

Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview

Abstract
1. Introduction
2. Abiotic Stresses in the Cereal Growth Environment
3. Responses of Cereal Plants to Saline Stress
4. Improvement of Crop Plants under Saline Stress
5. Responses of Wheat Plants Infected with Z. tritici
5.1. Symptoms and Life Cycle of Z. tritici
5.2. Mechanisms of Disease Progression of Z. tritici
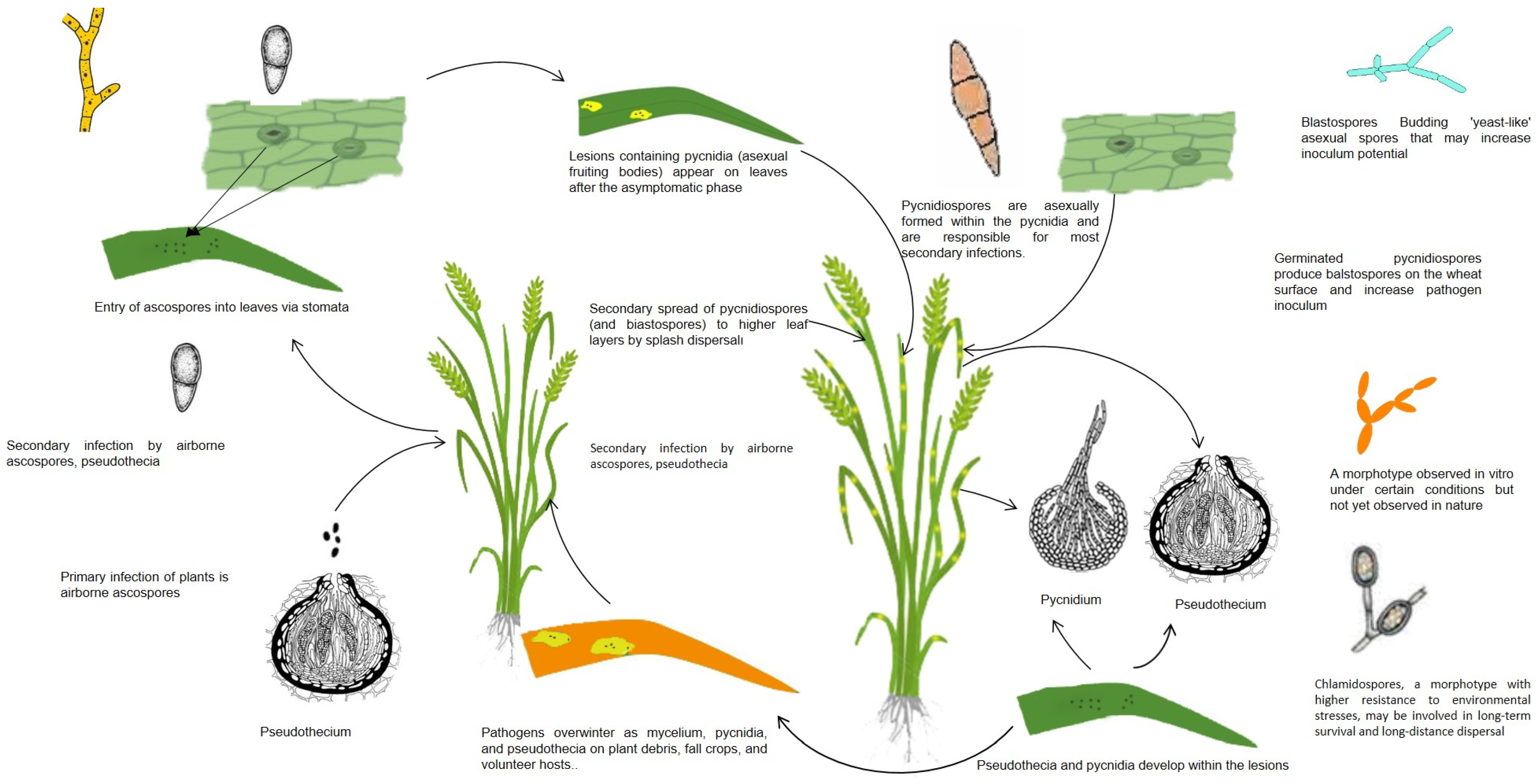
5.3. Role of Toxins and Pathogenic Enzymes of Z. tritici
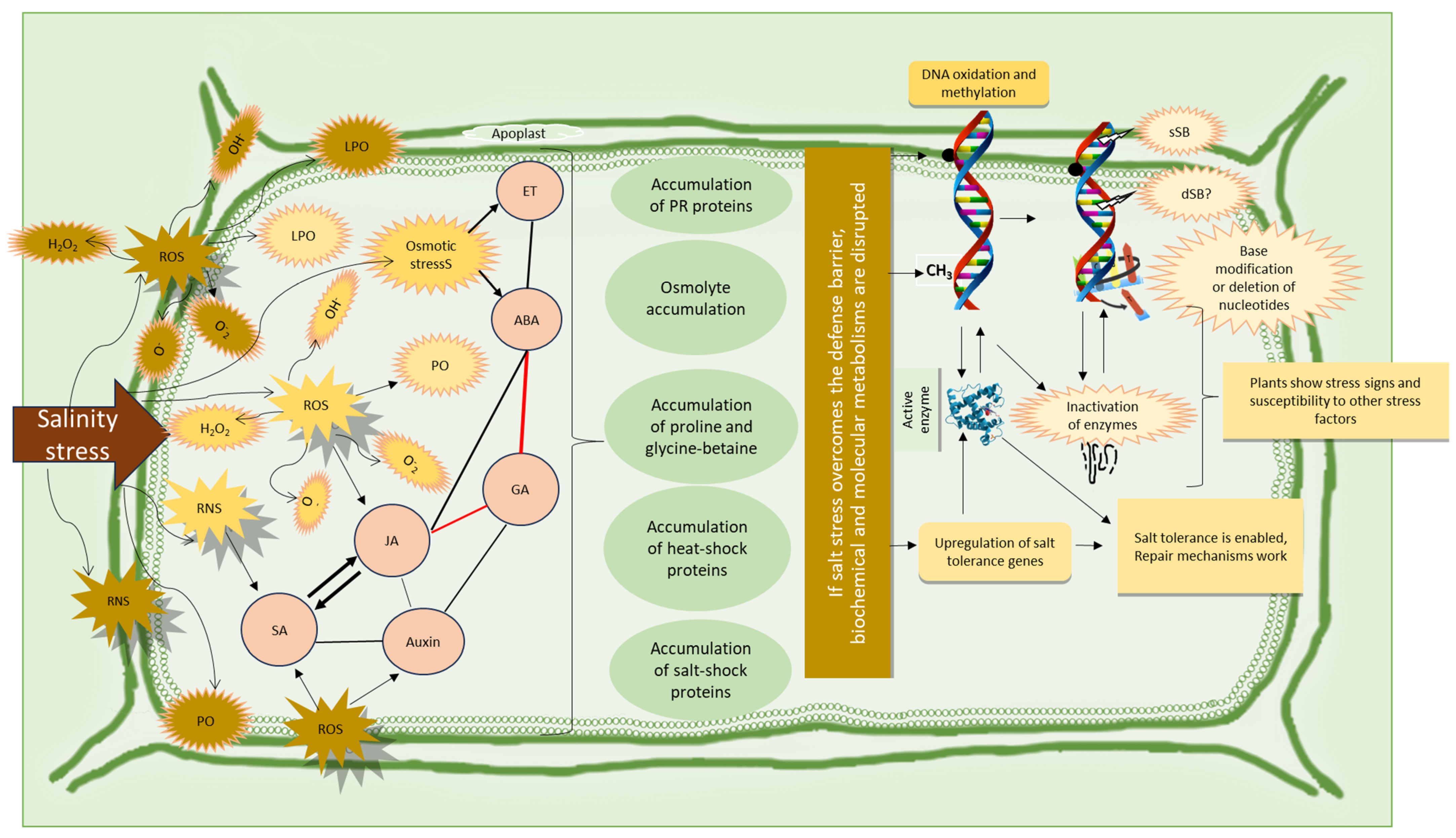
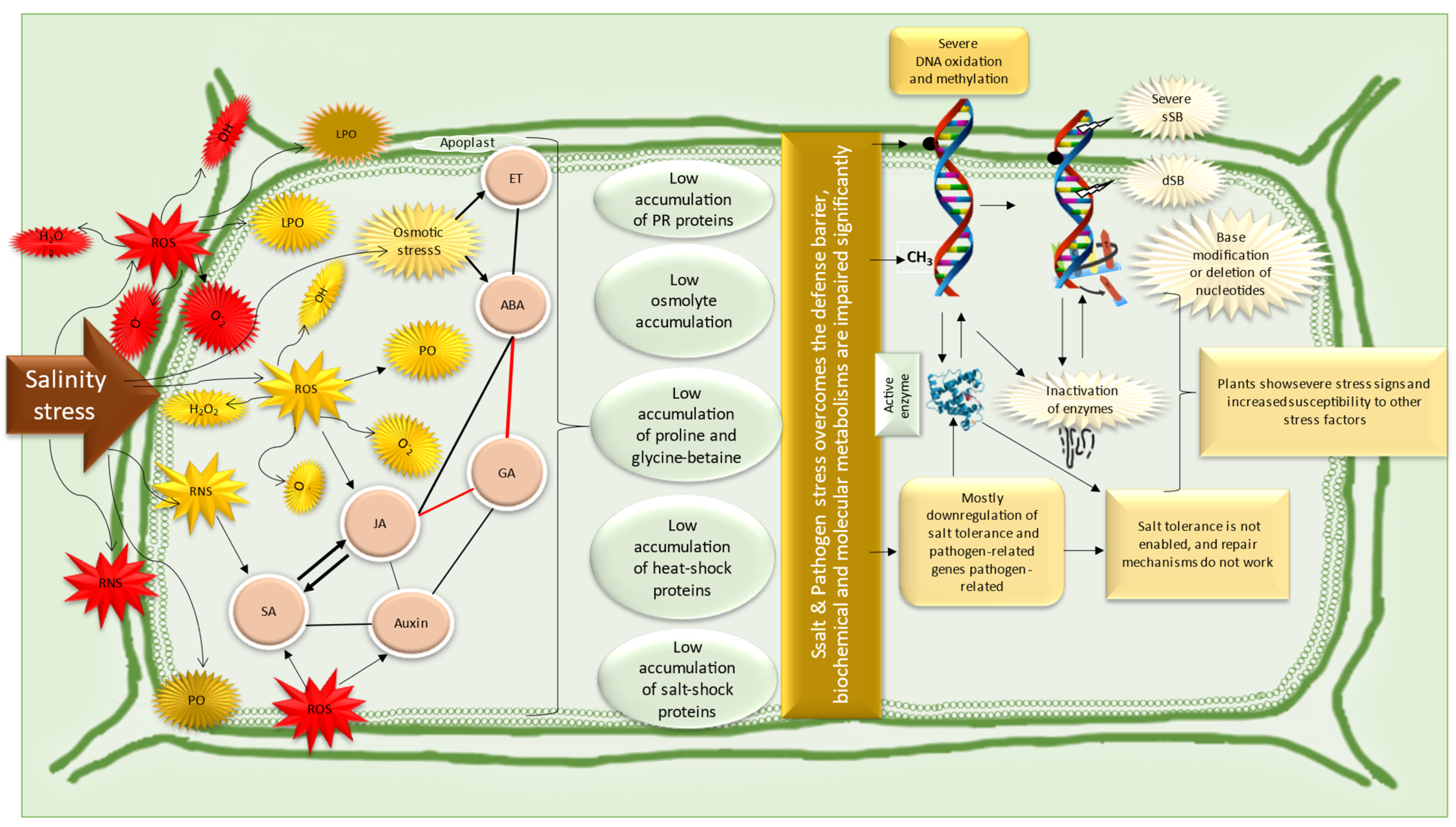
6. Responses of Cereal Plants and Pathogens to Salinity Stress
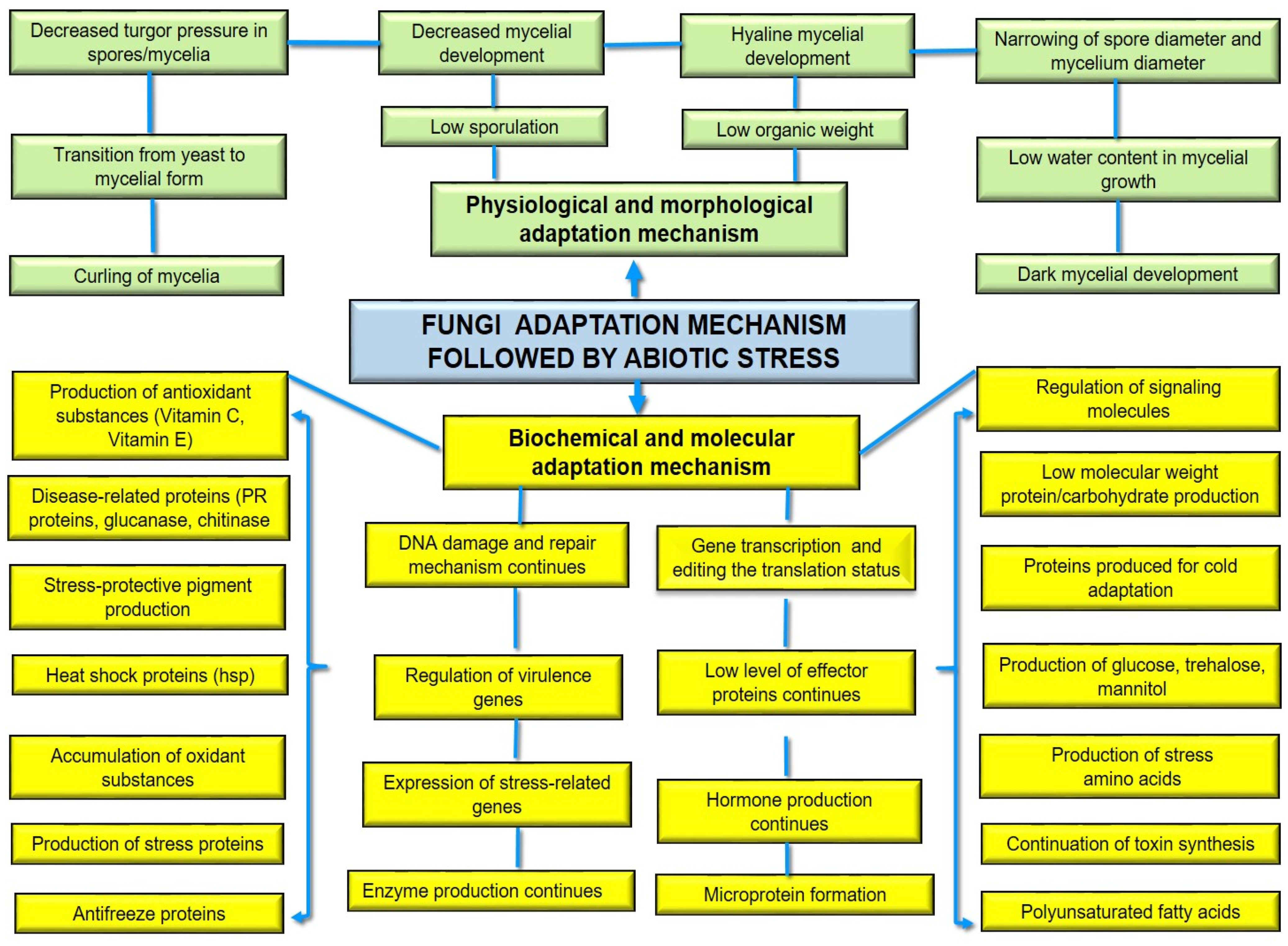
6.1. Adaptation of Fungi to Stress Conditions
6.2. Responses of Cereal Plants to Saline and Pathogenic Stress Conditions
7. Mitigation Strategies for Salinity-Induced Pathogenicity in Cereal Plants Exposed to Saline and Pathogen Stressors
8. Future Directions and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roubtsova, T.V.; Bostock, R.M. Episodic abiotic stress as a potential contributing factor to onset and severity of disease caused by Phytophthora ramorum in Rhododendron and Viburnum. Plant Dis. 2009, 93, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, P.; Achary, V.M.M.; Panda, B.B. Aluminum-induced abiotic stress counteracts Fusarium infection in Cajanus cajan (L.) Millsp. J. Plant Interact. 2012, 7, 121–128. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.; Feldman, T.; Sumner, L.; Roossinck, M. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Ramegowda, V.; Senthil, A.; Senthil-Kumar, M. Stress combinations and their interactions in crop plants. Plant Physiol. Rep. 2024, 29, 1–5. [Google Scholar] [CrossRef]
- Dikilitas, M.; Karakas, S. Behaviour of plant pathogens for crops under stress during the determination of physiological, biochemical, and molecular approaches for salt stress tolerance. In Crop Production for Agricultural Improvement; Springer: Berlin/Heidelberg, Germany, 2012; pp. 417–441. [Google Scholar]
- Dikilitas, M.; Oğuz, A.Ç.; Karakaya, A. Extracellular protease activity and glucose production in isolates of net blotch pathogens differing in virulence. Zemdirb.-Agric. 2018, 105, 89–94. [Google Scholar] [CrossRef]
- Dikilitas, M. Effect of Salinity and Its Interactions with Verticillium albo-atrum on the Disease Development in Tomato (Lycopersicon esculentum Mill) and Lucerne (Medicago sativa L. & M. media) Plants; Swansea University: Swansea, UK, 2003; ISBN 0-355-88288-4. [Google Scholar]
- Liu, Y.; Gong, X.; Li, M.; Si, H.; Zhou, Q.; Liu, X.; Fan, Y.; Zhang, X.; Han, J.; Gu, S. Effect of Osmotic Stress on the Growth, Development and Pathogenicity of Setosphaeria turcica. Front. Microbiol. 2021, 12, 706349. [Google Scholar] [CrossRef]
- Ghiasi Noei, F.; Imami, M.; Didaran, F.; Ghanbari, M.A.; Zamani, E.; Ebrahimi, A.; Aliniaeifard, S.; Farzaneh, M.; Javan-Nikkhah, M.; Feechan, A. Stb6 mediates stomatal immunity, photosynthetic functionality, and the antioxidant system during the Zymoseptoria tritici-wheat interaction. Front. Plant Sci. 2022, 13, 1004691. [Google Scholar] [CrossRef]
- 2015–2016 Yılı Buğday Değerlendirmesi—Ulusal Hububat Konseyi. Available online: http://www.uhk.org.tr/tr/2015-2016-yili-bugday-degerlendirmesi/ (accessed on 19 February 2024).
- Khan, A.Q.; Robe, B.L.; Girma, A. Evaluation of wheat genotypes (Triticum aestivum L.) for yield and yield characteristics under low land area at Arba Minch, Southern Ethiopia. Afr. J. Plant Sci. 2020, 14, 461–469. [Google Scholar]
- Gu, D.; Andreev, K.; Dupre, M.E. Major trends in population growth around the world. China CDC Wkly. 2021, 3, 604. [Google Scholar] [CrossRef] [PubMed]
- Türkiye İstatistik Kurumu (TÜİK). Available online: https://www.tuik.gov.tr/ (accessed on 19 February 2024).
- Jia, J.; Bai, J.; Gao, H.; Wang, W.; Yin, S.; Wang, D.; Han, L. Effects of salinity and moisture on sediment net nitrogen mineralization in salt marshes of a Chinese estuary. Chemosphere 2019, 228, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Arneth, A.; Shin, Y.-J.; Leadley, P.; Rondinini, C.; Bukvareva, E.; Kolb, M.; Midgley, G.F.; Oberdorff, T.; Palomo, I.; Saito, O. Post-2020 biodiversity targets need to embrace climate change. Proc. Natl. Acad. Sci. USA 2020, 117, 30882–30891. [Google Scholar] [CrossRef] [PubMed]
- Luck, J.; Spackman, M.; Freeman, A.; Trębicki, P.; Griffiths, W.; Finlay, K.; Chakraborty, S. Climate change and diseases of food crops. Plant Pathol. 2011, 60, 113–121. [Google Scholar] [CrossRef]
- Nicol, J.; Turner, S.; Coyne, D.L.; den Nijs, L.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Berlin/Heidelberg, Germany, 2011; pp. 21–43. [Google Scholar]
- Khan, R.W.A.; Awan, F.S.; Iqbal, R.K. Evaluation and identification of salt-tolerant wheat through in vitro salinity induction in seeds. Pak J. Bot. 2022, 54, 1987–1993. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar] [CrossRef]
- Nelson, T.; Hose, G.C.; Dabovic, J.; Korbel, K.L. Salinity as a major influence on groundwater microbial communities in agricultural landscapes. Mar. Freshw. Res. 2024, 75, MF23014. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.H.; Singh, R.K. Smart reprograming of plants against salinity stress using modern biotechnological tools. Crit. Rev. Biotechnol. 2022, 43, 1035–1062. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M. Salt Stress Tolerance in Rice and Wheat: Physiological and Molecular Mechanism; Intech Open: Rijeka, Croatia, 2022. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Singh, D.; Thapa, S.; Singh, J.P.; Mahawar, H.; Saxena, A.K.; Singh, S.K.; Mahla, H.R.; Choudhary, M.; Parihar, M.; Choudhary, K.B. Prospecting the Potential of Plant Growth-Promoting Microorganisms for Mitigating Drought Stress in Crop Plants. Curr. Microbiol. 2024, 81, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Damaris, R.N.; Tang, F.; Fan, X.; Fang, Z.; Yang, P.; Li, M. Salt induced protein dynamics in young rice roots of osmybcc-1 mutant and its involvement in salt stress. Plant Stress 2024, 11, 100385. [Google Scholar] [CrossRef]
- Lee, J.H.; Kasote, D.M. Nano-Priming for Inducing Salinity Tolerance, Disease Resistance, Yield Attributes, and Alleviating Heavy Metal Toxicity in Plants. Plants 2024, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Peng, X.; Yang, Z.; Zhao, W.; Xu, W.; Hao, J.; Wu, W.; Shen, X.L.; Luo, Y.; Huang, K. iTRAQ mitoproteome analysis reveals mechanisms of programmed cell death in Arabidopsis thaliana induced by ochratoxin A. Toxins 2017, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Vivancos, P.; Barba-Espín, G.; Hernández, J.A. Elucidating hormonal/ROS networks during seed germination: Insights and perspectives. Plant Cell Rep. 2013, 32, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xiao, C.; Qiu, T.; Deng, J.; Cheng, H.; Cong, X.; Cheng, S.; Rao, S.; Zhang, Y. Selenium regulates antioxidant, photosynthesis, and cell permeability in plants under various abiotic stresses: A review. Plants 2022, 12, 44. [Google Scholar] [CrossRef]
- Han, J.; Dai, J.; Chen, Z.; Li, W.; Li, X.; Zhang, L.; Yao, A.; Zhang, B.; Han, D. Overexpression of a ‘Beta’MYB Factor Gene, VhMYB15, Increases Salinity and Drought Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2024, 25, 1534. [Google Scholar] [CrossRef] [PubMed]
- Asghar, N.; Akram, N.A.; Ameer, A.; Shahid, H.; Kausar, S.; Asghar, A.; Idrees, T.; Mumtaz, S.; Asfahan, H.M.; Sultan, M. Foliar-applied hydrogen peroxide and proline modulates growth, yield and biochemical attributes of wheat (Triticum aestivum L.) Under varied n and p levels. Fresenenius Environ. Bull. 2021, 30, 5445–5465. [Google Scholar]
- Malik, S.; Ashraf, M.; Arshad, M.; Malik, T.A. Effect of ascorbic acid application on physiology of wheat under drought stress. Pak. J. Agric. Sci. 2015, 52, 209–217. [Google Scholar]
- Parveen, K.; Saddique, M.A.B.; Ali, Z.; Rehman, S.U.; Khan, Z.; Waqas, M.; Munir, M.Z.; Hussain, N.; Muneer, M.A. Genome-wide analysis of Glutathione peroxidase (GPX) gene family in Chickpea (Cicer arietinum L.) under salinity stress. Gene 2024, 898, 148088. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Berk, K.; Aronsson, H. Evolution and identification of DREB transcription factors in the wheat genome: Modeling, docking and simulation of DREB proteins associated with salt stress. J. Biomol. Struct. Dyn. 2022, 40, 7191–7204. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, L.; Demir, Y. In vivo and in vitro protective role of proline. Plant Growth Regul. 2002, 38, 259–264. [Google Scholar] [CrossRef]
- Mandhania, S.; Madan, S.; Sawhney, V. Antioxidant defense mechanism under salt stress in wheat seedlings. Biol. Plant. 2006, 50, 227–231. [Google Scholar] [CrossRef]
- Hu, L.; Li, H.; Pang, H.; Fu, J. Responses of antioxidant gene, protein and enzymes to salinity stress in two genotypes of perennial ryegrass (Lolium perenne) differing in salt tolerance. J. Plant Physiol. 2012, 169, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Maršálová, L.; Vítámvás, P.; Hynek, R.; Prášil, I.T.; Kosová, K. Proteomic response of Hordeum vulgare cv. Tadmor and Hordeum marinum to salinity stress: Similarities and differences between a glycophyte and a halophyte. Front. Plant Sci. 2016, 7, 1154. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, B.; Millar, H.; Taylor, N.; Staudinger, C.; Munns, R. Proteomic Responses of Wheat Root Tissues to Salt Stress in Relation to Root Physiology. In Proceedings of the Plant Biology Worldwide Summit 2020, Pittsburgh, PA, USA, 27–31 July 2020. [Google Scholar] [CrossRef]
- Sofy, M.R.; Sharaf, A.; Osman, M.S.; Sofy, A.R. Physiological changes, antioxidant activity, lipid peroxidation and yield characters of salt-stressed barely plant in response to treatment with Sargassum extract. Int. J. Adv. Res. Biol. Sci. 2017, 4, 90–109. [Google Scholar]
- Amirbakhtiar, N.; Ismaili, A.; Ghaffari, M.-R.; Mirdar Mansuri, R.; Sanjari, S.; Shobbar, Z.-S. Transcriptome analysis of bread wheat leaves in response to salt stress. PLoS ONE 2021, 16, e0254189. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Han, Z.; Yin, H.; Chen, F.; Dong, Y.; Zhang, L.; Lu, X.; Zeng, J.; Ma, W.; Mu, P. High-throughput sequencing-based identification of miRNAs and their target mRNAs in wheat variety Qing Mai 6 under salt stress condition. Front. Genet. 2021, 12, 724527. [Google Scholar] [CrossRef] [PubMed]
- Nassar, R.M.; Kamel, H.A.; Ghoniem, A.E.; Alarcón, J.J.; Sekara, A.; Ulrichs, C.; Abdelhamid, M.T. Physiological and anatomical mechanisms in wheat to cope with salt stress induced by seawater. Plants 2020, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tian, L.; Zhu, Z.; Sang, Z.; Ma, L.; Jia, Z. Growth and Physiological Responses of Magnoliaceae to NaCl Stress. Plants 2024, 13, 170. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance—Current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Cabot, C.; Bosch, R.; Martos, S.; Poschenrieder, C.; Perelló, A. Salinity is a prevailing factor for amelioration of wheat blast by biocontrol agents. Biol. Control 2018, 125, 81–89. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Afzal, I.; Ibrahim, A.M.; Bakhtavar, M.A.; Hafeez, M.B.; Jahanzaib; Maqbool, M.M. Mitigation of salinity stress in wheat (Triticum aestivum L.) seedlings through physiological seed enhancements. J. Plant Nutr. 2019, 42, 1192–1204. [Google Scholar] [CrossRef]
- Karakaya, A.; Dikilitas, M. Biochemical, physiological and molecular defence mechanisms of tea plants against pathogenic agents under changing climate conditions. In Stress Physiology of Tea in the Face of Climate Change; Springer: Singapore, 2018; pp. 241–268. [Google Scholar] [CrossRef]
- El Sabagh, A.; Islam, M.S.; Skalicky, M.; Ali Raza, M.; Singh, K.; Anwar Hossain, M.; Hossain, A.; Mahboob, W.; Iqbal, M.A.; Ratnasekera, D. Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and management strategies. Front. Agron. 2021, 3, 661932. [Google Scholar] [CrossRef]
- Fan, H.; Du, C.; Xu, Y.; Wu, X. Exogenous nitric oxide improves chilling tolerance of Chinese cabbage seedlings by affecting antioxidant enzymes in leaves. Hortic. Environ. Biotechnol. 2014, 55, 159–165. [Google Scholar] [CrossRef]
- Ma, N.; Hu, C.; Wan, L.; Hu, Q.; Xiong, J.; Zhang, C. Strigolactones improve plant growth, photosynthesis, and alleviate oxidative stress under salinity in rapeseed (Brassica napus L.) by regulating gene expression. Front. Plant Sci. 2017, 8, 1671. [Google Scholar] [CrossRef] [PubMed]
- Kausar, F.; Shahbaz, M.; Ashraf, M. Protective role of foliar-applied nitric oxide in Triticum aestivum under saline stress. Turk. J. Bot. 2013, 37, 1155–1165. [Google Scholar] [CrossRef]
- Abu-Qaoud, H.; Al-Fares, H.; Shtaya, M.J.; Shawarb, N. Effect of effective microorganisms on wheat growth under salt stress condition. Chil. J. Agric. Res. 2021, 81, 351–356. [Google Scholar] [CrossRef]
- Meena, K.K.; Bitla, U.M.; Sorty, A.M.; Singh, D.P.; Gupta, V.K.; Wakchaure, G.; Kumar, S. Mitigation of salinity stress in wheat seedlings due to the application of phytohormone-rich culture filtrate extract of methylotrophic actinobacterium Nocardioides sp. NIMMe6. Front. Microbiol. 2020, 11, 2091. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, O.; Abouseadaa, H.; Abdelmoneim, T.K.; Alshehri, M.A.; Mohamed, E.-M.; El-Beltagi, H.S.; Mohamed, A. Agronomical, physiological and molecular evaluation reveals superior salt-tolerance in bread wheat through salt-induced priming approach. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12310. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Fujita, M. Pretreatment of wheat (Triticum aestivum L.) seedlings with 2, 4-D improves tolerance to salinity-induced oxidative stress and methylglyoxal toxicity by modulating ion homeostasis, antioxidant defenses, and glyoxalase systems. Plant Physiol. Biochem. 2020, 152, 221–231. [Google Scholar] [CrossRef]
- Ekim, R.; Arikan, B.; Alp-Turgut, F.N.; Koyukan, B.; Ozfidan-Konakci, C.; Yildiztugay, E. Polyvinylpyrrolidone-coated copper nanoparticles dose-dependently conferred tolerance to wheat under salinity and/or drought stress by improving photochemical activity and antioxidant system. Environ. Res. 2024, 241, 117681. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Rizwan, M.; Huizhi, W.; Ulhassan, Z.; Xie, W. Silicon nanoparticles improved the osmolyte production, antioxidant defense system, and phytohormone regulation in Elymus sibiricus (L.) under drought and salt stress. Environ. Sci. Pollut. Res. 2024, 31, 8985–8999. [Google Scholar] [CrossRef] [PubMed]
- Maslennikova, D.; Ivanov, S.; Petrova, S.; Burkhanova, G.; Maksimov, I.; Lastochkina, O. Components of the phenylpropanoid pathway in the implementation of the protective effect of sodium nitroprusside on wheat under salinity. Plants 2023, 12, 2123. [Google Scholar] [CrossRef]
- Jamshidi, B.; Pour-Aboughadareh, A.; Bocianowski, J.; Shooshtari, L.; Bujak, H.; Türkoğlu, A.; Nowosad, K. Analysis of Physio-Biochemical Responses and Expressional Profiling Antioxidant-Related Genes in Some Neglected Aegilops Species under Salinity Stress. Agronomy 2023, 13, 1981. [Google Scholar] [CrossRef]
- Paul, A.; Mondal, S.; Pal, A.; Biswas, S.; Chakraborty, K.; Mazumder, A.; Biswas, A.K.; Kundu, R. Seed priming with NaCl helps to improve tissue tolerance, potassium retention ability of plants, and protects the photosynthetic ability in two different legumes, chickpea and lentil, under salt stress. Planta 2023, 257, 111. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bai, X.; Qiao, Y.; Si, L.; Yu, Z.; Ni, C.; Li, T.; Guo, C.; Xiao, K. tae-miR9674a, a microRNA member of wheat, confers plant drought and salt tolerance through modulating the stomata movement and ROS homeostasis. Plant Biotechnol. Rep. 2023, 17, 471–488. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Y.; Sang, S.; Qiu, Z. Jasmonic acid pretreatment improves salt tolerance of wheat by regulating hormones biosynthesis and antioxidant capacity. Front. Plant Sci. 2022, 13, 968477. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dong, S.; Teng, K.; Chang, Z.; Zhang, X. Exogenous salicylic acid optimizes photosynthesis, antioxidant metabolism, and gene expression in Perennial Ryegrass subjected to salt stress. Agronomy 2022, 12, 1920. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki Ourang, S.; Khalili, P.; Poczai, P. Unraveling salinity stress responses in ancestral and neglected wheat species at early growth stage: A baseline for utilization in future wheat improvement programs. Physiol. Mol. Biol. Plants 2020, 26, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, S.; Feki, K.; Hmidi, D.; Masmoudi, K.; Brini, F. Salt stress reveals differential physiological, biochemical and molecular responses in T. monococcum and T. durum wheat genotypes. Physiol. Mol. Biol. Plants 2017, 23, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.A.; Lin, W.A.N.; Wei, Z.H.; LIU, H.F.; Jun, L.I.; ZHANG, C.L. Exogenous strigolactones promote lateral root growth by reducing the endogenous auxin level in rapeseed. J. Integr. Agric. 2020, 19, 465–482. [Google Scholar]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 217856. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Ashraf, M.; Athar, H.-R. Does exogenous application of 24-epibrassinolide ameliorate salt-induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Elhindi, K.M.; El-Din, A.S.; Elgorban, A.M. The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 2017, 24, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed]
- Boamah, S.; Zhang, S.; Xu, B.; Li, T.; Calderón-Urrea, A.; Tiika, R.J. Trichoderma longibrachiatum TG1 increases endogenous salicylic acid content and antioxidants activity in wheat seedlings under salinity stress. PeerJ 2022, 10, e12923. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Ali, Q.; Jiang, Q.; Wang, R.; Wang, Z.; Mu, G.; Khan, S.A.; Khan, A.R.; Manghwar, H.; Wu, H. Salt tolerant Bacillus strains improve plant growth traits and regulation of phytohormones in wheat under salinity stress. Plants 2022, 11, 2769. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-tolerant plant growth-promoting Bacillus pumilus strain JPVS11 to enhance plant growth attributes of rice and improve soil health under salinity stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Alagoz, S.; Hadi, H.; Toorchi, M.; Pawłowski, T.A.; Asgari Lajayer, B.; Price, G.; Farooq, M.; Astatkie, T. Morpho-physiological responses and growth indices of triticale to drought and salt stresses. Sci. Rep. 2023, 13, 8896. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, S.; Etminan, A.; Rashidi, V.; Pour-Aboughadareh, A.; Shooshtari, L. Delineation of physiological and transcriptional responses of different barley genotypes to salt stress. Cereal Res. Commun. 2023, 51, 367–377. [Google Scholar] [CrossRef]
- Kiani, R.; Arzani, A.; Mirmohammady Maibody, S.; Rahimmalek, M.; Razavi, K. Morpho-physiological and gene expression responses of wheat by Aegilops cylindrica amphidiploids to salt stress. Plant Cell Tissue Organ Cult. 2021, 144, 619–639. [Google Scholar] [CrossRef]
- Yang, Y.; Yao, Y.; Bakpa, E.P.; Xie, J. Trehalose alleviated salt stress in tomato by regulating ROS metabolism, photosynthesis, osmolyte synthesis, and trehalose metabolic pathways. Front. Plant Sci. 2022, 13, 772948. [Google Scholar] [CrossRef] [PubMed]
- Ouertani, R.N.; Jardak, R.; Ben Chikha, M.; Ben Yaala, W.; Abid, G.; Karmous, C.; Hamdi, Z.; Mejri, S.; Jansen, R.K.; Ghorbel, A. Genotype-specific patterns of physiological and antioxidative responses in barley under salinity stress. Cereal Res. Commun. 2022, 50, 851–863. [Google Scholar] [CrossRef]
- Kiliç, G. Farklı Sıcaklık Değerlerinin Penicillium Expansum Fungusunun Protein Içeriği ve Proteaz Enzimi Üzerine Etkileri, Fen Bilimleri Enstitüsü. 2019. Available online: http://hdl.handle.net/11513/2116 (accessed on 19 February 2024).
- Kilinç, N.; Dikilitaş, M.; Tohumcu, E. Adıyaman ilinde Septoria Leke Hastalığının [Zymoseptoria tritici (Desm. Quaedvlieg & Crous)] çıkış zamanı üzerinde iklim faktörlerinin etkisinin belirlenmesi. Harran Tarım Ve Gıda Bilim. Derg. 2021, 25, 304–314. [Google Scholar]
- Morgounov, A.; Sonder, K.; Abugalieva, A.; Bhadauria, V.; Cuthbert, R.D.; Shamanin, V.; Zelenskiy, Y.; DePauw, R.M. Effect of climate change on spring wheat yields in North America and Eurasia in 1981–2015 and implications for breeding. PLoS ONE 2018, 13, e0204932. [Google Scholar] [CrossRef] [PubMed]
- Fones, H.; Gurr, S. The impact of Septoria tritici Blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Curran, E.; Burke, J.I.; Feechan, A. Mixed infections of Zymoseptoria tritici lead to reduced STB disease symptoms. Plant Pathol. 2023, 72, 1202–1217. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Kema, G.; Groenewald, J.; Verkley, G.; Seifbarghi, S.; Razavi, M.; Gohari, A.M.; Mehrabi, R.; Crous, P. Zymoseptoria gen. nov.: A new genus to accommodate Septoria-like species occurring on graminicolous hosts. Persoonia-Mol. Phylogeny Evol. Fungi 2011, 26, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Tiley, A.M.; White, H.J.; Foster, G.D.; Bailey, A.M. The ZtvelB gene is required for vegetative growth and sporulation in the wheat pathogen Zymoseptoria tritici. Front. Microbiol. 2019, 10, 2210. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, Y.; Chaudhari, Y.; Usher, J.; Cairns, T.; Csukai, M.; Haynes, K. A suite of Gateway® compatible ternary expression vectors for functional analysis in Zymoseptoria tritici. Fungal Genet. Biol. 2015, 79, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.B.; Ben M’Barek, S.; Dhillon, B.; Wittenberg, A.H.; Crane, C.F.; Hane, J.K.; Foster, A.J.; Van der Lee, T.A.; Grimwood, J.; Aerts, A. Finished genome of the fungal wheat pathogen Mycosphaerella graminicola reveals dispensome structure, chromosome plasticity, and stealth pathogenesis. PLoS Genet. 2011, 7, e1002070. [Google Scholar] [CrossRef] [PubMed]
- Solomon, P.S. Have we finally opened the door to understanding Septoria tritici blotch disease in wheat? New Phytol. 2017, 214, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Battache, M.; Lebrun, M.-H.; Sakai, K.; Soudière, O.; Cambon, F.; Langin, T.; Saintenac, C. Blocked at the stomatal gate, a key step of wheat Stb16q-mediated resistance to Zymoseptoria tritici. Front. Plant Sci. 2022, 13, 921074. [Google Scholar] [CrossRef]
- Yemelin, A.; Brauchler, A.; Jacob, S.; Foster, A.J.; Laufer, J.; Heck, L.; Antelo, L.; Andresen, K.; Thines, E. Two Novel Dimorphism-Related Virulence Factors of Zymoseptoria tritici Identified Using Agrobacterium-Mediated Insertional Mutagenesis. Int. J. Mol. Sci. 2021, 23, 400. [Google Scholar] [CrossRef] [PubMed]
- Kilaru, S.; Fantozzi, E.; Cannon, S.; Schuster, M.; Chaloner, T.M.; Guiu-Aragones, C.; Gurr, S.J.; Steinberg, G. Zymoseptoria tritici white-collar complex integrates light, temperature and plant cues to initiate dimorphism and pathogenesis. Nat. Commun. 2022, 13, 5625. [Google Scholar] [CrossRef] [PubMed]
- Hehir, J.G.; Connolly, C.; O’Driscoll, A.; Lynch, J.P.; Spink, J.; Brown, J.K.; Doohan, F.; Mullins, E. Temporal and spatial field evaluations highlight the importance of the presymptomatic phase in supporting strong partial resistance in Triticum aestivum against Zymoseptoria tritici. Plant Pathol. 2018, 67, 573–583. [Google Scholar] [CrossRef]
- Bartosiak, S.F.; Arseniuk, E.; Szechyńska-Hebda, M.; Bartosiak, E. Monitoring of natural occurrence and severity of leaf and glume blotch diseases of winter wheat and winter triticale incited by necrotrophic fungi Parastagonospora spp. and Zymoseptoria tritici. Agronomy 2021, 11, 967. [Google Scholar] [CrossRef]
- Li, J.; Zou, X.; Chen, G.; Meng, Y.; Ma, Q.; Chen, Q.; Wang, Z.; Li, F. Potential Roles of 1-Aminocyclopropane-1-carboxylic Acid Synthase Genes in the Response of Gossypium Species to Abiotic Stress by Genome-Wide Identification and Expression Analysis. Plants 2022, 11, 1524. [Google Scholar] [CrossRef]
- Pascholati, S.; Deising, H.; Leiti, B.; Anderson, D.; Nicholson, R. Cutinase and non-specific esterase activities in the conidial mucilage of Colletotrichum graminicola. Physiol. Mol. Plant Pathol. 1993, 42, 37–51. [Google Scholar] [CrossRef]
- Motoyama, T.; Imanishi, K.; Yamaguchi, I. cDNA cloning, expression, and mutagenesis of scytalone dehydratase needed for pathogenicity of the rice blast fungus, Pyricularia oryzae. Biosci. Biotechnol. Biochem. 1998, 62, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Tonukari, N.J.; Scott-Craig, J.S.; Waltonb, J.D. The Cochliobolus carbonum SNF1 gene is required for cell wall–degrading enzyme expression and virulence on maize. Plant Cell 2000, 12, 237–247. [Google Scholar] [PubMed]
- Ababa, G. Biology, taxonomy, genetics, and management of Zymoseptoria tritici: The causal agent of wheat leaf blotch. Mycology 2023, 14, 292–315. [Google Scholar] [CrossRef] [PubMed]
- Motteram, J.; Lovegrove, A.; Pirie, E.; Marsh, J.; Devonshire, J.; van de Meene, A.; Hammond-Kosack, K.; Rudd, J.J. Aberrant protein N-glycosylation impacts upon infection-related growth transitions of the haploid plant-pathogenic fungus Mycosphaerella graminicola. Mol. Microbiol. 2011, 81, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Mehrabi, R.; Zwiers, L.-H.; de Waard, M.A.; Kema, G.H. MgHog1 regulates dimorphism and pathogenicity in the fungal wheat pathogen Mycosphaerella graminicola. Mol. Plant. Microbe Interact. 2006, 19, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Hatta, R.; Ito, K.; Hosaki, Y.; Tanaka, T.; Tanaka, A.; Yamamoto, M.; Akimitsu, K.; Tsuge, T. A conditionally dispensable chromosome controls host-specific pathogenicity in the fungal plant pathogen Alternaria alternata. Genetics 2002, 161, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, B.; Gill, N.; Hamelin, R.C.; Goodwin, S.B. The landscape of transposable elements in the finished genome of the fungal wheat pathogen Mycosphaerella graminicola. BMC Genom. 2014, 15, 1132. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, Z. Distribution of Septoria tritici blotch disease agent Zymoseptoria tritici mating type idiomorphs in Turkey. Plant Prot. Bull. 2020, 60, 33–38. [Google Scholar] [CrossRef]
- Duba, A.; Goriewa-Duba, K.; Wachowska, U. A review of the interactions between wheat and wheat pathogens: Zymoseptoria tritici, Fusarium spp. and Parastagonospora nodorum. Int. J. Mol. Sci. 2018, 19, 1138. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, L.; Reder, M.; Siah, A.; Ducasse, A.; Langlands-Perry, C.; Marcel, T.C.; Morel, J.-B.; Saintenac, C.; Ballini, E. SeptoSympto: A precise image analysis of Septoria tritici blotch disease symptoms using deep learning methods on scanned images. Plant Methods 2024, 20, 18. [Google Scholar] [CrossRef]
- Newey, L.J.; Caten, C.E.; Green, J.R. Rapid adhesion of Stagonospora nodorum spores to a hydrophobic surface requires pre-formed cell surface glycoproteins. Mycol. Res. 2007, 111, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.H.; Solomon, P.S. Extracellular vesicles from the apoplastic fungal wheat pathogen Zymoseptoria tritici. Fungal Biol. Biotechnol. 2020, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Haddad Momeni, M.; Bollella, P.; Ortiz, R.; Thormann, E.; Gorton, L.; Abou Hachem, M. A novel starch-binding laccase from the wheat pathogen Zymoseptoria tritici highlights the functional diversity of ascomycete laccases. BMC Biotechnol. 2019, 19, 61. [Google Scholar] [CrossRef]
- Schwarczinger, I.; Kolozsváriné Nagy, J.; Király, L.; Mészáros, K.; Bányai, J.; Kunos, V.; Fodor, J.; Künstler, A. Heat stress pre-exposure may differentially modulate plant defense to powdery mildew in a resistant and susceptible barley genotype. Genes 2021, 12, 776. [Google Scholar] [CrossRef] [PubMed]
- Francisco, C.S.; Ma, X.; Zwyssig, M.M.; McDonald, B.A.; Palma-Guerrero, J. Morphological changes in response to environmental stresses in the fungal plant pathogen Zymoseptoria tritici. Sci. Rep. 2019, 9, 9642. [Google Scholar] [CrossRef]
- Qin, R.; LeBoldus, J.M. The infection biology of Sphaerulina musiva: Clues to understanding a forest pathogen. PLoS ONE 2014, 9, e103477. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, P.; Zafari, D.; Mahmoodi, S.B.; Hashemi, M.; Gholamhoseini, M.; Dolatabadian, A.; Ataei, R. Histopathology of charcoal rot disease (Macrophomina phaseolina) in resistant and susceptible cultivars of soybean. Rhizosphere 2018, 7, 27–34. [Google Scholar] [CrossRef]
- Yang, N.; McDonald, M.C.; Solomon, P.S.; Milgate, A.W. Genetic mapping of Stb19, a new resistance gene to Zymoseptoria tritici in wheat. Theor. Appl. Genet. 2018, 131, 2765–2773. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Chartrain, L.; Lasserre-Zuber, P.; Saintenac, C. Genetics of resistance to Zymoseptoria tritici and applications to wheat breeding. Fungal Genet. Biol. 2015, 79, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Siah, A.; Deweer, C.; Duyme, F.; Sanssené, J.; Durand, R.; Halama, P.; Reignault, P. Correlation of in planta endo-beta-1, 4-xylanase activity with the necrotrophic phase of the hemibiotrophic fungus Mycosphaerella graminicola. Plant Pathol. 2010, 59, 661–670. [Google Scholar] [CrossRef]
- Shetty, N.; Kristensen, B.; Newman, M.-A.; Møller, K.; Gregersen, P.L.; Jørgensen, H.L. Association of hydrogen peroxide with restriction of Septoria tritici in resistant wheat. Physiol. Mol. Plant Pathol. 2003, 62, 333–346. [Google Scholar] [CrossRef]
- Seybold, H.; Demetrowitsch, T.J.; Hassani, M.; Szymczak, S.; Reim, E.; Haueisen, J.; Lübbers, L.; Rühlemann, M.; Franke, A.; Schwarz, K. A fungal pathogen induces systemic susceptibility and systemic shifts in wheat metabolome and microbiome composition. Nat. Commun. 2020, 11, 1910. [Google Scholar] [CrossRef] [PubMed]
- van der Burgh, A.M.; Joosten, M.H. Plant immunity: Thinking outside and inside the box. Trends Plant Sci. 2019, 24, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Saintenac, C.; Cambon, F.; Aouini, L.; Verstappen, E.; Ghaffary, S.M.T.; Poucet, T.; Marande, W.; Berges, H.; Xu, S.; Jaouannet, M. A wheat cysteine-rich receptor-like kinase confers broad-spectrum resistance against Septoria tritici blotch. Nat. Commun. 2021, 12, 433. [Google Scholar] [CrossRef]
- Habig, M.; Bahena-Garrido, S.M.; Barkmann, F.; Haueisen, J.; Stukenbrock, E.H. The transcription factor Zt107320 affects the dimorphic switch, growth and virulence of the fungal wheat pathogen Zymoseptoria tritici. Mol. Plant Pathol. 2020, 21, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, N.; Mehrabi, R.; Gohari, A.M.; Goltapeh, E.M.; Safaie, N.; Kema, G.H. The ZtVf1 transcription factor regulates development and virulence in the foliar wheat pathogen Zymoseptoria tritici. Fungal Genet. Biol. 2017, 109, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, N.; Mehrabi, R.; Mirzadi Gohari, A.; Roostaei, M.; Mohammadi Goltapeh, E.; Safaie, N.; Kema, G.H. MADS-Box Transcription Factor ZtRlm1 Is Responsible for Virulence and Development of the Fungal Wheat Pathogen Zymoseptoria tritici. Front. Microbiol. 2020, 11, 1976. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Yu, D.; Peng, Y.; Min, H.; Lai, Z. Copper Ions are Required for Cochliobolus heterostrophus in Appressorium Formation and Virulence on Maize. Phytopathology 2020, 110, 494–504. [Google Scholar] [CrossRef]
- Kou, Y.; Qiu, J.; Tao, Z. Every coin has two sides: Reactive oxygen species during Rice–Magnaporthe oryzae interaction. Int. J. Mol. Sci. 2019, 20, 1191. [Google Scholar] [CrossRef] [PubMed]
- Choquer, M.; Rascle, C.; Gonçalves, I.R.; de Vallée, A.; Ribot, C.; Loisel, E.; Smilevski, P.; Ferria, J.; Savadogo, M.; Souibgui, E. The infection cushion of Botrytis cinerea: A fungal ‘weapon’of plant-biomass destruction. Environ. Microbiol. 2021, 23, 2293–2314. [Google Scholar] [CrossRef] [PubMed]
- Ors, M.; Randoux, B.; Siah, A.; Couleaud, G.; Maumené, C.; Sahmer, K.; Reignault, P.; Halama, P.; Selim, S. A plant nutrient-and microbial protein-based resistance inducer elicits wheat cultivar-dependent resistance against Zymoseptoria tritici. Phytopathology 2019, 109, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Eyal, Z. The Septoria Diseases of Wheat: Concepts and Methods of Disease Management; Cimmyt: Mexico City, Mexico, 1987; ISBN 968-6127-06-2. [Google Scholar]
- Agrios, G.N. Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005; ISBN 0-08-047378-4. [Google Scholar]
- Ponomarenko, A.; Goodwin, S.B.; Kema, G.H. Septoria tritici Blotch (STB) of Wheat. 2011. Available online: https://www.apsnet.org/edcenter/disandpath/fungalasco/pdlessons/Pages/Septoria.aspx (accessed on 19 February 2024). [CrossRef]
- El-Gamal, N.G.; El-Mougy, N.S.; Abdel-Kader, M.M.; Ali Khalil, M.S. Influence of inorganic salts and chitosan as foliar spray on wheat septoria leaf blotch disease severity under field conditions. Arch. Phytopathol. Plant Prot. 2021, 54, 836–849. [Google Scholar] [CrossRef]
- De Borba, M.C.; Velho, A.C.; Maia-Grondard, A.; Baltenweck, R.; Magnin-Robert, M.; Randoux, B.; Holvoet, M.; Hilbert, J.-L.; Flahaut, C.; Reignault, P. The algal polysaccharide ulvan induces resistance in wheat against Zymoseptoria tritici without major alteration of leaf metabolome. Front. Plant Sci. 2021, 12, 703712. [Google Scholar] [CrossRef] [PubMed]
- Mejri, S.; Magnin-Robert, M.; Randoux, B.; Ghinet, A.; Halama, P.; Siah, A.; Reignault, P. Saccharin provides protection and activates defense mechanisms in wheat against the Hemibiotrophic pathogen Zymoseptoria tritici. Plant Dis. 2021, 105, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Allioui, N.; Driss, F.; Dhouib, H.; Jlail, L.; Tounsi, S.; Frikha-Gargouri, O. Two Novel Bacillus Strains (Subtilis and Simplex Species) with Promising Potential for the Biocontrol of Zymoseptoria tritici, the Causal Agent of Septoria tritici Blotch of Wheat. BioMed Res. Int. 2021, 2021, 6611657. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Faris, J.; Meinhardt, S.; Ali, S.; Rasmussen, J.; Friesen, T. Genetic and physical mapping of a gene conditioning sensitivity in wheat to a partially purified host-selective toxin produced by Stagonospora nodorum. Phytopathology 2004, 94, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Mirzadi Gohari, A.; Fallahi, M.; Mohammadi, N. Investigation on TOX2 cluster genes of the fungal wheat pathogen Zymoseptoria tritici. Eur. J. Plant Pathol. 2022, 165, 17–26. [Google Scholar] [CrossRef]
- Rodrigues, A.I.; Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Sodium chloride effect on the aggregation behaviour of rhamnolipids and their antifungal activity. Sci. Rep. 2017, 7, 12907. [Google Scholar] [CrossRef] [PubMed]
- Podder, S.; Saha, D.; Ghosh, T.C. Deciphering the intrinsic properties of fungal proteases in optimizing phytopathogenic interaction. Gene 2019, 711, 143934. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Kautto, L.; Nevalainen, H. Secretion of proteases by an opportunistic fungal pathogen Scedosporium aurantiacum. PLoS ONE 2017, 12, e0169403. [Google Scholar] [CrossRef]
- Ben Ali, W.; Chaduli, D.; Navarro, D.; Lechat, C.; Turbé-Doan, A.; Bertrand, E.; Faulds, C.B.; Sciara, G.; Lesage-Meessen, L.; Record, E. Screening of five marine-derived fungal strains for their potential to produce oxidases with laccase activities suitable for biotechnological applications. BMC Biotechnol. 2020, 20, 27. [Google Scholar] [CrossRef] [PubMed]
- Damare, S.; Raghukumar, C.; Muraleedharan, U.D.; Raghukumar, S. Deep-sea fungi as a source of alkaline and cold-tolerant proteases. Enzyme Microb. Technol. 2006, 39, 172–181. [Google Scholar] [CrossRef]
- Malik, P.L.; Janss, L.; Nielsen, L.K.; Borum, F.; Jørgensen, H.; Eriksen, B.; Schjoerring, J.K.; Rasmussen, S.K. Breeding for dual-purpose wheat varieties using marker–trait associations for biomass yield and quality traits. Theor. Appl. Genet. 2019, 132, 3375–3398. [Google Scholar] [CrossRef] [PubMed]
- Amezrou, R.; Audéon, C.; Compain, J.; Gélisse, S.; Ducasse, A.; Saintenac, C.; Lapalu, N.; Louet, C.; Orford, S.; Croll, D. A secreted protease-like protein in Zymoseptoria tritici is responsible for avirulence on Stb9 resistance gene in wheat. PLoS Pathog. 2023, 19, e1011376. [Google Scholar] [CrossRef] [PubMed]
- Somai-Jemmali, L.; Siah, A.; Harbaoui, K.; Fergaoui, S.; Randoux, B.; Magnin-Robert, M.; Halama, P.; Reignault, P.; Hamada, W. Correlation of fungal penetration, CWDE activities and defense-related genes with resistance of durum wheat cultivars to Zymoseptoria tritici. Physiol. Mol. Plant Pathol. 2017, 100, 117–125. [Google Scholar] [CrossRef]
- Tian, S.M.; Weinert, J.; Zhao, Q.H. Correlation between cell wall-degrading enzymes in wheat leaves infected by Septoria tritici and disease severity. Can. J. Plant Pathol. 2009, 31, 387–392. [Google Scholar] [CrossRef]
- Derbyshire, M.C.; Gohari, A.M.; Mehrabi, R.; Kilaru, S.; Steinberg, G.; Ali, S.; Bailey, A.; Hammond-Kosack, K.; Kema, G.H.; Rudd, J.J. Phosphopantetheinyl transferase (Ppt)-mediated biosynthesis of lysine, but not siderophores or DHN melanin, is required for virulence of Zymoseptoria tritici on wheat. Sci. Rep. 2018, 8, 17069. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.F.; da Rocha, I.M.; Vieira-da-Motta, O.; de Paula Santos, C. The Role of Melanin in the Biology and Ecology of Nematophagous Fungi. J. Chem. Ecol. 2021, 47, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Aranda, C.; Méndez, I.; Barra, P.J.; Hernández-Montiel, L.; Fallard, A.; Tortella, G.; Briones, E.; Durán, P. Melanin Induction Restores the Pathogenicity of Gaeumannomyces graminis var. tritici in Wheat Plants. J. Fungi 2023, 9, 350. [Google Scholar] [CrossRef] [PubMed]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P.; Georgeault, S.; Mallet, R.; Chabasse, D.; Symoens, F.; Bouchara, J.-P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Tiley, A.M.; Lawless, C.; Pilo, P.; Karki, S.J.; Lu, J.; Long, Z.; Gibriel, H.; Bailey, A.M.; Feechan, A. The Zymoseptoria tritici white collar-1 gene, ZtWco-1, is required for development and virulence on wheat. Fungal Genet. Biol. 2022, 161, 103715. [Google Scholar] [CrossRef]
- Gong, X.; Feng, S.; Zhao, J.; Tang, C.; Tian, L.; Fan, Y.; Cao, Z.; Hao, Z.; Jia, H.; Zang, J. StPBS2, a MAPK kinase gene, is involved in determining hyphal morphology, cell wall development, hypertonic stress reaction as well as the production of secondary metabolites in Northern Corn Leaf Blight pathogen Setosphaeria turcica. Microbiol. Res. 2017, 201, 30–38. [Google Scholar] [PubMed]
- Hagiwara, D.; Sakai, K.; Suzuki, S.; Umemura, M.; Nogawa, T.; Kato, N.; Osada, H.; Watanabe, A.; Kawamoto, S.; Gonoi, T. Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 2017, 12, e0177050. [Google Scholar]
- Dura, S.A.; Duwayri, M.A.; Nachit, M.M. Effects of different salinity levels on growth, yield and physiology on durum wheat (Triticum turgidum var. durum). Jordan J. Agric. Sci. 2011, 7, 527–528. [Google Scholar]
- You, T.; Ingram, P.; Jacobsen, M.D.; Cook, E.; McDonagh, A.; Thorne, T.; Lenardon, M.D.; De Moura, A.P.; Romano, M.C.; Thiel, M. A systems biology analysis of long and short-term memories of osmotic stress adaptation in fungi. BMC Res. Notes 2012, 5, 258. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Linares, D.R.; Veresoglou, S.D.; Rillig, M.C. Temperature priming and memory in soil filamentous fungi. Fungal Ecol. 2016, 21, 10–15. [Google Scholar] [CrossRef]
- Boumaaza, B.; Benkhelifa, M.; Belkhoudja, M. Effects of two salts compounds on mycelial growth, sporulation, and spore germination of six isolates of Botrytis cinerea in the western north of Algeria. Int. J. Microbiol. 2015, 2015, 572626. [Google Scholar] [CrossRef] [PubMed]
- Dikilitas, M.; Karakas, S. Salts as potential environmental pollutants, their types, effects on plants and approaches for their phytoremediation. In Plant Adaptation and Phytoremediation; Springer: Berlin/Heidelberg, Germany, 2010; pp. 357–381. [Google Scholar]
- Dikilitas, M.; Karakas, S. Crop plants under saline-adapted fungal pathogens: An Overview. Emerg. Technol. Manag. Crop Stress Toler. 2014, 2014, 173–192. [Google Scholar]
- Kilinç, N.; Dikilitaş, M.; Kayim, M.; Gülsüm, Ü. Septoria yaprak leke hastalığı etmeni Zymoseptoria tritici (Desm. Quaedvlieg & Crous)’ye ait izolatların farklı sıcaklıklardaki fizyolojik ve biyokimyasal özelliklerin belirlenmesi. Harran Tarım Ve Gıda Bilim. Derg. 2021, 25, 469–479. [Google Scholar]
- .Pazhamala, L.T.; Giri, J. Plant phosphate status influences root biotic interactions. J. Exp. Bot. 2023, 74, 2829–2844. [Google Scholar] [CrossRef] [PubMed]
- Antal, K.; Gila, B.C.; Pocsi, I.; Emri, T. General stress response or adaptation to rapid growth in Aspergillus nidulans? Fungal Biol. 2020, 124, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.-S.; Wilkolazka, A.J.; Staszczak, M.; Cho, H.-Y.; Ohga, S. The Role of Laccase from White Rot Fungi to Stress Conditions; Faculty of Agriculture, Kyushu University: Fukuoka, Japan, 2009; pp. 81–83. [Google Scholar]
- Wangsanut, T.; Pongpom, M. The role of the glutathione system in stress adaptation, morphogenesis and virulence of pathogenic fungi. Int. J. Mol. Sci. 2022, 23, 10645. [Google Scholar] [CrossRef] [PubMed]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.-P.; Papon, N. Oxidative stress response pathways in fungi. Cell. Mol. Life Sci. 2022, 79, 333. [Google Scholar] [CrossRef] [PubMed]
- Clair, S.B.S.; Carlson, J.E.; Lynch, J.P. Evidence for oxidative stress in sugar maple stands growing on acidic, nutrient imbalanced forest soils. Oecologia 2005, 145, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Fungal adaptation to extremely high salt concentrations. Adv. Appl. Microbiol. 2011, 77, 71–96. [Google Scholar] [PubMed]
- McCorison, C.B.; Goodwin, S.B. The wheat pathogen Zymoseptoria tritici senses and responds to different wavelengths of light. BMC Genom. 2020, 21, 513. [Google Scholar] [CrossRef] [PubMed]
- Karakas, S. Development of Tomato Growing in Soil Differing in Salt Levels and Effects of Companion Plants on Same Physiological Parameters and Soil Remediation. Ph.D. Thesis, University of Harran, Sanlıurfa, Turkey, 2013. [Google Scholar]
- Kashyap, P.L.; Solanki, M.K.; Kushwaha, P.; Kumar, S.; Srivastava, A.K. Biocontrol potential of salt-tolerant Trichoderma and Hypocrea isolates for the management of tomato root rot under saline environment. J. Soil Sci. Plant Nutr. 2020, 20, 160–176. [Google Scholar] [CrossRef]
- Daami-Remadi, M.; Souissi, A.; Oun, H.B.; Mansour, M.; Nasraoui, B. Salinity effects on Fusarium wilt severity and tomato growth. Dyn. Soil Dyn. Plant 2009, 3, 61–69. [Google Scholar]
- Dikilitas, M.; Yucel, N.; Dervis, S. Production of antioxidant and oxidant metabolites in tomato plants infected with verticillium dahliae under saline conditions. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Springer: Berlin/Heidelberg, Germany, 2017; pp. 315–329. [Google Scholar]
- Qiu, Z.; Li, Z.; Fu, H.; Zhang, H.; Zhu, Z.; Wang, A.; Li, H.; Zhang, L.; Zhang, H. Corrosion mechanisms of Zr-based bulk metallic glass in NaF and NaCl solutions. J. Mater. Sci. Technol. 2020, 46, 33–43. [Google Scholar] [CrossRef]
- Gonçalves, D.R.; Pena, R.; Zotz, G.; Albach, D.C. Effects of fungal inoculation on the growth of Salicornia (Amaranthaceae) under different salinity conditions. Symbiosis 2021, 84, 195–208. [Google Scholar] [CrossRef]
- Yan, S.; Chong, P.; Zhao, M. Effect of salt stress on the photosynthetic characteristics and endogenous hormones, and: A comprehensive evaluation of salt tolerance in Reaumuria soongorica seedlings. Plant Signal. Behav. 2022, 17, 2031782. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, Y.M.; Heo, Y.M.; Lee, H.; Hong, J.-H.; Jang, S.; Park, M.S.; Lim, Y.W.; Kim, J.-J. Halo-tolerance of marine-derived fungi and their enzymatic properties. BioResources 2015, 10, 8450–8460. [Google Scholar] [CrossRef]
- Vashishtha, A.; Meghwanshi, G.K. Fungi inhabiting in hypersaline conditions: An insight. In Fungi and Their Role in Sustainable Development: Current Perspectives; Springer: Berlin/Heidelberg, Germany, 2018; pp. 449–465. [Google Scholar]
- Manjunatha, B.S.; Nivetha, N.; Krishna, G.K.; Elangovan, A.; Pushkar, S.; Chandrashekar, N.; Aggarwal, C.; Asha, A.D.; Chinnusamy, V.; Raipuria, R.K. Plant growth-promoting rhizobacteria Shewanella putrefaciens and Cronobacter dublinensis enhance drought tolerance of pearl millet by modulating hormones and stress-responsive genes. Physiol. Plant. 2022, 174, e13676. [Google Scholar] [CrossRef] [PubMed]
- Kamei, I.; Tomitaka, N.; Yamasaki, Y. Selective Homologous Expression of Recombinant Manganese Peroxidase Isozyme of Salt-Tolerant White-Rot Fungus Phlebia sp. MG-60, and Its Salt-Tolerance and Thermostability. J. Microbiol. Biotechnol 2022, 32, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Pombeiro-Sponchiado, S.R.; Sousa, G.S.; Andrade, J.C.; Lisboa, H.F.; Gonçalves, R.C. Production of melanin pigment by fungi and its biotechnological applications. In Melanin; InTech: Rijeka, Croatia, 2017; pp. 47–75. [Google Scholar] [CrossRef]
- Pandey, P.; Patil, M.; Priya, P.; Senthil-Kumar, M. When two negatives make a positive: The favorable impact of the combination of abiotic stress and pathogen infection on plants. J. Exp. Bot. 2024, 75, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.Y.; Zhao, W.S.; Chen, Z.; Xing, Q.K.; Zhang, W.; Chethana, K.T.; Xue, M.F.; Xu, J.P.; Phillips, A.J.; Wang, Y. Comparative genome and transcriptome analyses reveal adaptations to opportunistic infections in woody plant degrading pathogens of Botryosphaeriaceae. DNA Res. 2018, 25, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Rubinowska, K. 20 Role of Polyamines in Abiotic and Biotic Stress Tolerance in Plants. Biostimul. Crop Prod. Sustain. Agric. 2022, 323, 87–102. [Google Scholar] [CrossRef] [PubMed]
- de Leite, M.L.; da Silva, M.C.; Simões, V.J.; de Lucena, L.R.; Sales, A.T. Biostimulant use in pangolão grass Digitaria pentzii subjected to saline stress. Rev. Bras. Eng. Agríc. E Ambient. 2022, 27, 26–33. [Google Scholar] [CrossRef]
- Wang, X.; Ma, R.; Cao, Q.; Shan, Z.; Jiao, Z. Enhanced tolerance to salt stress in highland barley seedlings (Hordeum vulgare ssp. vulgare) by gamma irradiation pretreatment. Acta Physiol. Plant. 2018, 40, 174. [Google Scholar] [CrossRef]
- Shunkao, S.; Thitisaksakul, M.; Pongdontri, P.; Theerakulpisut, P. Additive effects of combined heat and salt stress is manifested in an enhanced sodium ions accumulation and increased membrane damage in wheat seedlings. Chil. J. Agric. Res. 2022, 82, 552–563. [Google Scholar] [CrossRef]
- Yu, Y.; Assmann, S.M. The effect of NaCl on stomatal opening in Arabidopsis wild type and agb1 heterotrimeric G-protein mutant plants. Plant Signal. Behav. 2016, 11, e1085275. [Google Scholar] [CrossRef] [PubMed]
- Al-Hammouri, A.A.; Ibbini, J.; Bsoul, E.; Sanogo, S. Effect of salinity on recovery of Rhizoctonia solani from infected tomato (Solanum lycopersicon mill). Bulg. J. Agric. Sci. 2017, 23, 757–761. [Google Scholar]
- Javed, S.; Javaid, A.; Hanif, U.; Bahadur, S.; Sultana, S.; Shuaib, M.; Ali, S. Effect of necrotrophic fungus and PGPR on the comparative histochemistry of Vigna radiata by using multiple microscopic techniques. Microsc. Res. Tech. 2021, 84, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- De Pascali, M.; Vergine, M.; Sabella, E.; Aprile, A.; Nutricati, E.; Nicolì, F.; Buja, I.; Negro, C.; Miceli, A.; Rampino, P. Molecular effects of Xylella fastidiosa and drought combined stress in olive trees. Plants 2019, 8, 437. [Google Scholar] [CrossRef] [PubMed]
- Weldon, W.A.; Palumbo, C.D.; Kovaleski, A.P.; Tancos, K.; Gadoury, D.M.; Osier, M.V.; Cadle-Davidson, L. Transcriptomic profiling of acute cold stress-induced disease resistance (SIDR) genes and pathways in the grapevine powdery mildew pathosystem. Mol. Plant. Microbe Interact. 2020, 33, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Dikilitas, M.; Karakas, S.; Simsek, E.; Yadav, A.N. Microbes from cold deserts and their applications in mitigation of cold stress in plants. In Microbiomes of Extreme Environments; CRC Press: Boca Raton, FL, USA, 2021; pp. 126–152. [Google Scholar]
- Dikilitas, M.; Karakas, S.; Hashem, A.; Abd Allah, E.; Ahmad, P. Oxidative stress and plant responses to pathogens under drought conditions. Water Stress Crop Plants Sustain. Approach 2016, 1, 102–123. [Google Scholar]
- Shikari, A.B.; Dikilitas, M.; Guldur, M.E.; Simsek, E.; Kaya Demirsoy, F.; Sakina, A.; Abdel Latef, A.A.H. Cereals Under Abiotic Stress: An Overview. In Sustainable Remedies for Abiotic Stress in Cereals; Springer: Singapore, 2022; pp. 3–27. [Google Scholar] [CrossRef]
- DiLeo, M.V.; Pye, M.F.; Roubtsova, T.V.; Duniway, J.M.; MacDonald, J.D.; Rizzo, D.M.; Bostock, R.M. Abscisic acid in salt stress predisposition to Phytophthora root and crown rot in tomato and chrysanthemum. Phytopathology 2010, 100, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sengar, R.; Pathak, R.K.; Singh, A.K. Integrated approaches to develop drought-tolerant rice: Demand of era for global food security. J. Plant Growth Regul. 2023, 42, 96–120. [Google Scholar] [CrossRef]
- Gul, S.; Hussain, A.; Ali, Q.; Alam, I.; Alshegaihi, R.M.; Meng, Q.; Zaman, W.; Manghwar, H.; Munis, M.F.H. Hydropriming and osmotic priming induce resistance against Aspergillus niger in wheat (Triticum aestivum L.) by activating β-1, 3-glucanase, chitinase, and thaumatin-like protein genes. Life 2022, 12, 2061. [Google Scholar] [CrossRef] [PubMed]
- Habora, M.E.E.; Eltayeb, A.E.; Oka, M.; Tsujimoto, H.; Tanaka, K. Cloning of allene oxide cyclase gene from Leymus mollis and analysis of its expression in wheat–Leymus chromosome addition lines. Breed. Sci. 2013, 63, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Chalbi, A.; Sghaier-Hammami, B.; Baazaoui, N.; Hammami, S.B.; Hatem, B.-J.; Garcia-Caparros, P.; Djebali, N.; Regaya, I.; Debez, A.; Jesús, V. Comparative study of the effect of salt stress, Alternaria alternata attack or combined stress on the Cakile maritima growth and physiological performance. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12446. [Google Scholar] [CrossRef]
- Gupta, A.; Patil, M.; Qamar, A.; Senthil-Kumar, M. ath-miR164c influences plant responses to the combined stress of drought and bacterial infection by regulating proline metabolism. Environ. Exp. Bot. 2020, 172, 103998. [Google Scholar] [CrossRef]
- Naz, R.; Batool, S.; Shahid, M.; Keyani, R.; Yasmin, H.; Nosheen, A.; Hassan, M.N.; Mumtaz, S.; Siddiqui, M.H. Exogenous silicon and hydrogen sulfide alleviates the simultaneously occurring drought stress and leaf rust infection in wheat. Plant Physiol. Biochem. 2021, 166, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Stocco, M.C.; Mansilla, A.Y.; Mónaco, C.I.; Segarra, C.; Lampugnani, G.; Abramoff, C.; Marchetti, M.F.; Kripelz, N.; Cordo, C.C.; Consolo, V.F. Native isolates of Trichoderma harzianum inducting resistance to Zymoseptoria tritici on wheat plants. Bol. Soc. Argent. Botánica 2015, 50, 291–301. [Google Scholar] [CrossRef]
- Korostyleva, T.V.; Shiyan, A.N.; Odintsova, T.I. The Genetic Resource of Thinopyrum elongatum (Host) DR Dewey in Breeding Improvement of Wheat. Russ. J. Genet. 2023, 59, 983–990. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity Stress (Leaf/Root) | Characteristic Symptoms and Improvement Strategies | References |
|---|---|---|
| 150 mmol L−1 NaCl + Drought (Root) | Salinity and water deprivation caused significant growth retardation and a 3-fold reduction in the osmotic potential in wheat seedlings. Polyvinylpyrrolidone (PVP)-coated Cu nanoparticles in Triticum aestivum under salt and/or drought stress restored the growth rate and regulated water uptake. | [57] |
| 120 mmol L−1 NaCl, Root | Exogenous silicon nanoparticles (SiNPs) on Elymus sibiricus seeds increased the chlorophyll and osmolyte accumulation and improved the activities of SOD, CAT, POD, and APX. | [58] |
| Root (2% NaCl) | Exogenous sodium nitroprusside (SNP, a donor of NO) increased the lignin and salicylic acid (SA) contents under normal and salinity (2% NaCl) conditions via triggering the phenylpropanoid pathway as well as increasing the level of transcription of the pathogenesis-related protein 1 (PR1), TaPAL and TaPRX genes responsible for the syntheses of POD, PAL, and tyrosine ammonia lyase (TAL) enzymes. | [59] |
| Root (0–250 mmol L−1 NaCl) | Wild common wheat species were proposed for the improvement of wheat cultivars due to possessing resistance genes to various environmental stresses via synthesizing CAT, SOD, and APX and expressing their encoding genes. | [60] |
| Root (50–150 mmol L−1 NaCl) | Seed priming with NaCl was suggested to improve plant growth by altering the Na+ and K+ content under salt stress. | [61] |
| Root (150–200 mmol L−1 NaCl | The roles of the microRNA (miRNA) family in modulating plant growth and development through tae-miR9674a were proposed as an essential mediator in the wheat plant for the osmotic stress tolerance, cellular ROS homeostasis, and defensive processes. | [62] |
| Root (0–200 mmol L−1 NaCl) | Exogenous jasmonic acid (JA) significantly improved the salt tolerance of wheat seedlings via alleviating membrane lipid peroxidation and enhanced the contents of abscisic acid (ABA), jasmonic acid (JA), and salicylic acid (SA). In the RNA-seq profiles, 2% of the unigenes were differentially expressed upon application. | [63] |
| Root (0–250 mmol L−1 NaCl) | SA application enhanced the salt tolerance of perennial ryegrass (Lolium perenne L.) via improving photosynthesis, stomatal conductance, and defense systems. | [64] |
| Root (250 mmol L−1 NaCl) | Salt-pretreated plants maintained higher photosynthetic efficacy and lower proline and MDA contents than non-pretreated plants. Salt-pretreated plants also sustained high expressional levels of salt-responsive genes (TaNHX1, TaSOS1, TaSOS4, TaHKT1, TaHKT2, and TaAKT1). | [55] |
| Root (350 mmol L−1 NaCl) | Two neglected (Aegilops triuncialis and Ae. tauschii) wheat genotypes responded better to salinity stress, and they were suggested for breeding lines to improve salt tolerance in wheat plants. | [65] |
| Root (150–250 mmol L−1 NaCl) | Triticum aestivum L. cv. Norin 61 treated with 2,4-dichlorophenoxyacetic acid (2,4-D) herbicide increased the glyoxalase enzyme activity that enhanced salt tolerance via inducing methylglyoxal (MG) detoxification systems. | [56] |
| Root (0–200 mmol L−1 NaCl) | The salt-tolerant diploid (Triticum monococcum cv. Turkey) cultivar accumulated less toxic sodium in the photosynthetic tissues with the high expression of SOS1, HKT, and NHX genes in the leaf blade than the salt-sensitive tetraploid wheat (T. durum cv. Om Rabia3). | [66] |
| Leaf (0–100 mmol L−1 NaCl) | GR24, a synthesized strigolactone plant hormone, alleviated salinity tolerance in rapeseed (Brassica napus L.) | [67] |
| Root (150 mmol L−1 NaCl) | Trichoderma longibrachiatum T6 (T6) promoted wheat growth and induced plant resistance under saline conditions. | [68] |
| Leaf (0–100 mmol L−1 NaCl) | Foliarly applied 24-epibrassionlide (24-epiBL) alleviated the adverse effects of salt on wheat by enhancing the photosystem-II efficiency. | [69] |
| Root (EC = 10 dS m−1 NaCl) | The application of arbuscular mycorrhizal (Glomus deserticola) mitigated the reduction of K, P, and Ca uptake in sweet basil (Osmium basilicum) plants under high saline conditions. | [70] |
| Root (0–200 mmol L−1 NaCl) | The plant growth-promoting (PGP) bacterial isolate CDP-13 from Serratia marcescens enhanced the growth of wheat plants up to the 6% NaCl level in vitro and modulated the osmoprotectants (proline, malondialdehyde, total soluble sugar, total protein content, and indole acetic acid) to enable salt tolerance. | [71] |
| Root (0–150 mmol L−1 NaCl) | Pretreatment of Triticum aestivum L. seed with Trichoderma longibrachiatum (TG1) increased seed germination and increased the SA concentration, PAL activity, and the defense-related gene expression under saline conditions. | [72] |
| Root (200 mmol L−1 NaCl) | Highly halophilic Bacillus strains, isolated from the rhizosphere in the extreme environment, significantly enhanced wheat growth parameters under saline conditions. | [73] |
| Root (0–150 mmol L−1 NaCl | Saline-adapted Bacillus pumilus strain JPVS11 improved the growth performance of rice (Oryza sativa L.) under saline conditions as well as increasing soil enzyme activities. | [74] |
| Root (0–200 mmol L−1 NaCl) | The combined effects of drought and salt stresses increased soluble sugars, leaf-free proline, carotenoid contents, and electrolyte leakage. The authors suggested that these parameters could be used as physiological indicators of drought and salinity tolerance in triticale. | [75] |
| Root (0–200 mmol L−1 NaCl) | Salt treatment significantly increased the root and shoot Na+ contents, and the activity of APX and GPX antioxidant enzymes, as well as increasing the relative expressions of HvHKT2, HvHKT3, HvSOS1, HvSOS3, HvNHX1, HvNHX3, APX, and GPX genes. The genotypes responded better to salinity and could be selected for the aim of releasing commercial varieties. | [76] |
| Leaf (250 mmol L−1 NaCl) | Amphidiploids of Aegilops cylindrica, the salt-tolerant species in the Triticeae tribe, showed significant expression patterns of the HKT1;5, NHX1, and SOS1 genes associated with the exclusion of Na+ from the roots. | [77] |
| Root (0–200 mmol L−1 NaCl) | Trehalose application significantly increased the enzymatic activity of the trehalose metabolic pathway and increased plant biomass in tomatoes. | [78] |
| Root (0.200 mmol L−1 NaCl) | The salt-tolerant Boulifa (B) variety of barley expressed higher activities of antioxidant enzymes (SOD, APX, glutathione peroxidase, GPX, and glutathione reductase, GR) and the corresponding genes than the salt-sensitive Manzel Habib (MH) genotypes under salinity stress. | [79] |
| Soil Salinity Class | Electrical Conductivity (μS/cm) of the Soil and the Corresponding NaCl Concentration (mmol L−1) | Effect on Plants | Effect on Pathogenic Microorganisms | |
|---|---|---|---|---|
| EC | NaCl (mmol L−1) * | |||
| No detectable salt | 0–2.41 | 0–25 | The effect of salinity is negligible [168]. | The effect on pathogenic microorganisms is negligible [7]. |
| Lightly salted | 2.41–4.83 | 25–50 | Salt-sensitive plants may exhibit growth retardation [168]. | Pathogenic microorganisms develop without any retardation [7]. |
| Medium salinity | 4.80–9.96 | 50–100 | The development of many plants slows down and causes symptoms [72]. | Pathogens can easily develop. Growth can be stimulated. The pathogenicity may increase [6]. |
| Medium–high salinity | 9.96–15.78 | 100–150 | Vegetative development of crop plants is restricted. The defense system is significantly impaired [169]. | Pathogens can develop. Sporulation and mycelial development are delayed, but ample sporulation is achieved [170]. |
| High salinity (stage I) | 15.78–19.61 | 150–200 | Only tolerant plants can grow. Halophytes grow well. Other plants may produce very few flowers [168,171] | Pathogens can easily develop. Sporulation is reduced, and mycelial growth may show hyaline development [7]. |
| High salinity (stage II) | 19.61–24.56 | 200–250 | Few tolerant plants can develop. Only halotolerant plants can grow well. Glycophytes show severe growth retardation [168]. | Pathogens are stressed, and sporulation and mycelial growth decrease but continue. Stress metabolites of the pathogen tend to increase [5]. |
| Very high salinity (stage I) | 24.56–29.45 | 250–300 | The development of salt-tolerant plants is retarded. Only obligate halotolerant plants can grow [2,168]. | Sporulation and mycelial growth of pathogens are inhibited, but they can sporulate sufficiently for pathogenicity [46,171] |
| Very high salinity (stage II) | 29.45–39.32 | 300–400 | Growth retardation is observed in obligate halotolerant plants. Salt-tolerant glycophytes do not develop and cannot produce products [168]. | The development of pathogens is severely restricted, and the infection capacity continues. Halotolerant microorganisms can easily develop [170]. |
| Extreme salinity (stage I) | 39.32–49.15 | 400–500 | Except for obligate halophytes, plants cannot grow. Salt-tolerant glycophytes cannot grow or germinate [168]. | Salt-tolerant pathogens and halotolerant microorganisms may develop, whereas other pathogens may sporulate a little. This may still be sufficient for infection [137,170]. |
| Extreme salinity (stage II) | 49.15–56.10 | 500–600 | Obligate halophytes can develop. Almost very little vegetation survives [172,173,174]. | Salt-tolerant pathogens can sporulate even if they experience significant growth retardation. Saprophytes may still develop [164,175]. |
| Seawater–salt lake | >50 | 600–800 | Only obligate halophytes may develop with great retardation [172,173,174]. | Salt-tolerant pathogens can develop and produce high levels of stress metabolites and stress hormones. Saprophytic and salt-tolerant microorganisms experience severe growth retardation [166]. |
| Salt lake | >50 | >800 | The plant cannot grow [176]. | Most salt-tolerant pathogens cannot survive. Very few saprophytes show mycelial growth. Endophytes may develop. Tolerant pathogens have an infectious capacity. Mutant fungal strains may develop [176,177,178]. |
| Salinity (Leaf/Root) and Pathogen Stress on Wheat | Effect on Crop Plants and Improvement Strategies ** | References |
|---|---|---|
| Root (0–150 mmol L−1 NaCl) and Fusarium pseudograminearum | Trichoderma longibrachiatum (TG1) induced defenses in wheat seedlings under salinity and Fusarium pseudograminearum (Fg), a fungal agent for wheat crown rot, infection. | [72] |
| Root (200 mmol L−1 NaCl) and Pyricularia oryzae | Biocontrol agents (BCAs), such as Pseudomonas stutzeri AN10 and Trichoderma harzianum Th56, helped wheat plants infected with the less aggressive isolate Pyricularia oryzae under 200 mmol L−1 NaCl stress. BCAs did not enhance plant tolerance to high salt stress but improved the performance of salt-stressed wheat plants. ** The status of highly aggressive isolates remains controversial. | [46] |
| Aspergillus niger | Different priming treatments—viz., hydropriming, osmotic priming, halopriming, and hormonal priming techniques in wheat induced resistance against Aspergillus niger via improving the protein, proline, and sugar contents. Thaumatin-like protein (TLP), chitinase, and β-1,3-glucanase genes were highly expressed in all the primed plants. | [196] |
| Salinity and pathogen stress | Leymus mollis (Triticeae; Poaceae), a useful genetic resource for wheat (Triticum aestivum L.) breeding, synthesized high levels of allene oxide cyclase involved in the biosynthesis of jasmonic acid under salt stress. ** Gene transfer could be used from this plant to generate both disease-resistant and salt-tolerant wheat plants. | [197] |
| Salinity and pathogen stress | Cakile maritima, a halophytic plant, tolerated the pathogenic attack of Alternaria alternate under 200 mmol L−1 NaCl conditions. ** However, this is not always the case if plants cannot tolerate the high level of salinity. | [198] |
| Abiotic and pathogen stress | miRNA-mediated modulation of AtP5CS1 gene expression under combined stress filled crucial gaps. | [199] |
| Abiotic and pathogen stress | Silicon and hydrogen sulfide improved the physiological resistance of wheat plants to drought stress and the pathogen Puccinia tritici infection. Drought stress increased the susceptibility of wheat plants to leaf rust pathogen infection. | [200] |
| Z. tritici stress on wheats | Trichoderma harzianum reduced pycnidia coverage of the pathogen in the susceptible cultivar. ** Salt-tolerant T. harzianum or other species could be used under both saline and pathogen conditions. | [201] |
| Salinity on wheat | Effective microorganisms (EMs) could be used to remediate the saline soils growing wheat. ** Promising EM species controlling Z. tritici could be very useful to improve the conditions of wheat under the combined stress. | [53] |
| Z. tritici stress on wheat | Thinopyrum elongatum is suggested for use since it contains important genetic resources. It carries genes resistant to Fusarium, Zymoseptoria, and rust diseases as well as tolerance genes to soil salinity, low temperatures, water logging, etc., and some quality traits for bakery. | [202] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baran, B.; Ölmez, F.; Çapa, B.; Dikilitas, M. Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview. Life 2024, 14, 648. https://doi.org/10.3390/life14050648
Baran B, Ölmez F, Çapa B, Dikilitas M. Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview. Life. 2024; 14(5):648. https://doi.org/10.3390/life14050648
Chicago/Turabian StyleBaran, Behzat, Fatih Ölmez, Beritan Çapa, and Murat Dikilitas. 2024. "Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview" Life 14, no. 5: 648. https://doi.org/10.3390/life14050648
APA StyleBaran, B., Ölmez, F., Çapa, B., & Dikilitas, M. (2024). Defense Pathways of Wheat Plants Inoculated with Zymoseptoria tritici under NaCl Stress Conditions: An Overview. Life, 14(5), 648. https://doi.org/10.3390/life14050648