
Effects of Flavanone Derivatives on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
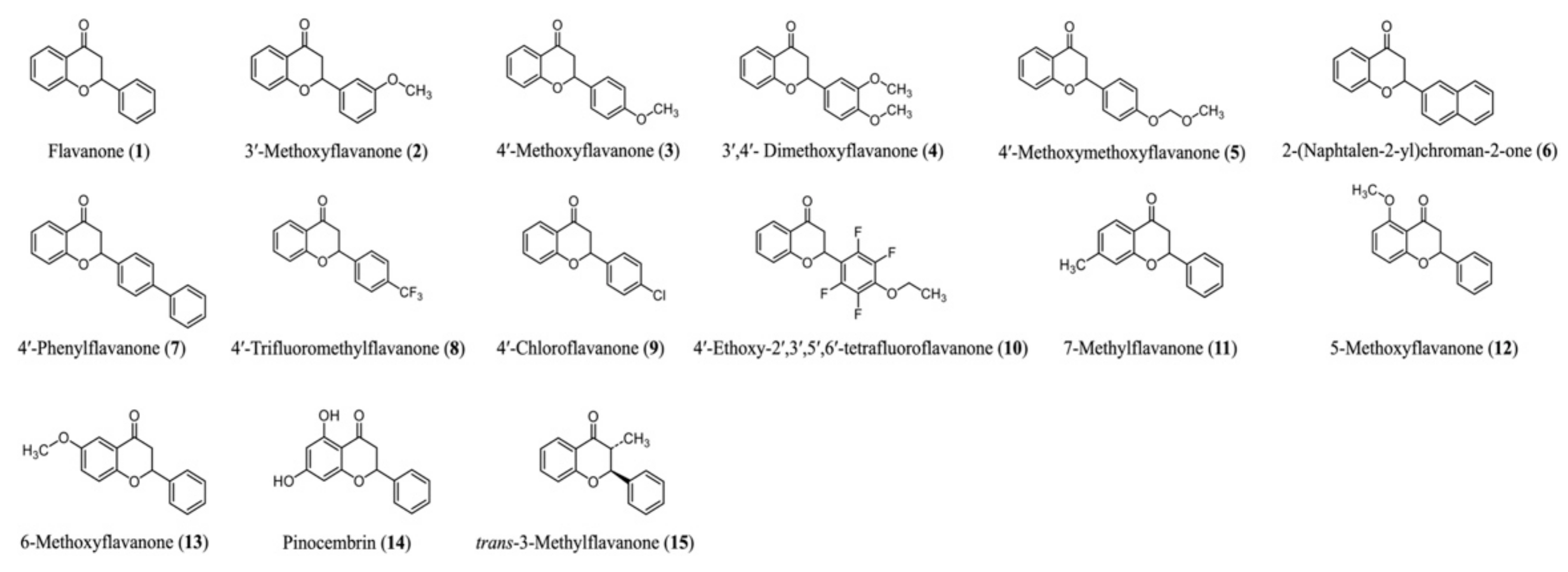
2.1. Flavanone Derivatives
2.2. Cell Culture and Adipocyte Differentiation
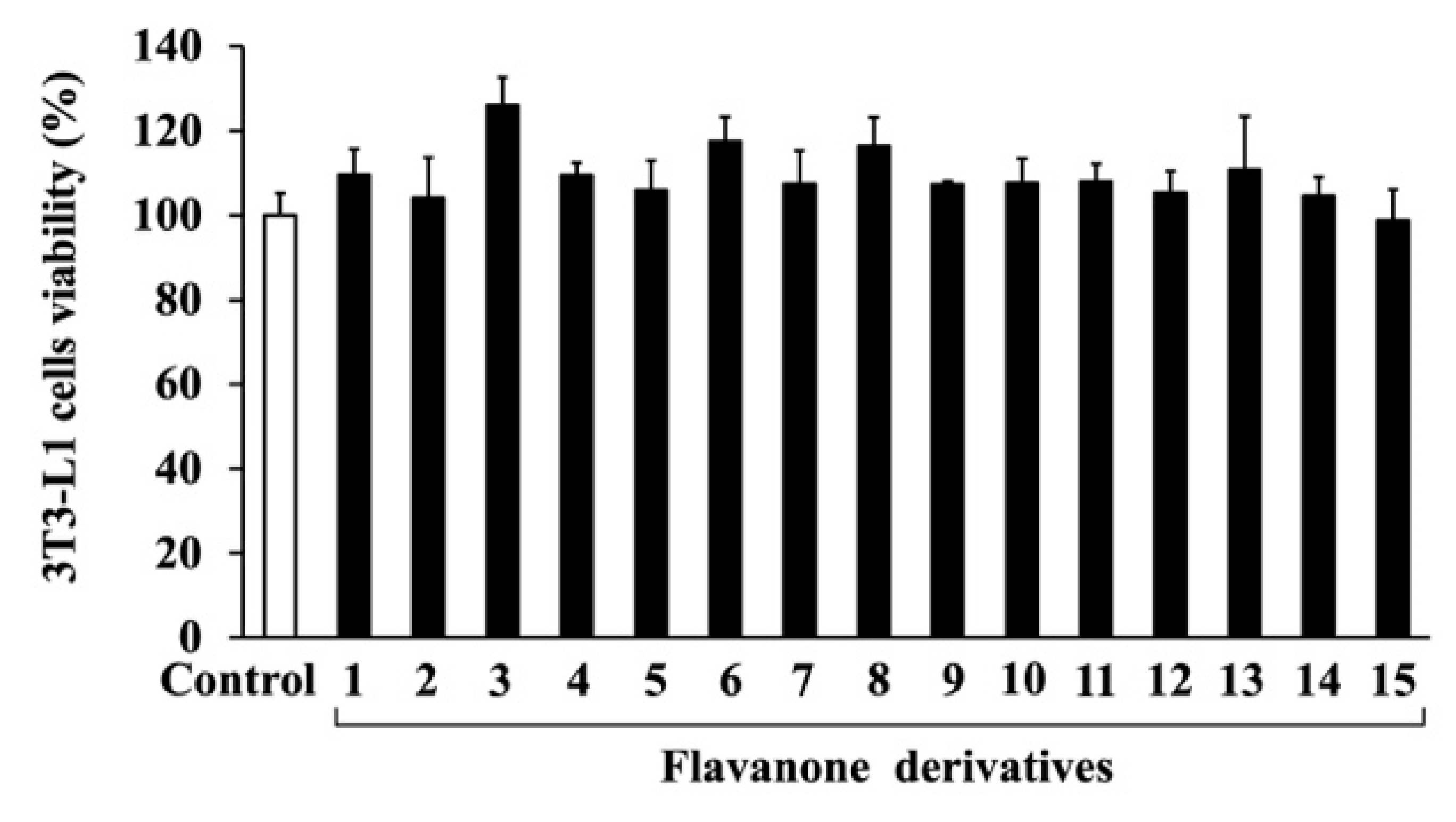
2.3. Cell Viability Measurement
2.4. Oil Red O Staining
2.5. Quantitative RT-PCR (qRT-PCR)
2.6. Plasmid Constructs, Cell Transfection, and Mammalian Two-Hybrid System
2.7. Cell Proliferation Assay
2.8. Statistical Analysis
3. Results
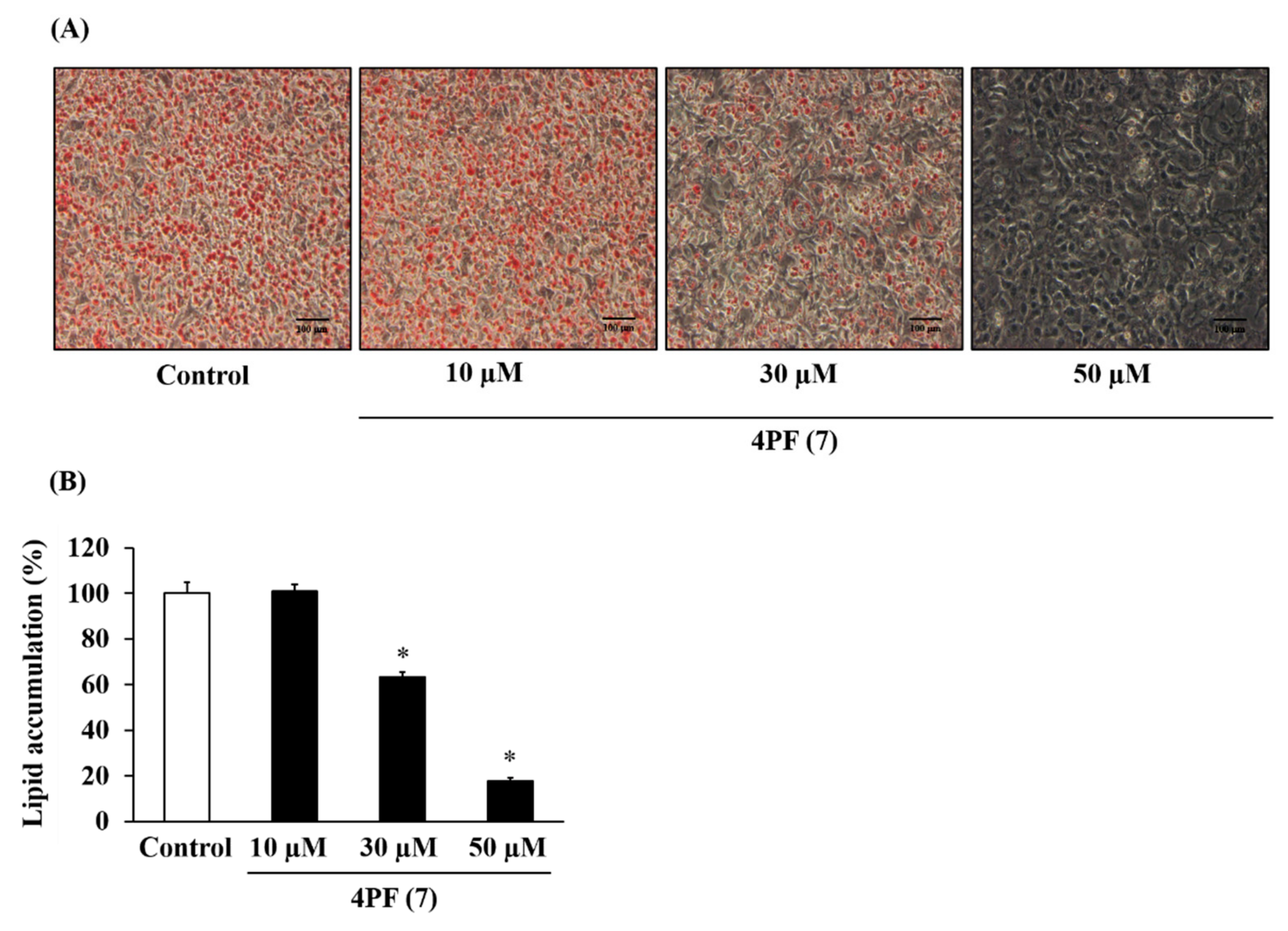
3.1. Effects of Flavanone Derivatives on Lipid Accumulation in 3T3-L1 Cells
3.2. Effect of 4PF (7) on Gene Expression During Adipocyte Differentiation
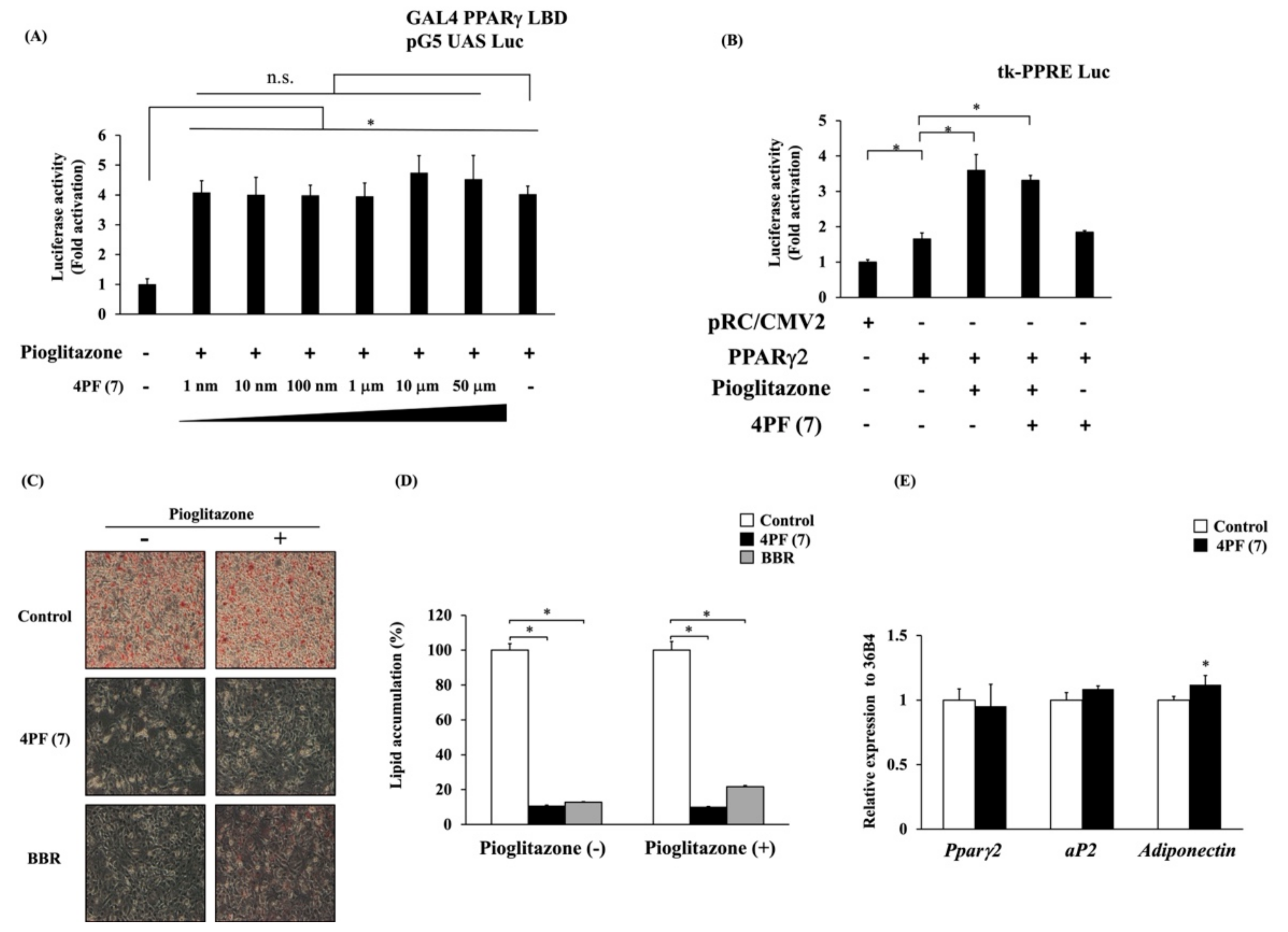
3.3. Effect of 4PF (7) on the Ligand-Dependent Activation and Gene Expression of PPARγ
3.4. Effect of 4PF (7) on Mitotic Clonal Expansion (MCE) During Adipocyte Differentiation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Formiguera, X.; Cantón, A. Obesity: Epidemiology and clinical aspects. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 1125–1146. [Google Scholar] [CrossRef] [PubMed]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, T.I.A. From fat cells through an obesity theory. Eur. J. Clin. Nutr. 2018, 72, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Parelee, S.D.; Mori, H.; Lentz, S.I.; MacDougald, O.A. Quantifying size and number of adipocytes in adipose tissue. Methods Enzymol. 2014, 537, 93–122. [Google Scholar]
- Nobushi, Y.; Hamada, Y.; Yasukawa, K. Inhibitory effects of the edible mushroom Flammulina velutipes on lipid accumulation in 3T3-L1 cells. J. Pharm. Nutr. Sci. 2013, 3, 222–227. [Google Scholar] [CrossRef]
- Kraus, N.A.; Ehebauer, F.; Zapp, B.; Rudolphi, B.; Kraus, B.J.; Kraus, D. Quantitative assessment of adipocyte differentiation in cell culture. Adipocyte 2016, 5, 351–358. [Google Scholar] [CrossRef]
- Nobushi, Y.; Oikawa, N.; Uchiyama, T.; Ohhara, H.; Kishikawa, Y. Inhibitory effects of Burchellin derivatives on lipid accumulation in 3T3-L1 cells. J. Jpn. Mibyou Assoc. 2020, 6, 13–19. [Google Scholar]
- Nobushi, Y.; Saito, H.; Miyairi, S.; Uchiyama, T.; Kishikawa, Y. Inhibitory effects of indirubin-3′-oxime derivatives on lipid accumulation in 3T3-L1 cells. Biol. Pharm. Bull. 2020, 43, 503–508. [Google Scholar] [CrossRef]
- Nobushi, Y.; Wada, T.; Koike, Y.; Kaneko, H.; Shimba, S.; Uchiyama, T.; Kishikawa, Y. Inhibitory effects of hydrolysable tannins on lipid accumulation in 3T3-L1 cells. Biol. Pharm. Bull. 2022, 45, 1458–1465. [Google Scholar] [CrossRef]
- Iwashina, T. The structure and distribution of the flavonoids in plants. J. Plant Res. 2000, 113, 287–299. [Google Scholar] [CrossRef]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a bioactive compound from medicinal plants and its therapeutic applications. BioMed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef]
- Hankowá, Z.; Hošek, J.; Kutil, Z.; Temml, V.; Landa, P.; Vaněk, T.; Schuster, D.; Dall’Acqua, S.; Cvačka, J.; Polanský, O.; et al. Anti-inflammatory activity of natural geranylated flavonoids: Cyclooxygenase and lipoxygenase inhibitory properties and proteomic analysis. J. Nat. Prod. 2017, 80, 999–1006. [Google Scholar]
- Chen, P.; Cao, Y.; Bao, B.; Zhang, L.; Ding, A. Antioxidant capacity of Typha angustifolia extracts and two active flavonoids. Pharm. Biol. 2017, 55, 1283–1288. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.J.; Guan, R.; Du, L.; Gao, J.; Fu, X.L. Strategies to target glucose metabolism in tumor microenvironment on cancer by flavonoids. Nutr. Cancer 2017, 69, 534–554. [Google Scholar] [CrossRef]
- Pouget, C.; Fagnere, C.; Basly, J.P.; Habrioux, G.; Chulia, A.J. New aromatase inhibitors. Synthesis and inhibitory activity of pyridinyl-substituted flavanone derivatives. Bioorg. Med. Chem. Lett. 2002, 12, 1059–1061. [Google Scholar] [CrossRef] [PubMed]
- Ankihiwala, M.D. Studies on flavonoids. Part-II. Synthesis and antimicrobial activity of 8-bromo-7-n-butoxy-6-nitro-flavones, -flavonols and -flavanones. J. Indian Chem. Soc. 1990, 67, 913–915. [Google Scholar]
- Wdowiak, K.; Walkowiak, J.; Pietrzak, R.; Bazan-Woźniak, A.; Cielecka-Piontek, J. Bioavailability of hesperidin and its aglycone hesperetin-compounds found in citrus fruits as a parameter conditioning the pro-health potential (neuroprotective and antidiabetic activity)-mini-review. Nutrients 2022, 14, 2647. [Google Scholar] [CrossRef]
- Assini, J.M.; Mulvihill, E.E.; Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Chhoker, S.S.; Sawyez, C.G.; Drangova, M.; Adams, A.C.; Kharitonenkov, A.; et al. Naringenin prevents obesity, hepatic steatosis, and glucose intolerance in male mice independent of fibroblast growth factor 21. Endocrinology 2015, 156, 2087–2102. [Google Scholar] [CrossRef]
- Dayarathne, L.A.; Ranaweera, S.S.; Natraj, P.; Rajan, P.; Lee, Y.J.; Han, C.H. Restoration of the adipogenic gene expression by naringenin and naringin in 3T3-L1 adipocytes. J. Vet. Sci. 2021, 22, e55. [Google Scholar] [CrossRef]
- Saito, T.; Abe, D.; Sekiya, K. Sakuranetin induces adipogenesis of 3T3-L1 cells through enhanced expression of PPARγ2. Biochem. Biophys. Res. Commun. 2008, 372, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Abe, D.; Sekiya, K. Flavanone exhibits PPARγ ligand activity and enhances differentiation of 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2009, 380, 281–285. [Google Scholar] [CrossRef]
- Oikawa, N.; Nobushi, Y.; Wada, T.; Sonoda, K.; Okazaki, Y.; Tsutsumi, S.; Park, Y.K.; Kurokawa, M.; Shimba, S.; Yasukawa, K. Inhibitory effects of compounds isolated from the dried branches and leaves of murta (Myrceugenia euosma) on lipid accumulation in 3T3-L1 cells. J. Nat. Med. 2016, 70, 502–509. [Google Scholar] [CrossRef]
- Miura, M.; Shigematsu, K.; Toriyama, M.; Motohashi, S. Convenient synthesis of flavanone derivatives via oxa-Michael addition using catalytic amount of aqueous cesium fluoride. Tetrahedron Lett. 2021, 85, 153480. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Kanno, Y.; Li, W.; Sasaki, T.; Zhang, X.; Wang, J.; Cheng, M.; Koike, K.; Nemoto, K.; Li, H. Identification of picrasidine C as a subtype-selective PPARα agonist. J. Nat. Med. 2016, 79, 3127–3133. [Google Scholar] [CrossRef]
- Shimba, S.; Wada, T.; Hara, S.; Tezuka, M. EPAS1 promotes adipose differentiation in 3T3-L1 cells. J. Biol. Chem. 2004, 279, 40946–40953. [Google Scholar] [CrossRef]
- Kim, J.Y.; Wall, E.V.D.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G.; et al. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Investig. 2007, 117, 2621–2637. [Google Scholar] [CrossRef]
- Carvalho, E.; Schellhorn, S.E.; Zabolotny, J.M.; Martin, S.; Tozzo, E.; Peroni, O.D.; Houseknecht, K.L.; Mundt, A.; James, D.E.; Kahn, B.B. GLUT4 overexpression or deficiency in adipocytes of transgenic mice alters the composition of GLUT4 vesicles and the subcellular localization of GLUT4 and insulin-responsive aminopeptidase. J. Biol. Chem. 2004, 279, 21598–21605. [Google Scholar] [CrossRef]
- Hishida, T.; Nishizaka, M.; Osada, S.; Imagawa, M. The role of C/EBPδ in the early stage of adipogenesis. Biochimie 2009, 91, 654–657. [Google Scholar] [CrossRef]
- Hallenborg, P.; Petersen, R.K.; Feddersen, S.; Sundekilde, U.; Hansen, J.B.; Blagoev, B.; Madsen, L.; Kristiansen, K. PPARγ ligand production is tightly linked to clonal expansion during initiation of adipocyte differentiation. J. Lipid Res. 2014, 55, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Baumann, A.; Burger, K.; Brandt, A.; Staltner, R.; Jung, F.; Rajcic, D.; Lorenzo Pisarello, M.J.; Bergheim, I. GW9662, a peroxisome proliferator-activated receptor gamma antagonist, attenuates the development of non-alcoholic fatty liver disease. Metabolism 2022, 133, 155233. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, T.; Han, Y.; Sheng, J.Z.; Jin, S.; Jin, M.W. Pentamethylquercetin improves adiponectin expression in differentiated 3T3-L1 cells via a mechanism that implicates PPARgamma together with TNF-alpha and IL-6. Molecules 2011, 16, 5754–5768. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Aci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Z.; Wu, N. Association between intake of flavanones and the overweight/obesity and central obesity in children and adolescents: A cross-sectional study from the NHANES database. Front Nutr. 2024, 11, 1430140. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Iwasa, M.; Marunaka, Y. Metabolic functions of flavonoids: From human epidemiology to molecular mechanism. Neuropeptides 2021, 88, 102163. [Google Scholar] [CrossRef]
- Kim, M.A.; Kang, K.; Lee, H.J.; Kim, M.; Kim, C.Y.; Nho, C.W. Apigenin isolated from Daphne genkwa Siebold et Zucc. inhibits 3T3-L1 preadipocyte differentiation through a modulation of mitotic clonal expansion. Life Sci. 2014, 101, 64–72. [Google Scholar] [CrossRef]
- Kanda, K.; Nishi, K.; Kadota, A.; Nishimoto, S.; Liu, M.C.; Sugahara, T. Nobiletin suppresses adipocyte differentiation of 3T3-L1 cells by an insulin and IBMX mixture induction. Biochim. Biophys. Acta 2012, 1820, 461–468. [Google Scholar] [CrossRef]
- Kim, S.C.; Kim, Y.H.; Son, S.W.; Moon, E.Y.; Pyo, S.; Um, S.H. Fisetin induces Sirt1 expression while inhibiting early adipogenesis in 3T3-L1 cells. Biochem. Biophys. Res. Commun. 2015, 467, 638–644. [Google Scholar] [CrossRef]
- Qadi, H.H.; Bendary, M.A.; Almaghrabi, S.Y.; Zaher, M.A.F.; Karami, M.M.; Alsehli, A.M.; Babateen, O.; Arbaeen, A.F.; Burzangi, A.S.; Bazuhair, M.A. Exploring the therapeutic potential of apigenin in obesity-associated fibrinolytic dysfunction: Insights from an animal study. Cureus 2023, 15, e40943. [Google Scholar] [CrossRef]
- Lee, Y.S.; Cha, B.Y.; Choi, S.S.; Choi, B.K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Nobiletin improves obesity and insulin resistance in high-fat diet-induced obese mice. J. Nutr. Biochem. 2013, 24, 156–162. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, H.; Deng, R.; Wang, N.; Zhang, Y.; Wang, Y.; Liu, Y.; Li, F.; Wang, X.; Zhou, L. Berberine suppresses adipocyte differentiation via decreasing CREB transcriptional activity. PLoS ONE 2015, 10, e0125667. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; Mckeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell. 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, M.; Cai, W.; Yu, L.; Feng, L.; Zhang, L.; Zang, Q.; Wang, Y.; Wang, D.; Chen, H.; et al. Dietary component isorhamnetin is a PPARγ antagonist and ameliorates metabolic disorders induced by diet or leptin deficiency. Sci. Rep. 2016, 6, 19288. [Google Scholar] [CrossRef]
- Choi, E.J.; Jung, J.Y.; Kim, G.H. Genistein inhibits the proliferation and differentiation of MCF-7 and 3T3-L1 cells via the regulation of ERalpha expression and induction of apoptosis. Exp. Ther. Med. 2014, 8, 454–458. [Google Scholar] [CrossRef]
- Feng, X.; Weng, D.; Zhou, F.; Owen, Y.D.; Qin, H.; Zhao, J.; Wen, Y.; Huang, Y.; Chen, J.; Fu, H.; et al. Activation of PPARγ by a natural flavonoid modulator, apigenin ameliorates obesity-related inflammation via regulation of macrophage polarization. EBioMedicine 2016, 9, 61–76. [Google Scholar] [CrossRef]
- Cao, Z.; Umek, R.M.; Mcknight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef]
- MacDougald, O.A.; Lane, M.D. Adipocyte differentiation. When precursors are also regulators. Curr. Biol. 1995, 5, 618–621. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Y.; Li, S.; Yang, Y.; Dai, Z.; Wang, F.; Wu, Z.; Tso, P.; Wu, G. E2F1 regulates adipocyte differentiation and adipogenesis by activating ICAT. Cells 2020, 9, 1024. [Google Scholar] [CrossRef] [PubMed]
- Abdesselem, H.; Madani, A.; Hani, A.; Al-Noubi, M.; Goswami, N.; Hamidane, H.B.; Billing, A.M.; Pasquier, J.; Bonkowski, M.S.; Halabi, N.; et al. SIRT1 limits adipocyte hyperplasia through c-Myc inhibition. J. Biol. Chem. 2016, 291, 2119–2135. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Kogami, Y.; Nakamura, S.; Sugiyama, T.; Ueno, T.; Yoshikawa, M. Structural requirements of flavonoids for the adipogenesis of 3T3-L1 cells. Bioorg. Med. Chem. 2011, 19, 2835–2841. [Google Scholar] [CrossRef]
- Miyahisa, I.; Kaneko, M.; Funa, N.; Kawasaki, H.; Kojima, H.; Ohnishi, Y.; Horinouchi, S. Efficient production of (2S)-flavanones by Escherichia coli containing an artificial biosynthetic gene cluster. Appl. Microbiol. Biotechnol. 2005, 68, 498–504. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | (5′–3′ Sequence) |
|---|---|---|
| Ppary2 | Forward | GCTGTTATGGGTGAAACTCTG |
| Reverse | ATAATAAGGTGGAGATGCAGG | |
| C/ebpα | Forward | TGGACAAGAACAGCAACGAG |
| Reverse | TCACTGGTCAACTCCAGCAC | |
| aP2 | Forward | ATGAAATCACCGCAGACGACAGGA |
| Reverse | TGTGGTCGACTTTCCATCCCACTT | |
| Adiponectin | Forward | AAGGACAAGGCCGTTCTCT |
| Reverse | TATGGGTAGTTGCAGTCAGTTGG | |
| Glut4 | Forward | GCTTTGTGGCCTTCTTTGAG |
| Reverse | CGGCAAATAGAAGGAAGACG | |
| C/ebpδ | Forward | CCCCAAAGCTATGTGCCTTTC |
| Reverse | CCTGGAGGGTTTGTGTTTTC | |
| C/ebpβ | Forward | GGTTTCGGGACTTGATGCA |
| Reverse | CAACAACCCCGCAGGAAC | |
| p16 | Forward | GTCGCAGGTTCTTGGTCACT |
| Reverse | CGAATCTGCACCGTAGTTGA | |
| p21 | Forward | CGGTGGAACTTTGACTTCGT |
| Reverse | CAGGGCAGAGGAAGTACTGG | |
| p27 | Forward | AGCAGTGTCCAGGGATGAGGAA |
| Reverse | TTCTTGGGCGTCTGCTCCACAG | |
| p57 | Forward | AACGTCTGAGATGAGTTAGTTTAGAGG |
| Reverse | AAGCCCAGAGTTCTTCCATCGT | |
| Pcna | Forward | CCACATTGGAGATGCTGTTG |
| Reverse | CAGTGGAGTGGCTTTTGTGA | |
| cMyc | Forward | TCGCTGCTGTCCTCCGAGTCC |
| Reverse | GGTTTGCCTCTTCTCCACAGAC | |
| CyclinA2 | Forward | TACCTGCCTTCACTCATTGCTGGA |
| Reverse | ATTGACTGTTGGGCATGTTGTGGC | |
| CyclinD1 | Forward | TGCTGCAAATGGAACTGCTTCTGG |
| Reverse | TACCATGGAGGGTGGGTTGGAAAT | |
| CyclinE | Forward | AAGCCCTCTGACCATTGTGTCC |
| Reverse | CTAAGCAGCCAACATCCAGGAC | |
| Cdk2 | Forward | CCCCAGAACCTGCTTATCAA |
| Reverse | TGTGTTCCCCACACACTTA | |
| Cdk4 | Forward | TTTGTGGCCCTCAAGAGTGTGAGA |
| Reverse | TCCTTAACAAGGCCACCTCACGAA | |
| Cdk6 | Forward | TAAAGCTGGCTGACTTTGGCCTTG |
| Reverse | TCTGCAAAGATGCAACCGACACTC | |
| 36b4 | Forward | AAGCGCGTCCTGGCATTGTCT |
| Reverse | CCGCAGGGGCAGCAGTGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nobushi, Y.; Wada, T.; Miura, M.; Onoda, R.; Ishiwata, R.; Oikawa, N.; Shigematsu, K.; Nakakita, T.; Toriyama, M.; Shimba, S.; et al. Effects of Flavanone Derivatives on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells. Life 2024, 14, 1446. https://doi.org/10.3390/life14111446
Nobushi Y, Wada T, Miura M, Onoda R, Ishiwata R, Oikawa N, Shigematsu K, Nakakita T, Toriyama M, Shimba S, et al. Effects of Flavanone Derivatives on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells. Life. 2024; 14(11):1446. https://doi.org/10.3390/life14111446
Chicago/Turabian StyleNobushi, Yasuhito, Taira Wada, Motofumi Miura, Rikuto Onoda, Ryuta Ishiwata, Naoki Oikawa, Karin Shigematsu, Toshinori Nakakita, Masaharu Toriyama, Shigeki Shimba, and et al. 2024. "Effects of Flavanone Derivatives on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells" Life 14, no. 11: 1446. https://doi.org/10.3390/life14111446
APA StyleNobushi, Y., Wada, T., Miura, M., Onoda, R., Ishiwata, R., Oikawa, N., Shigematsu, K., Nakakita, T., Toriyama, M., Shimba, S., & Kishikawa, Y. (2024). Effects of Flavanone Derivatives on Adipocyte Differentiation and Lipid Accumulation in 3T3-L1 Cells. Life, 14(11), 1446. https://doi.org/10.3390/life14111446