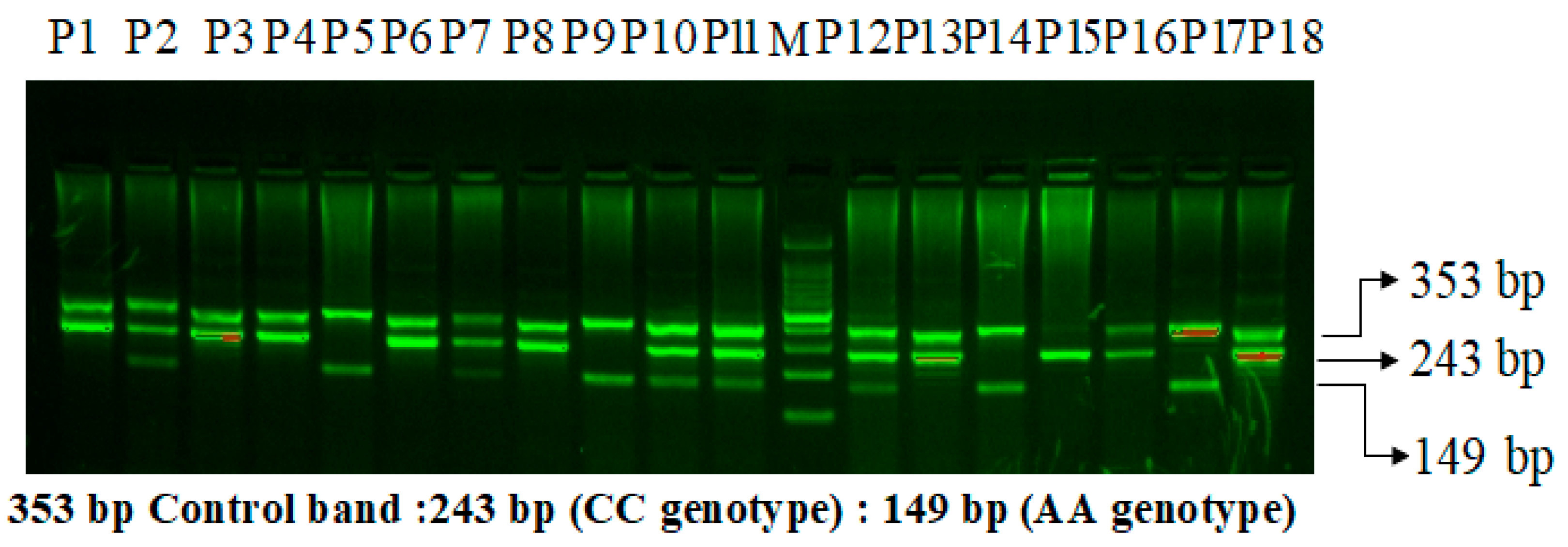
2.8. VEGF-rs699947 (-2578) C>A Genotyping
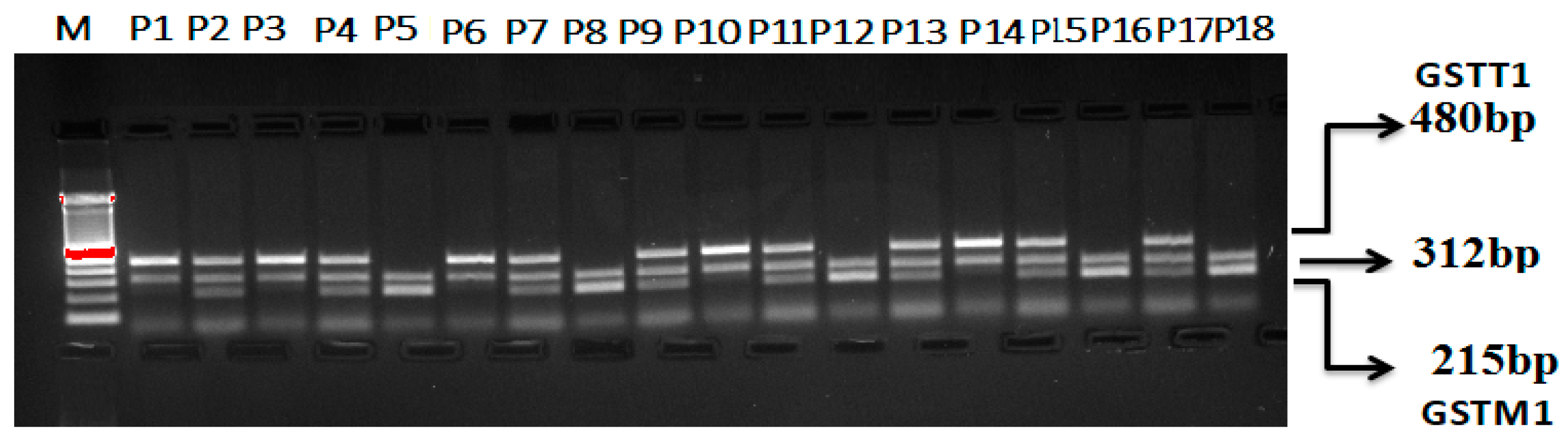
Multiplex PCR for GSTT1 and GSTM1 (rs4025935 and rs71748309) Genotyping
3. Results
4. Discussion
Comparative Association of VEGF rs699947 C>A Genotypes in PCOS Patients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stein, I.F.; Leventhal, M.L. Amenorrhea associated with bilateral polycystic ovaries. Am. J. Obstet. Gynecol. 1935, 29, 181–191. [Google Scholar] [CrossRef]
- Sirmans, S.M.; Pate, K.A. Epidemiology, diagnosis, and management of polycystic ovary syndrome. Clin. Epidemiol. 2013, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, F.R.; Behboudi-Gandevani, S. Polycystic Ovary Syndrome. In Contemporary Gynecologic Practice; InTech: Rijeka, Croatia, 2015. [Google Scholar] [CrossRef]
- Ndefo, U.A.; Eaton, A.; Green, M.R. Polycystic ovary syndrome: A review of treatment options with a focus on pharmacological approaches. Pharm. Ther. Peer-Rev. J. Formul. Manag. 2013, 38, 336–355. [Google Scholar]
- Hart, R.; Hickey, M.; Franks, S. Definitions, prevalence and symptoms of polycystic ovaries and polycystic ovary syndrome. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 671–683. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhou, Q.; Guo, X.; Xu, C.; Jia, X.; Cao, Z.; Lu, Q. From Pathophysiology to Treatment: The Role of Ferroptosis in PCOS. Front. Biosci. (Landmark Ed) 2025, 30, 25586. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, Z.; Yu, Y.; Wang, X.; Zhao, Y. Natural compounds in the management of polycystic ovary syndrome: A comprehensive review of hormonal regulation and therapeutic potential. Front. Nutr. 2025, 12, 1520695. [Google Scholar] [CrossRef] [PubMed]
- Xita, N.; Georgiou, I.; Tsatsoulis, A. The genetic basis of polycystic ovary syndrome. Eur. J. Endocrinol. 2002, 147, 717–725. [Google Scholar] [CrossRef]
- Ajmal, N.; Khan, S.Z.; Shaikh, R. Polycystic ovary syndrome (PCOS) and genetic predisposition: A review article. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 3, 100060. [Google Scholar] [CrossRef]
- Azevedo, M.M.P.; de Marqui, A.B.T.; Bacalá, B.T.; Balarin, M.A.S.; de Resende, E.A.M.R.; Lima, M.F.P.; Gomes, M.K.d.O.; Cintra, M.T.R. Polymorphisms of the GSTT1 and GSTM1 genes in polycystic ovary syndrome. Rev. Assoc. Médica Bras. 2020, 66, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Murri, M.; Luque-Ramírez, M.; Insenser, M.; Ojeda-Ojeda, M.; Escobar-Morreale, H.F. Circulating markers of oxidative stress and polycystic ovary syndrome (PCOS): A systematic review and meta-analysis. Hum. Reprod. Update 2013, 19, 268–288. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.; Prasad, N.R.; Manohar, S.M.; Sachan, A.; Narasimha, S.R.P.V.L.; Bitla, A.R.R. Oxidative stress in non-obese women with polycystic ovarian syndrome. J. Clin. Diagn. Res. JCDR 2014, 8, CC01–CC03. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Rambhia, P.; Gishto, A. Human embryonic stem cell cultivation: Historical perspective and evolution of xeno-free culture systems. Reprod. Biol. Endocrinol. 2015, 13, 9. [Google Scholar] [CrossRef]
- Abu-Duhier, F.; Mir, R. GSTT1 (rs4025935) null genotype is associated with increased risk of sickle cell disease in the populations of Tabuk-Northwestern region of Saudi Arabia. Hematology 2017, 22, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Sam, S. Obesity and Polycystic Ovary Syndrome. Obes. Manag. 2007, 3, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Kandarakis, H.; Legro, R.S. The Role of Genes and Environment in the Etiology of PCOS. Endocrine 2006, 30, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.S.; Marqui, A.B.T.D.; Teles, D.R.F.; Resende, E.A.M.R.; Lima, M.F.P.; Gomes, M.K.O.; Cintra, M.T.R. VEGF gene rs35569394 polymorphism in patients with Polycystic Ovary Syndrome. Rev. Assoc. Médica Bras. 2020, 66, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar] [CrossRef]
- Di Pietro, M.; Pascuali, N.; Parborell, F.; Abramovich, D. Ovarian angiogenesis in polycystic ovary syndrome. Reproduction 2018, 155, R199–R209. [Google Scholar] [CrossRef]
- Albalawi, I.A.; Mir, R.; Duhier, F.A. Genetic effects of Vascular Endothelial Growth Factor A (VEGF-A) and Its Association with Disease Progression in Breast Cancer Population of Saudi Arabia. Asian Pac. J. Cancer Prev. APJCP 2020, 21, 139–145. [Google Scholar] [CrossRef]
- Kshetrimayum, C.; Sharma, A.; Mishra, V.V.; Kumar, S. Polycystic ovarian syndrome: Environmental/occupational, lifestyle factors; an overvie. J. Turk. Ger. Gynecol. Assoc. 2019, 20, 255–263. [Google Scholar] [CrossRef]
- Vicente, A.L.S.A.; Marqui, A.B.T.; Gomes, M.K.O.; Assunção-Luiz, A.V.; Balarin, M.A.S.; Tanaka, S.C.S.V.; Resende, E.A.M.R.; Lima, M.F.P.; Cintra, M.T.R. Polymorphisms rs2010963 and rs833061 of the VEGF gene in polycystic ovary syndrome. Rev. Assoc. Med. Bras. (1992) 2022, 68, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Elfaki, I.; Mir, R.; Elnageeb, M.E.; Hamadi, A.; Alharbi, Z.M.; Bedaiwi, R.I.; Javid, J.; Alrasheed, T.; Alatawi, D.; Alrohaf, B.M.; et al. Identification of Interactive Genetic Loci Linked to Insulin Resistance in Metabolic Syndrome-An Update. Medicina (Kaunas) 2025, 61, 83. [Google Scholar] [CrossRef]
- Gao, J.; Song, Y.; Huang, X.; Wang, D.; Wang, H. The expression of platelet-derived growth factor, epidermal growth factor, and insulin-like growth factor-II in patients with polycystic ovary syndrome and its correlation with pregnancy outcomes. Ann. Palliat. Med. 2021, 10, 5671–5678. [Google Scholar] [CrossRef] [PubMed]
- Artini, P.G.; Monti, M.; Matteucci, C.; Valentino, V.; Cristello, F.; Genazzani, A.R. Vascular endothelial growth factor and basic fibroblast growth factor in polycystic ovary syndrome during controlled ovarian hyperstimulation. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2006, 22, 465–470. [Google Scholar] [CrossRef]
- Fagiani, E.; Christofori, G. Angiopoietins in angiogenesis. Cancer Lett. 2013, 328, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Abramovich, D.; Irusta, G.; Bas, D.; Cataldi, N.I.; Parborell, F.; Tesone, M. Angiopoietins/TIE2 system and VEGF are involved in ovarian function in a DHEA rat model of polycystic ovary syndrome. Endocrinology 2012, 153, 3446–3456. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, C.; Lilford, R.J.; Marjoribanks, J.; Vandekerckhove, P. Laparoscopic ‘drilling’ by diathermy or laser for ovulation induction in anovulatory polycystic ovary syndrome. Cochrane Database Syst. Rev. 2007, 18, CD001122. [Google Scholar]
- Ben Salem, A.; Megdich, F.; Kacem, O.; Souayeh, M.; Hachani Ben Ali, F.; Hizem, S.; Janhai, F.; Ajina, M.; Abu-Elmagd, M.; Assidi, M.; et al. Vascular endothelial growth factor (VEGFA) gene variation in polycystic ovary syndrome in a Tunisian women population. BMC Genom. 2016, 17 (Suppl. S9), 748. [Google Scholar] [CrossRef]
- Al Balawi, I.A.; Mir, R.; Abu-Duhier, F. Potential Impact of Vascular Endothelial Growth Factor Gene Variation (-2578C>A) on Breast Cancer Susceptibility in Saudi Arabia: A Case-Control Study. Asian Pac. J. Cancer Prev. APJCP 2018, 19, 1135–1143. [Google Scholar]
- Elfaki, I.; Mir, R.; Duhier, F.M.A.; Alotaibi, M.A.; Alalawy, A.I.; Barnawi, J.; Babakr, A.T.; Mir, M.M.; Altayeb, F.; Mirghani, H.; et al. Clinical Implications of MiR128, Angiotensin I Converting Enzyme and Vascular Endothelial Growth Factor Gene Abnormalities and Their Association with T2D. Curr. Issues Mol. Biol. 2021, 43, 1859–1875. [Google Scholar] [CrossRef]
- Zhao, J.; Li, D.; Tang, H.; Tang, L. Association of vascular endothelial growth factor polymorphisms with polycystic ovarian syndrome risk: A meta-analysis. Reprod. Biol. Endocrinol. 2020, 18, 18. [Google Scholar] [CrossRef]
- Wijaya, A.R.; Surudarma, I.W.; Wihandani, D.M.; Putra, I.W.A.S. Polymorphisms of Vascular Endothelial Growth Factor -2578C/A rs699947 are Risk Factors for Diabetic Retinopathy in Type-2 Diabetes Mellitus Patients in Bali, Indonesia. BioMedicine 2021, 11, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.; Tayeb, F.J.; Barnawi, J.; Jalal, M.M.; Saeedi, N.H.; Hamadi, A.; Altayar, M.A.; Alshammari, S.E.; Mtiraoui, N.; Ali, M.E.; et al. Biochemical Characterization and Molecular Determination of Estrogen Receptor-α (ESR1 PvuII-rs2234693 T>C) and MiRNA-146a (rs2910164 C>G) Polymorphic Gene Variations and Their Association with the Risk of Polycystic Ovary Syndrome. Int. J. Environ. Res. Public Health 2022, 19, 3114. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.; Altayar, M.A.; Hamadi, A.; Tayeb, F.J.; Saeedi, N.H.; Jalal, M.M.; Barnawi, J.; Alshammari, S.E.; Mtiraoui, N.; Ali, M.E.M.; et al. Molecular determination of progesterone receptor’s PROGINS allele (Alu insertion) and its association with the predisposition and susceptibility to polycystic ovary syndrome (PCOS). Mamm Genome 2022, 33, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, A. Effects of opium alkaloid, noscapine in RU486 induced experimental model of polycystic ovary syndrome. Indian J. Biochem. Biophys. 2022, 59, 468–478. [Google Scholar]
- Vural, F.; Vural, B.; Kardaş, E.; Coşkun, A.D.E.; Yildirim, I. The diagnostic performance of antimullerian hormone for polycystic ovarian syndrome and polycystic ovarian morphology. Arch. Gynecol. Obstet. 2023, 307, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Peigné, M.; Pigny, P.; Pankhurst, M.W.; Drumez, E.; Loyens, A.; Dewailly, D.; Catteau-Jonard, S.; Giacobini, P. The proportion of cleaved anti-Mullerian hormone is higher in serum but not follicular fluid of obese women independently of polycystic ovary syndrome. Reprod. Biomed. Online 2020, 41, 1112–1121. [Google Scholar] [CrossRef]
- Mukherjee, S. Pathomechanisms of polycystic ovary syndrome multidimensional approaches. Front. Biosci. 2018, 10, 384–422. [Google Scholar] [CrossRef]
- Kubiszeski, E.H.; de Medeiros, S.F.; da Silva Seidel, J.A.; Barbosa, J.S.; Galera, M.F.; Galera, B.B. Glutathione S-transferase M1 and T1 gene polymorphisms in Brazilian women with endometriosis. J. Assist. Reprod. Genet. 2015, 32, 1531–1535. [Google Scholar] [CrossRef]
- Chrysanthopoulos, I.; Petsavas, A.; Mavrogianni, D.; Potiris, A.; Machairiotis, N.; Drakaki, E.; Vrachnis, D.; Machairoudias, P.; Karampitsakos, T.; Perros, P.; et al. Detection of GSTM1-null Genotype in Women Undergoing IVF Treatment. J. Clin. Med. 2023, 12, 7269. [Google Scholar] [CrossRef]
- Alves, M.M.C.; Almeida, M.; Oliani, A.H.; Breitenfeld, L.; Ramalhinho, A.C. Women with polycystic ovary syndrome and other causes of infertility have a higher prevalence of GSTT1 deletion. Reprod. Biomed. Online 2020, 41, 892. [Google Scholar] [CrossRef] [PubMed]
- Babu, K.A.; Rao, K.L.; Kanakavalli, M.K.; Suryanarayana, V.V.; Deenadayal, M.; Singh, L. CYP1A1, GSTM1 and GSTT1 genetic polymorphism is associated with susceptibility to polycystic ovaries in South Indian women. Reprod. Biomed. Online 2004, 9, 194–200. [Google Scholar] [CrossRef]
- Li, L.; Gu, Z.-P.; Bo, Q.-M.; Wang, D.; Yang, X.-S.; Cai, G.-H. Association of CYP17A1gene -34T/C polymorphism with polycystic ovary syndrome in Han Chinese population. Gynecol. Endocrinol. 2015, 31, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Lizneva, D.; Suturina, L.; Walker, W.; Brakta, S.; Gavrilova-Jordan, L.; Azziz, R. Criteria, prevalence, and phenotypes of polycystic ovary syndrome. Fertil. Steril. 2016, 106, 6–15. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic ovary syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16057. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.R.; Khanna, A.; Singh, K. Association of GSTT1 and GSTM1 polymorphisms with early pregnancy loss in an Indian population and a meta-analysis. Reprod. Biomed. Online. 2013, 26, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Makoui, M.H.; Fekri, S.; Makoui, R.H.; Ansari, N. Individual effects of GSTM1 and GSTT1 polymorphisms on the risk of polycystic ovarian syndrome: A systematic review and meta-analysis. J. Turk. Soc. Obstet. Gynecol. 2023, 20, 314–319. [Google Scholar] [CrossRef]
- Huang, L.; Wang, L. Association between VEGF gene polymorphisms (11 sites) and polycystic ovary syndrome risk. Biosci. Rep. 2020, 40, BSR20191691. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Y.; Fang, L.; Yu, Y.; Shi, H.; Wang, S.; Li, Y.; Ma, Y.; Yan, Y.; Sun, Y.P. Association between vascular endothelial growth factor gene polymorphisms and PCOS risk: A meta-analysis. Reprod. Biomed. Online 2020, 40, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.R.; Gupta, R.; Shukla, M.; Jain, A.; Shukla, D. Association of VEGFA promoter polymorphisms rs699947 and rs35569394 with diabetic retinopathy among North-Central Indian subjects: A case-control study. Ophthalmic Genet. 2022, 43, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Gong, C.; Chen, X.; Zhou, H.; Yan, J.; Hong, W. Associations between Vascular Endothelial Growth Factor Gene Polymorphisms and Different Types of Diabetic Retinopathy Susceptibility: A Systematic Review and Meta-Analysis. J. Diabetes Res. 2021, 2021, 7059139. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cheng, B.; Wu, A.; Zhou, X. Association Between VEGF-460C/T Gene Polymorphism and Risk of Diabetic Retinopathy in Type 2 Diabetes Mellitus: A Meta-Analysis. Horm. Metab. Res. 2024, 56, 214–222. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ARMS-Primers for VEGF-rs699947 C>A | |||||
|---|---|---|---|---|---|
| Gene | Allele | Sequence | PCR Product | Annealing Temperature | References |
| VEGF Fo | 5-CCTTTTCCTCATAAGGGCCTTAG-3 | 353 bp | 58.1 °C | [20,30] | |
| VEGF Ro | 5-AGGA AGC AGCT TGGAA AAA TTC-3 | ||||
| VEGF FI | A allele | 5-TAGG CCA GAC CCTG GCAA-3 | 149bp | ||
| VEGF RI | C allele | 5-GTC TGA TTA TCCA CCC AGAT CG-3 | 243bp | ||
| MS-Primers for VEGF rs4646994 I/D | |||||
| VEGF F | D-allele | 5′-GCTGAGAGTGGGGCTGACTAGGTA-3′ | 211 bp | 58.8 °C | [31] |
| VEGF R | I-allele | 5′-GTTTCTGACCTGGCTATTTCCAGG-3′ | 229 bp | ||
| GSTT1 and GSTM1 (rs4025935 and rs71748309) Multiplex PCR Primers for Genotyping | |||||
| GSTT1 F | GSTT1 | 5′-TTCCTTACTGGTCCTCACATCTC-3′ | 480 bp | 60 °C | [14] |
| GSTT1 R | 5′-TCA CGG GAT CAT GGC CAG CA-3′ | ||||
| GSTM1 F | GSTM1 | 5′-GAA CTC CCT GAA AAG CTA AAG C-3′ | 215 bp | ||
| GSTM1 R | 5′-GTT GGG CTC AAA TAT ACG GTG G-3′ | ||||
| CYP1A1F | CYP1A1 | 5′-GAA CTG CCA CTT CAG CTG TCT-3′ | 312 bp | ||
| CYP1A1R | 5′-CA G CTG C AT T T GG AA G T G C TC-3′ | ||||
| Characteristic | Controls J | Cases J | p K |
|---|---|---|---|
| FBG (mmol/L) G | 4.60 ± 0.66 | 7.60 ± 5.16 | <0.0020 |
| Age Y | 28.55 ± 4.99 | 29.20 ± 6.10 | 0.420 |
| BMI (kg/m2) G | 29.10 ± 4.05 | 27.11 ± 5.90 | <0.004 |
| HDL (mmol/L) G | 1.73 ± 0.89 | 1.78 ± 0.91 | <0.0032 |
| LDL (mmol/L) G | 2.98 ± 0.70 | 4.98 ± 1.90 | <0.0024 |
| Cholesterol (mmol/L) G | 3.80 ± 0.60 | 5.23 ± 0.81 | <0.0023 |
| Triglycerides (mmol/L) G | 1.78 ± 0.65 | 3.58 ± 1.41 | <0.0471 |
| Luteinizing hormone (mIU/mL) M | 0.09 (0.09–1.90) | 3.65(0.75–9.10) | <0.0040 |
| Testosterone (ng/dL) M | 14.90 (6.80–13.55) | 60.15 (46.80–70.30) | <0.0040 |
| Estradiol (pmol/L) M | 334.70 ± 249.49 | 386.66 ± 336.79 | 0.169 |
| Progesterone (ng/mL) M | 15.55 (3.10–18.10) | 18.30 (2.40–33.10) | <0.0040 |
| HOMA-IR G | 4.00 ± 0.82 | 6.00 ± 5.88 | <0.0030 |
| Free Insulin (mU/mL) G | 7.90 ± 2.99 | 14.39 ± 5.97 | <0.0021 |
| FSH (mIU/mL) M | 0.89 (0.11–5.21) | 4.10 (3.15–6.11) | <0.0040 |
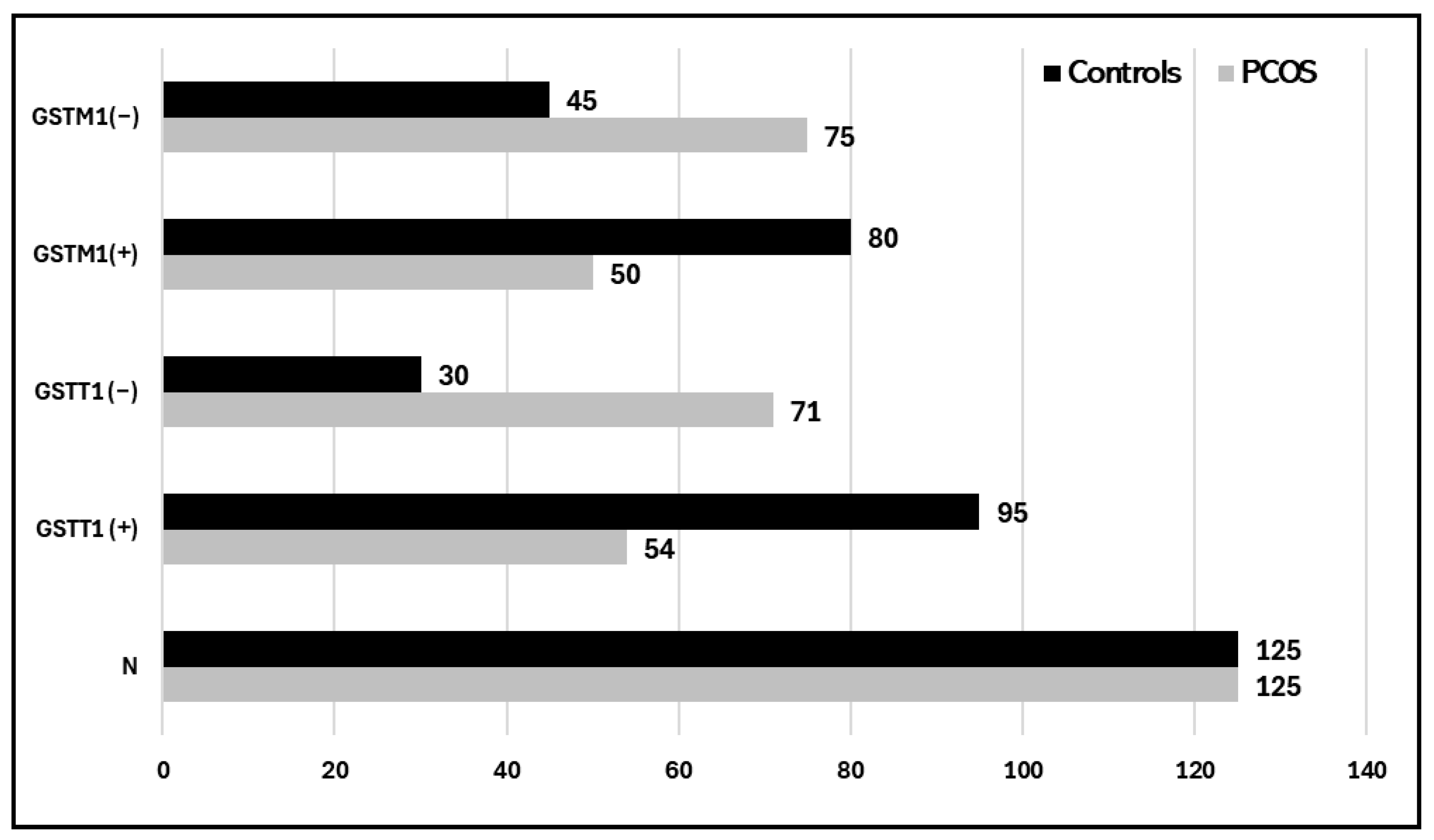
| Variables | Controls (125) | PCOS (125) | |||||
|---|---|---|---|---|---|---|---|
| GSTT1 genotype frequency in case and control groups | |||||||
| GSTT1 (+) | 95 (76%) | 54 (43.2%) | |||||
| GSTT1 (−) | 30 (24%) | 71 (56.8%) | |||||
| Genotype frequencies of GSTM1 in case and control groups | |||||||
| GSTM1(+) | 80 (64%) | 50 (40%) | |||||
| GSTM1(−) | 45 (36%) | 75 (60%) | |||||
| Genotype frequencies of GSTT1/GSTM1 in case and control groups | |||||||
| GSTT1 (−) GSTM1 (−) | 42 (33.6%) | 25 (20%) | |||||
| GSTT1 (+) GSTM1 (+) | 33 (26.4%) | 31 (24.8%) | |||||
| GSTT1 (+) GSTM1 (−) | 30 (24%) | 54 (43.2%) | |||||
| GSTT1 (−) GSTM1 (+) | 20 (16%) | 15 (12%) | |||||
| Association of GSTM1/GSTT1 null genotypes with PCOS risk | |||||||
| Association of GSTT1 (+) and GSTT1 (−) genotypes with PCOS risk | |||||||
| Variables | N = 125 | N = 125 | OR (95% CI) | RR (95% CI) | p value | ||
| GSTT1 (+) | 95(76%) | 54 (43.2%) | Ref. 1.00 | Ref. 1.00 | |||
| GSTT1 (−) | 30 (24%) | 71 (56.8%) | 4.16 (2.4215 to 7.1589) | 2.14 (1.5532 to 2.9665) | 0.0001 | ||
| Association of GSTM1 (+) and GSTM1 (−) genotypes with PCOS risk | |||||||
| Variables | N = 125 | N = 125 | OR (95% CI) | RR (95% CI) | |||
| GSTM1 (+) | 80 (64%) | 50 (40%) | Ref. 1.00 | Ref. 1.00 | |||
| GSTM1 (−) | 45 (36%) | 75 (60%) | 2.66 (1.5992 to 4.4466) | 1.64 (1.2552 to 2.1454) | 0.0002 | ||
| Association of GSTT1 (+) GSTM1 (+) and GSTT1 (−) GSTM1 (−) genotypes with PCOS risk | |||||||
| Variables | N = 125 | N = 125 | OR (95% CI) | RR (95% CI) | p value | ||
| GSTT1 (−) GSTM1 (−) | 42 (33.6%) | 25 (20%) | Ref. 1.00 | Ref. 1.00 | |||
| GSTT1 (+) GSTM1 (+) | 33 (26.4%) | 31 (24.8%) | 1.57 (0.7863 to 3.1678) | 1.08 (0.7959 to 1.4820) | 0.19 | ||
| Association of GSTT1 (+) GSTM1 (+) and GSTT1 (−) GSTM1 (−) genotypes with PCOS risk | |||||||
| Variables | N = 125 | N = 125 | OR (95% CI) | RR (95% CI) | p value | ||
| GSTT1 (+) GSTM1 (+) | 33 (21.81%) | 31 (38.18%) | Ref. 1.00 | Ref. 1.00 | |||
| GSTT1 (−) GSTM1 (−) | 42 (29.09%) | 25(30%) | 0.63 (0.3157 to 1.2719) | 0.82 (0.6088 to 1.1113) | 0.199 | ||
| Association of GSTT1 (+) GSTM1 (−) and GSTT1 (−) GSTM1 (−) genotypes with PCOS risk | |||||||
| Variables | N = 125 | N = 125 | OR (95% CI) | RR (95% CI) | p value | ||
| GSTT1 (+) GSTM1 (−) | 30 (24%) | 54 (43.2%) | Ref. 1.00 | Ref. 1.00 | |||
| GSTT1 (−) GSTM1 (+) | 20 (16%) | 15 (12%) | 0.41 (0.1864 to 0.9316) | 0.62 (0.4165 to 0.9378) | 0.003 | ||
| Subjects | N= | I | ID | D | Df | X2 | I | D | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Cases | 125 | 32(25.5%) | 50(40%) | 43(32%) | 2 | 11.38 | 0.58 | 0.52 | 0.0034 |
| Controls | 125 | 38(30.4%) | 67(53.6%) | 20(16%) | 0.65 | 0.35 | |||
| Association of VEGF-rs699947 C>A gene variation in PCOS cases and controls | |||||||||
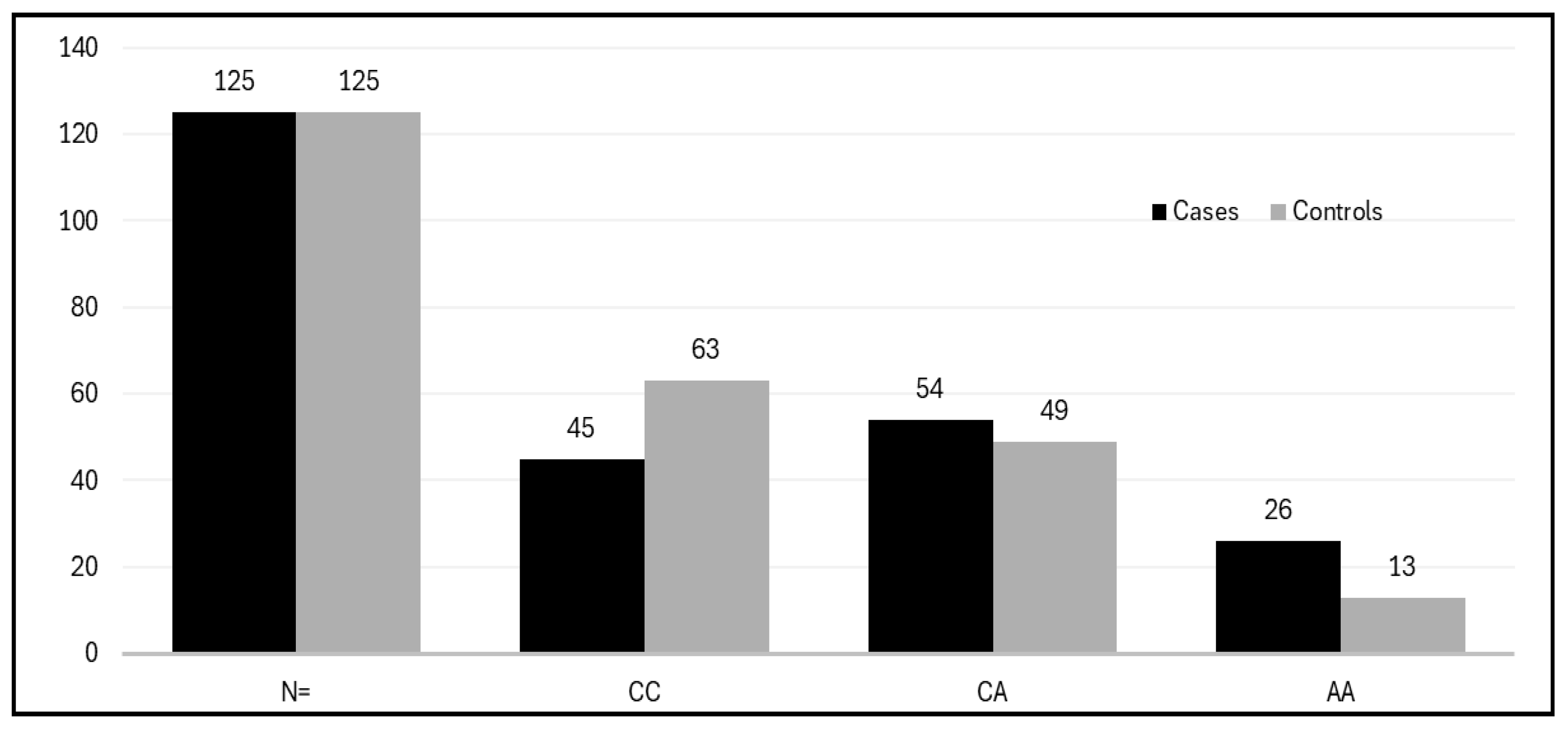
| Subjects | N= | CC | CA | AA | Df | X2 | C | A | p value |
| Cases | 125 | 45(36%) | 54(43.2%) | 26(20.8%) | 2 | 7.58 | 0.58 | 0.42 | 0.020 |
| Controls | 125 | 63(50.4%) | 49(39.2%) | 13(10.4%) | 0.70 | 0.30 | |||
| Genotypes | Controls | PCOS Cases | OR (95% CI) | Risk Ratio (RR) | p-Value |
|---|---|---|---|---|---|
| (N = 125) | (N = 125) | ||||
| Codominant inheritance model | |||||
| VEGF-(II) | 38 | 32 | 1 (ref.) | 1 (ref.) | |
| VEGF-(ID) | 67 | 50 | 0.88 (0.4883 to 1.6084) | 0.94 (0.7266 to 1.2368) | 0.69 |
| VEGF-(DD) | 20 | 43 | 2.55 (1.2566 to 5.1874) | 1.71 (1.1223 to 2.6054) | 0.009 |
| Dominant inheritance model | |||||
| VEGF-(II) | 38 | 32 | 1 (ref.) | 1 (ref.) | |
| VEGF-(ID+DD) | 87 | 93 | 1.26 (0.7297 to 2.2084) | 1.12 (0.8636 to 1.4606) | 0.390 |
| Recessive inheritance model | |||||
| VEGF-(I+ID) | 105 | 82 | 1 (ref.) | 1 (ref.) | |
| VEGF-(D) | 20 | 43 | 2.75 (1.5048 to 5.0367) | 1.76 (1.2052 to 2.5957) | 0.001 |
| Additive inheritance model (Allele) | |||||
| VEGF-(I) | 143 | 112 | 1 (ref.) | 1 (ref.) | |
| VEGF-(D) | 107 | 136 | 1.62 (1.1390 to 2.3121) | 1.27 (1.0653 to 1.5226) | 0.007 |
| Genotypes | Healthy Controls | PCOS Cases | OR (95% CI) | Risk Ratio (RR) | p-Value |
|---|---|---|---|---|---|
| (n = 125) | (n = 125) | ||||
| Codominant inheritance model | |||||
| VEGF-(CC) | 63 | 45 | 1 (ref.) | 1 (ref.) | |
| VEGF-(CA) | 49 | 54 | 1.54 (0.8955 to 2.6580) | 1.22 (0.947 to 1.586) | 0.11 |
| VEGF-(AA) | 13 | 26 | 2.80 (1.2992 to 6.0344) | 1.75 (1.092 to 2.804) | 0.008 * |
| Dominant inheritance model | |||||
| VEGF-(CC) | 63 | 45 | 1 (ref.) | 1 (ref.) | |
| VEGF-(CA + AA) | 62 | 80 | 1.80 (1.0888 to 2.9971) | 1.33 (1.0451 to 1.7079) | 0.022 * |
| Recessive inheritance model | |||||
| VEGF-(CC + CA) | 112 | 99 | 1 (ref.) | 1 (ref.) | |
| VEGF-(AA) | 13 | 26 | 2.26 (1.1029 to 4.6418) | 1.59 (1.0036 to 2.5266) | 0.025 * |
| Additive inheritance model (Allele) | |||||
| VEGF-(C) | 175 | 144 | 1 (ref.) | 1 (ref.) | |
| VEGF-(A) | 75 | 106 | 1.71 (1.1877 to 2.483) | 1.32 (1.0842 to 1.6167) | 0.004 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almotairi, R.; Mir, R.; Almasoudi, K.S.; Husain, E.; Mtiraoui, N. Evaluating the Impact of rs4025935, rs71748309, rs699947, and rs4646994 Genetic Determinants on Polycystic Ovary Syndrome Predisposition—A Case-Control Study. Life 2025, 15, 558. https://doi.org/10.3390/life15040558
Almotairi R, Mir R, Almasoudi KS, Husain E, Mtiraoui N. Evaluating the Impact of rs4025935, rs71748309, rs699947, and rs4646994 Genetic Determinants on Polycystic Ovary Syndrome Predisposition—A Case-Control Study. Life. 2025; 15(4):558. https://doi.org/10.3390/life15040558
Chicago/Turabian StyleAlmotairi, Reema, Rashid Mir, Kholoud S. Almasoudi, Eram Husain, and Nabil Mtiraoui. 2025. "Evaluating the Impact of rs4025935, rs71748309, rs699947, and rs4646994 Genetic Determinants on Polycystic Ovary Syndrome Predisposition—A Case-Control Study" Life 15, no. 4: 558. https://doi.org/10.3390/life15040558
APA StyleAlmotairi, R., Mir, R., Almasoudi, K. S., Husain, E., & Mtiraoui, N. (2025). Evaluating the Impact of rs4025935, rs71748309, rs699947, and rs4646994 Genetic Determinants on Polycystic Ovary Syndrome Predisposition—A Case-Control Study. Life, 15(4), 558. https://doi.org/10.3390/life15040558