
Cytogenetic Studies on Genoprotective Effect of Rosa damascena Mill. Hydrosol in Plant and Lymphocyte Test Systems

 , , and
, , and
Abstract
:
{kind=link}
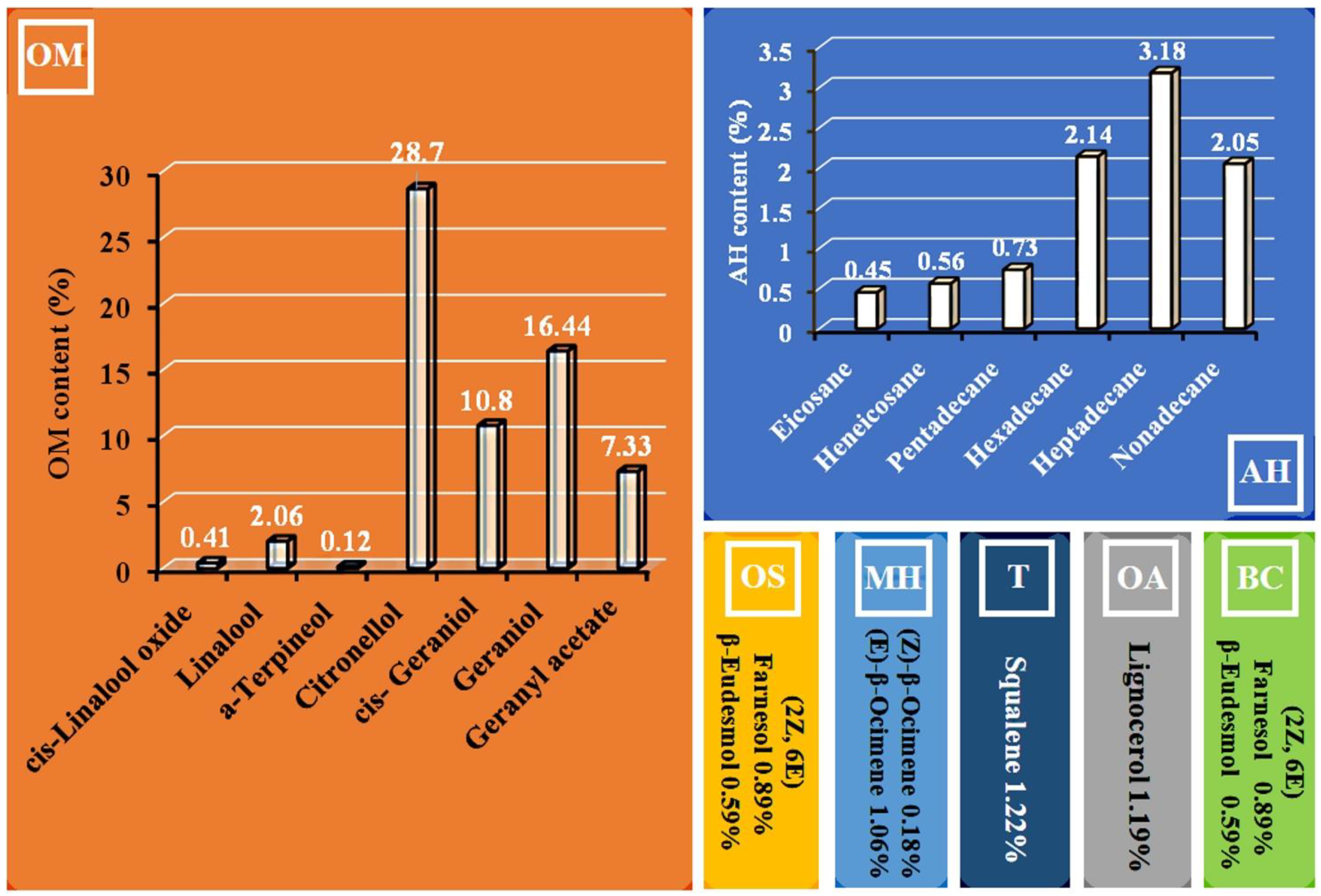{kind=link}
{kind=link}
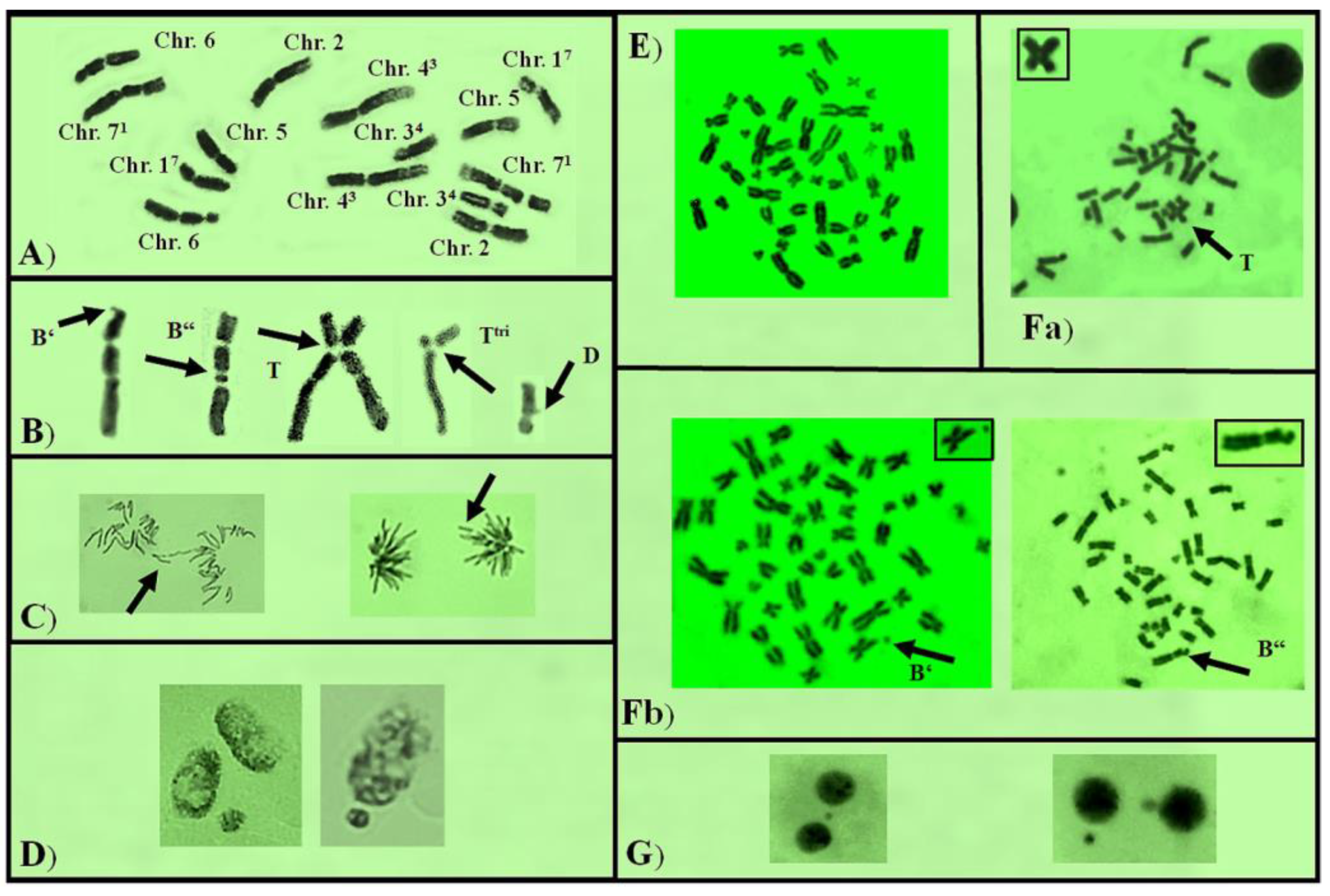{kind=link}
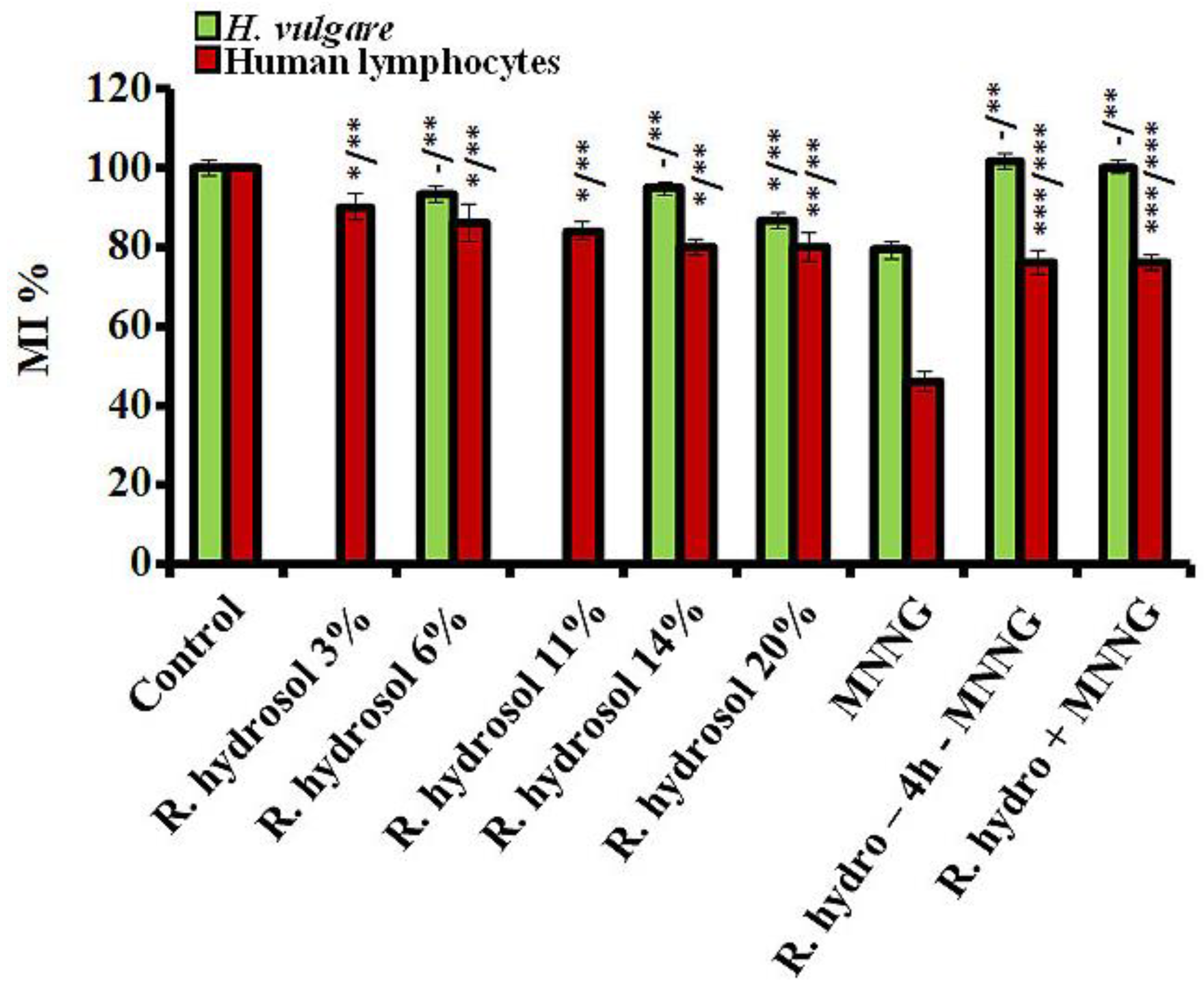{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
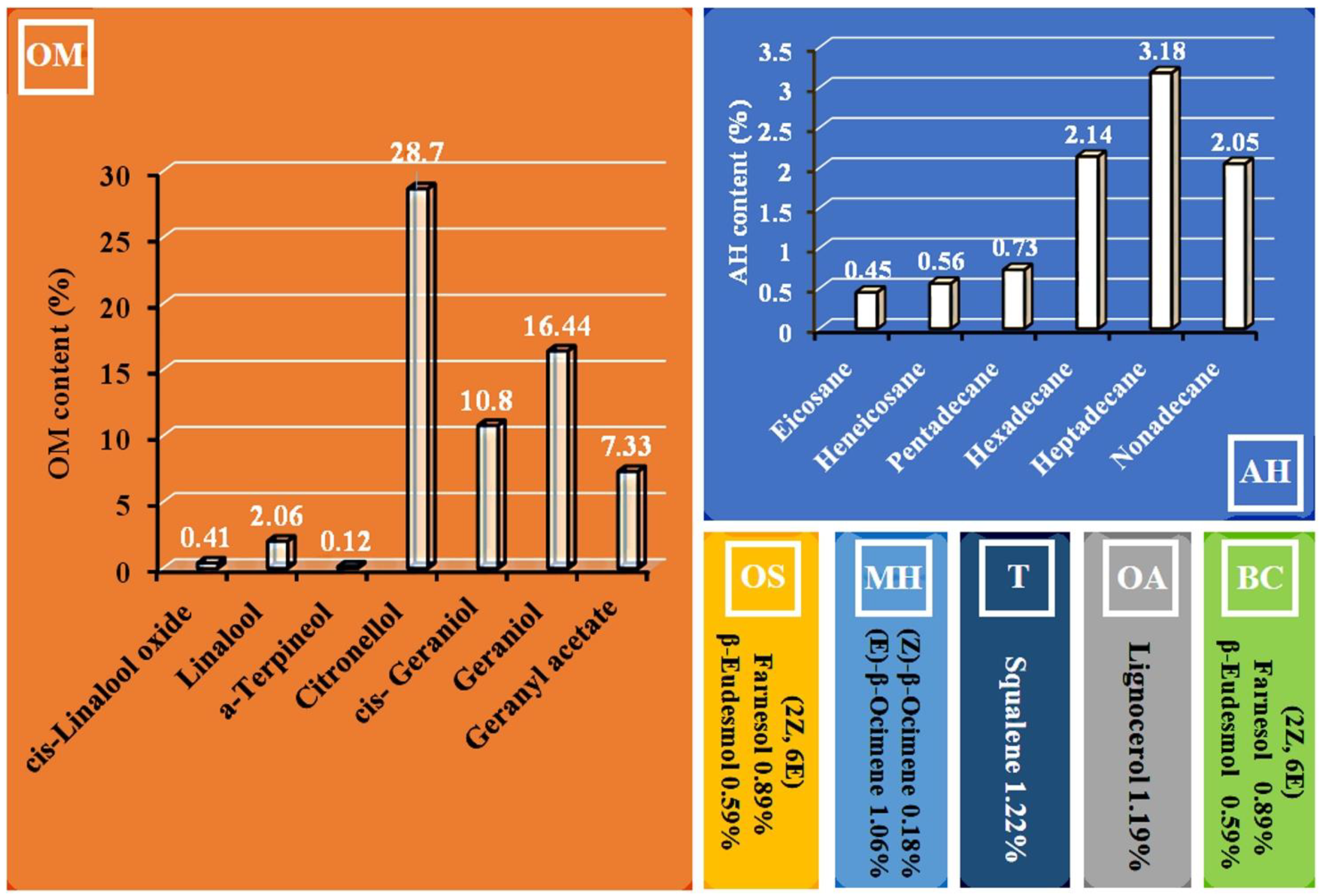
2.2. Plant Hydrosol
2.3. Test Systems
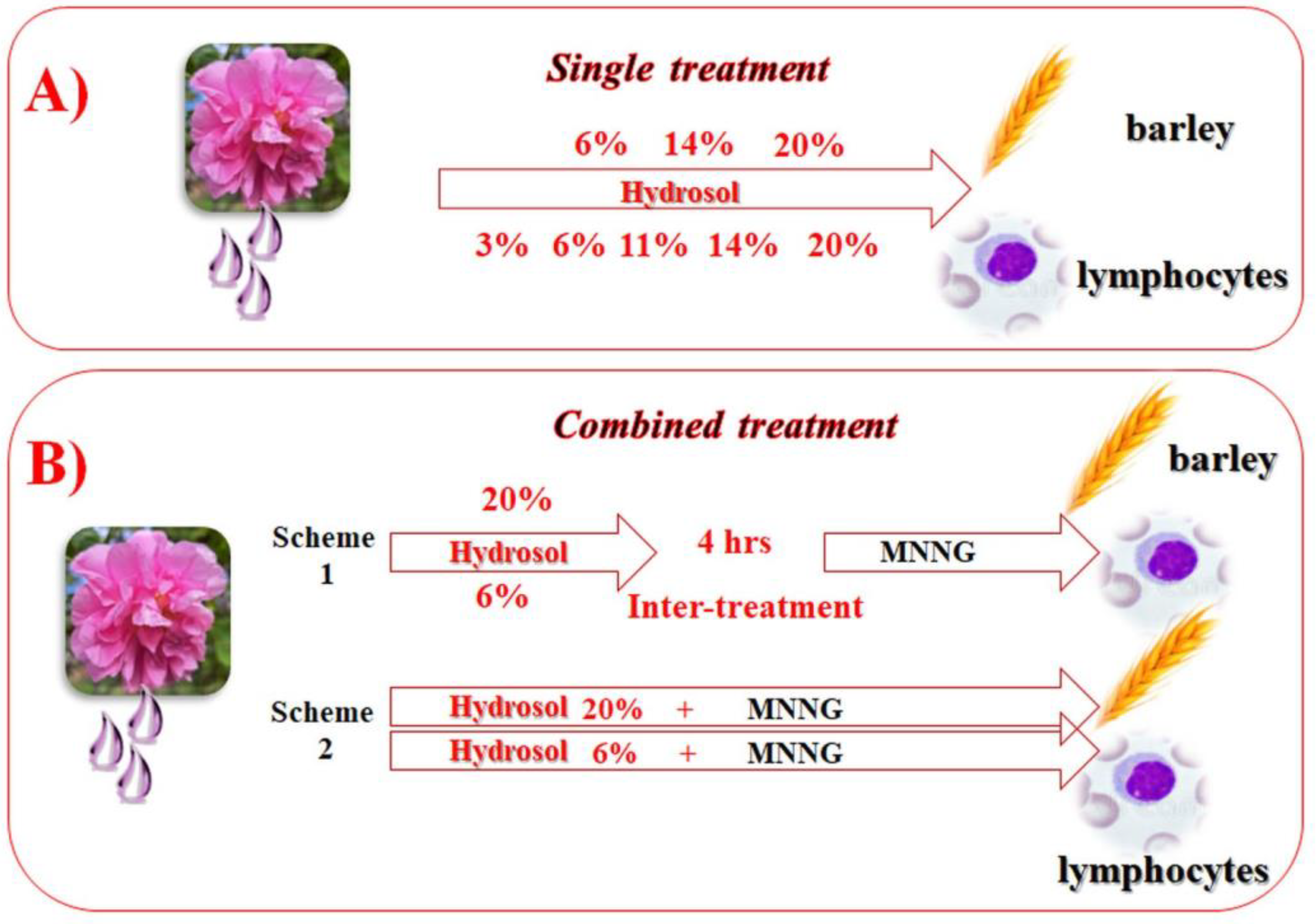
2.4. Experimental Schemes
2.5. Endpoints
2.6. Statistical Analysis
3. Results
3.1. Cytotoxicity/Anticytotoxicity
3.1.1. Cytotoxicity
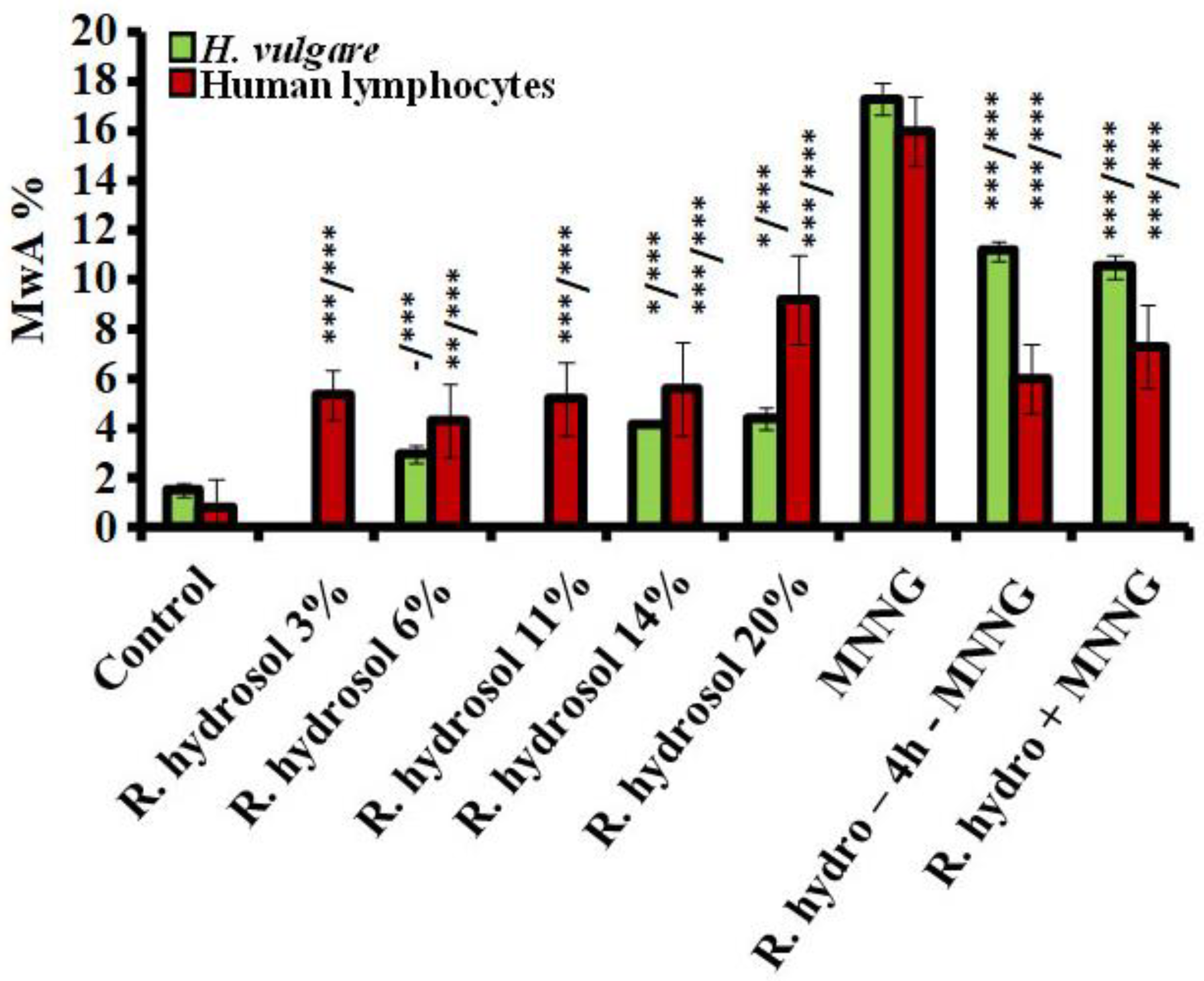
3.1.2. Anticytotoxicity
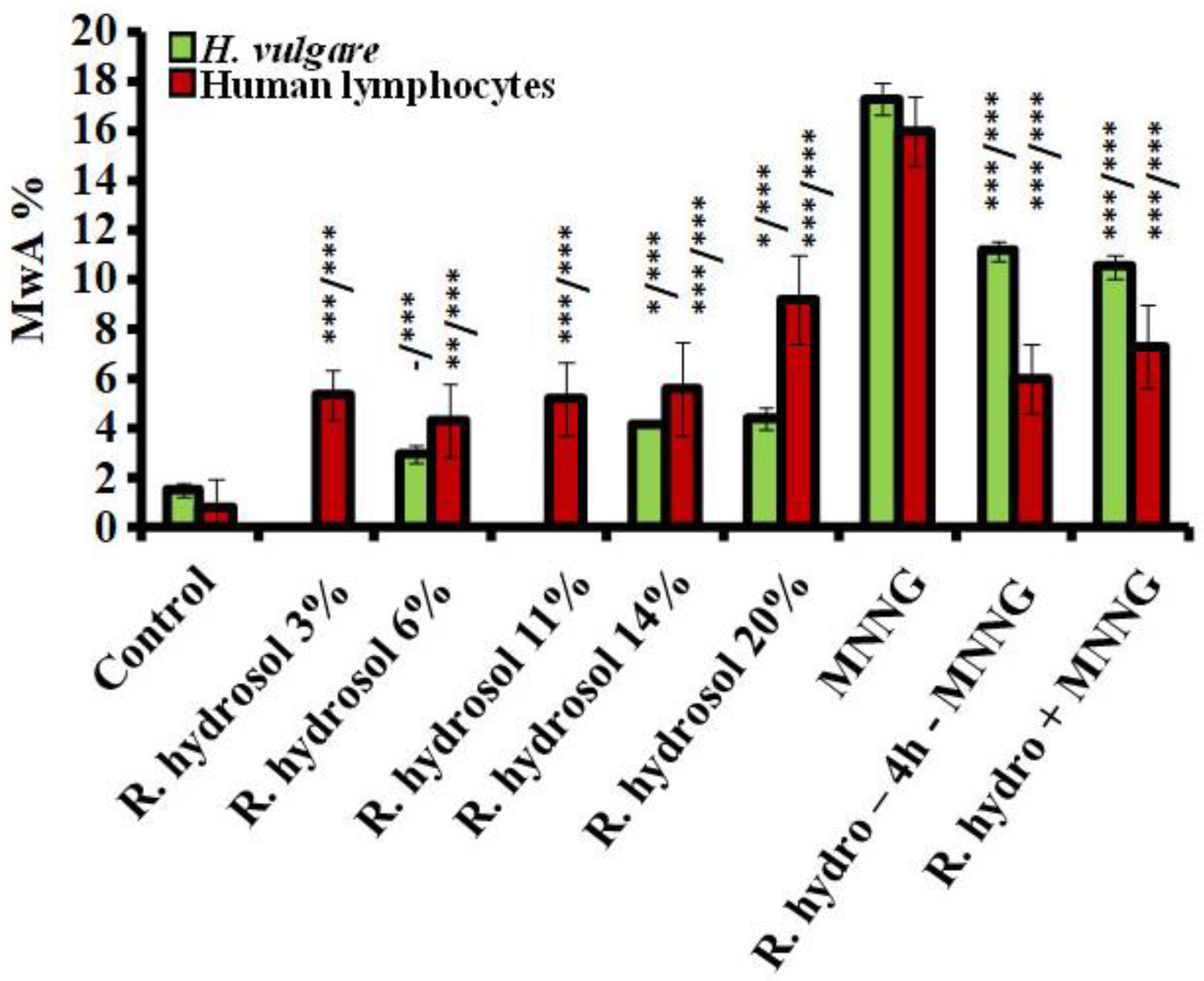
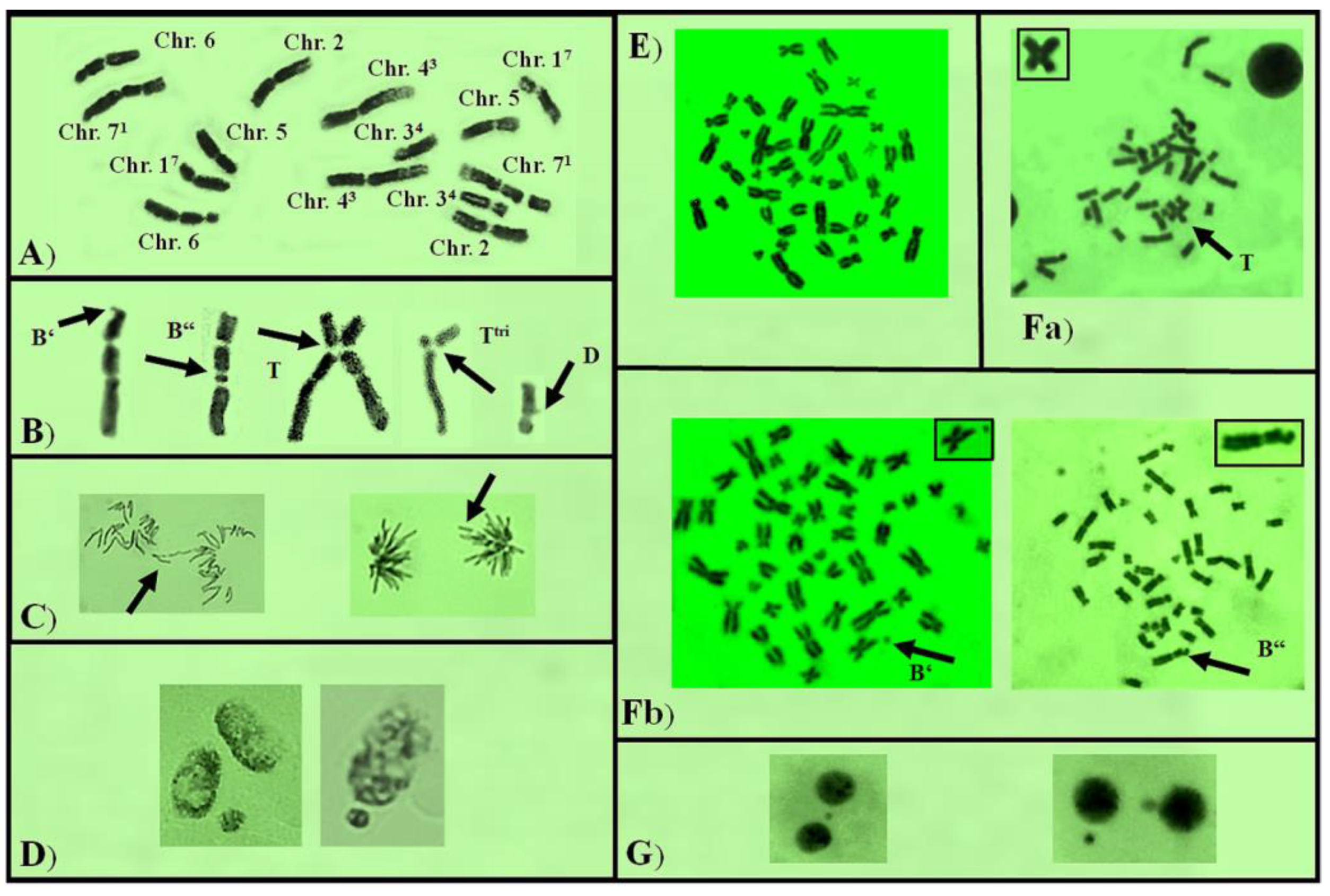
3.2. Genotoxicity/Antigenotoxicity
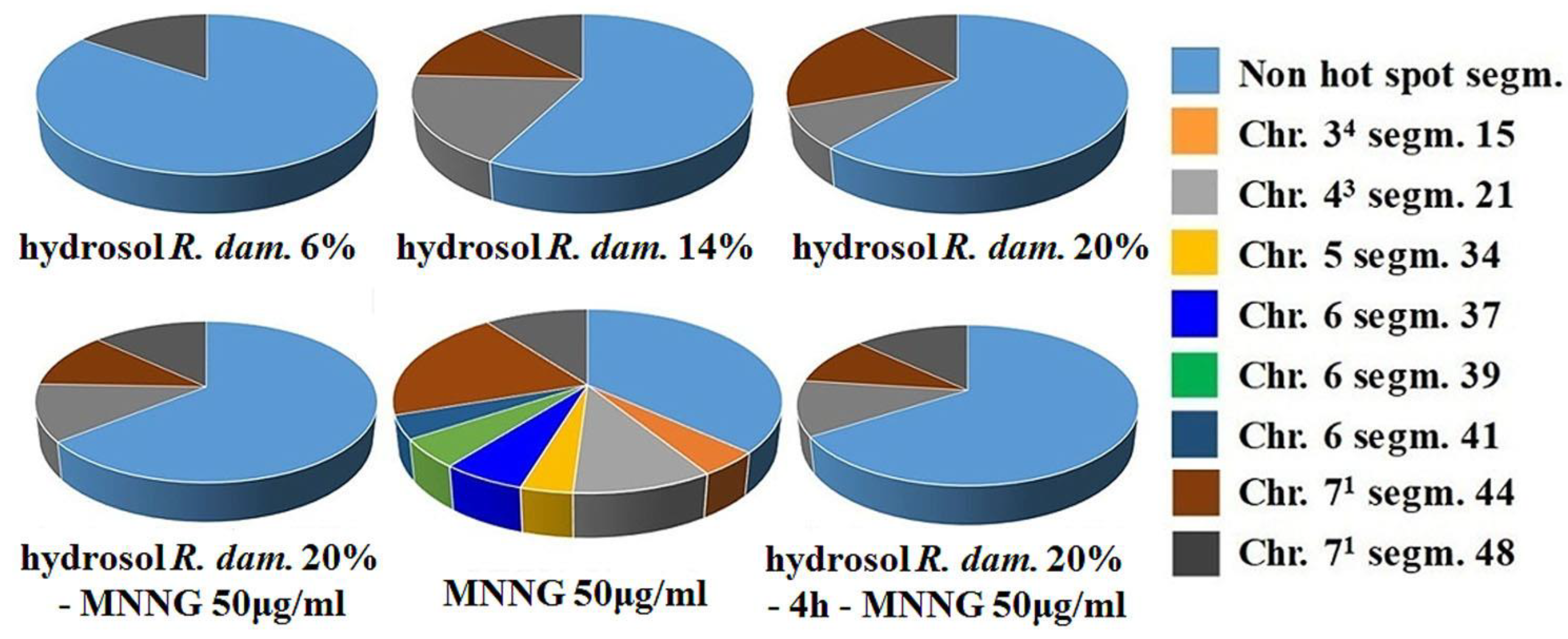
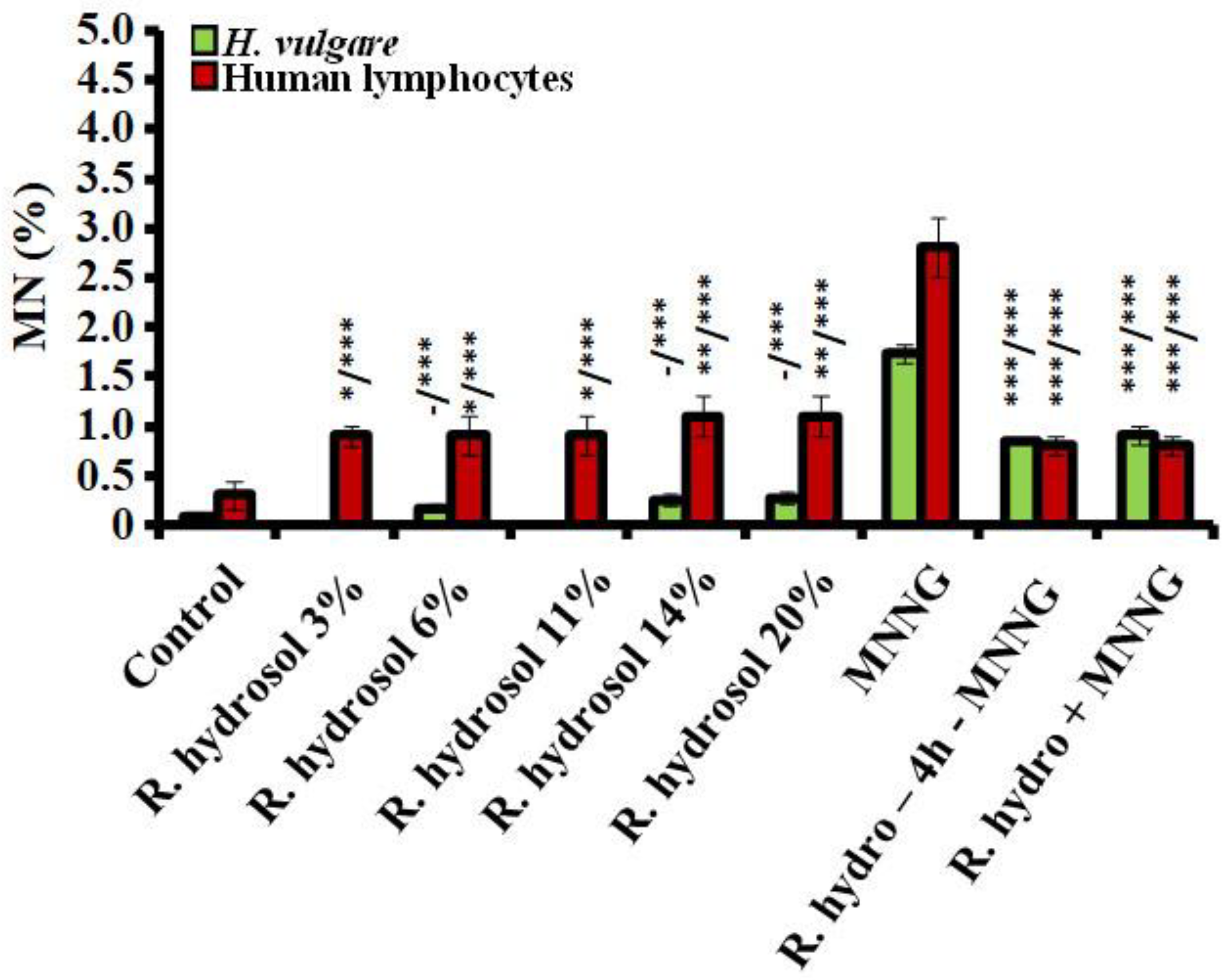
3.2.1. Genotoxicity
3.2.2. Antigenotoxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mezzoug, N.; Abrini, J.; Serano, A.M.; Alonso-Moraga, A.; Idaomar, M. Study on antigenotoxic effects of Moroccan medicinal plants and spices using the white/white + somatic assay in Drosophila. Afr. J. Trad. CAM 2006, 3, 22–31. [Google Scholar]
- Mosaffa, F.; Behravan, J.; Karimi, G.; Iranshahi, M. Antigenotoxic effects of Satureja hortensis L. on rat lymphocytes exposed to oxidative stress. Arch. Pharm. Res. 2006, 29, 159–164. [Google Scholar] [CrossRef]
- Boukhris, M.; Hadricha, F.; Chtouroua, H.; Dhouiba, A.; Bouaziz, M.; Sayadi, S. Chemical composition, biological activities and DNA damage protective effect of Pelargonium graveolens L’Hér. essential oils at different phenological stages. Ind. Crop. Prod. 2015, 74, 600–606. [Google Scholar] [CrossRef]
- Lopez-Romero, D.L.; Izquierdo-Vega, J.A.; Morales-González, J.A.; Madrigal-Bujaidar, E.; Chamorro-Cevallos, G.; Sánchez-Gutiérrez, M.; Betanzos-Cabrera, G.; Alvarez-Gonzalez, I.; Morales-González, A.; Madrigal-Santillán, E. Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin. Nutrients 2018, 10, 1954. [Google Scholar] [CrossRef]
- Sabahi, Z.; Soltani, F.; Moein, M. Insight into DNA protection ability of medicinal herbs and potential mechanisms in hydrogen peroxide damages model. Asian Pac. J. Trop Biomed. 2018, 8, 120–129. [Google Scholar] [CrossRef]
- Ho, Y.S.; Wu, J.-Y.; Chang, C.Y. A New Natural Antioxidant Biomaterial from Cinnamomum osmophloeum Kanehira Leaves Represses Melanogenesis and Protects against DNA Damage. Antioxidants 2019, 8, 474. [Google Scholar] [CrossRef]
- Aly, F.A.E.; Farghaly, A.A.; Salman, A.S.; Hassan, E.M.; Hassan, E.E.; Omara, E.A.A.; Salama, A.B. Protective Effect of Rosmarinus officinalis Essential Oil Against Genotoxic and Histopathological Alterations Induced by Paracetamol. Int. J. Pharmacol. 2019, 15, 686–695. [Google Scholar] [CrossRef]
- Mondal, S.; Bandyopadhyay, A. Photoprotective, antioxidant potential and DNA damage protection assay of leaf methanolic extract of Holoptelea integrifolia (Roxb) Planch and determination of some bioactive phenolic compounds by RP-HPLC. Biocatal. Agric. Biotechnol. 2023, 50, 102728. [Google Scholar] [CrossRef]
- Ulusoy, S.; Boşgelmez-Tinaz, G.; Seçilmiş-Canbay, H. Tocopherol, carotene, phenolic contents and antibacterial properties of rose essential oil, hydrosol and absolute. Curr. Microbiol. 2009, 59, 554–558. [Google Scholar] [CrossRef]
- Boskabady, M.H.; Shafei, M.N.; Saberi, Z.; Amini, S. Pharmacological effects of Rosa damascena. Iran. J. Basic Med. Sci. 2011, 14, 295–307. [Google Scholar]
- Hajhashemi, V.; Ghannadi, A.; Hajiloo, M. Analgesic and anti-inflammatory effects of Rosa damascena hydroalcoholic extract and its essential oil in animal models. Iran. J. Pharm. Res. 2010, 9, 163–168. [Google Scholar]
- Shafei, M.N.; Rakhshandah, H.; Boskabady, M.H. Antitussive effect of Rosa damascena in guinea pigs. Iran. J. Pharm. Res. 2010, 2, 231–234. [Google Scholar]
- Mileva, M.; Krumova, E.; Stavreva, J.; Kostadinova, M. Chemical compounds, in vitro antioxidant and antifungal activities of some plant essential oils belonging to Rosaceae family. Compt. Rend. Acad. Bulg. Sci. 2014, 67, 1363–1368. [Google Scholar]
- Abdel-Hameed, E.S.; Bazaid, S.A.; Hagag, H.A. Chemical characterization of Rosa damascena Miller var. trigintipetala Dieck essential oil and its in vitro genotoxic and cytotoxic properties. J. Essent. Oil Res. 2016, 28, 121–129. [Google Scholar] [CrossRef]
- Hamedi, S.; Shomali, T.; Haghighat, A. Rosa damascena Mill. Essential Oil Has Protective Effect Against Testicular Damage in Diabetic Rats. J. Diet Suppl. 2017, 15, 311–317. [Google Scholar] [CrossRef]
- Kurkcuoglu, M.; Baser, K.N.C.; Aĸterian, S.G.; Fidan, H.N.; Stoyanova, A.S. Chemical composition, sensory evaluation and antimicrobial activity of Taif rose (Rosa damascena Mill.) essential oils. Bulg. Chem. Commun. 2020, 52, 460–466. [Google Scholar]
- Hagag, H.A.; Bazaid, S.A.; Abdel-Hameed, E.S.; Salman, M. Cytogenetic, cytotoxic and GC–MS studies on concrete and absolute oils from Taif rose, Saudi Arabia. Cytotechnology 2014, 66, 913–923. [Google Scholar] [CrossRef]
- Labadie, C.; Ginies, C.; Guinebretiere, M.; Renard, C.M.; Cerutti, C.; Carlin, F. Hydrosols of orange blossom (Citrus aurantium), and rose flower (Rosa damascena and Rosa centifolia) support the growth of a heterogeneous spoilage microbiota. Food Res. Int. 2015, 76, 576–586. [Google Scholar] [CrossRef]
- Rajeswara, R.B.R. Hydrosols and water-soluble essential oils: Medicinal and biological properties. Recent progress in medicinal plants. Essent. Oils I 2013, 36, 119e140. [Google Scholar]
- Eikani, M.M.; Golmohammad, F.; Rowshanzamir, S.; Mirza, M. Recovery of water-soluble constituents of rose oil using simultaneous distillation-extraction. Flavour Fragr. J. 2005, 20, 555–558. [Google Scholar] [CrossRef]
- Moein, M.; Zarshenas, M.M.; Delnavaz, S. Chemical composition analysis of rose water samples from Iran. Pharm. Biol. 2014, 52, 1358–1361. [Google Scholar] [CrossRef]
- Georgieva, A.; Dobreva, A.; Tzvetanova, E.; Alexandrova, A.; Mileva, M. Comparative Study of Phytochemical Profiles and Antioxidant Properties of Hydrosols from Bulgarian Rosa alba L. and Rosa damascena Mill. J. Essent. Oil Bear. Plant 2019, 22, 1362–1371. [Google Scholar] [CrossRef]
- Sharma, D.; Jain, O. Preparation of eco-friendly cosmetics and medicines by extraction of valuable essential oils and waters from flora. J. Appl. Chem. 2013, 2, 63–65. [Google Scholar]
- Mahboubi, M. Rosa damascena as holy ancient herb with novel applications. J. Tradit Complement. Med. 2016, 6, 10–16. [Google Scholar] [CrossRef]
- Haghighi, M.; Tehranifar, A.; Nikbakht, A.; Kafi, M. Research and current profile of Iranian production of Damask rose. ActaHort ISHS 2008, 769, 449–455. [Google Scholar] [CrossRef]
- Osama, M.; Ikram, R.; Sarfaraz, S.; Ahmed, S.; Iqbal, A. Screening of water distilled Rosa damascena Mill. flowers as hematopoietic agent in an animal model. Pak J. Pharm. Sci. 2020, 33, 103–107. [Google Scholar]
- Demirbolat, İ.; Ekinci, C.; Nuhoğlu, F.; Kartal, M.; Yıldız, P.; Özgün, M.; Geçer, M.Ö. Effects of Orally Consumed Rosa damascena Mill. Effect of orally consumed Hydrosol on Hematology, Clinical Chemistry, Lens Enzymatic Activity, and Lens Pathology in Streptozotocin-Induced Diabetic Rats. Molecules 2019, 24, 4069. [Google Scholar] [CrossRef]
- Ozkan, G.; Sagdic, O.; Baydar, N.G.; Baydar, H. Note: Antioxidant and antibacterial activities of Rosa damascena flower extracts. Food Sci. Technol. Int. 2004, 10, 277–281. [Google Scholar] [CrossRef]
- Chaleshtori, F.S.; Saholi, M.; Chaleshtori, R.S. Chemical Composition, Antioxidant and Antibacterial Activity of Buniumpersicum, Eucalyptus globulus, and Rose Water on Multidrug-Resistant Listeria Species. J. Evid. Based Integr. Med. 2018, 23, 2515690X17751314. [Google Scholar] [CrossRef]
- Gerasimova, T.; Jovtchev, G.; Gateva, S.; Topashka-Ancheva, M.; Stankov, A.; Angelova, T.; Dobreva, A.; Mileva, M. Study on Cytotoxic and Genotoxic Potential of Bulgarian Rosa damascena Mill. and Rosa alba L. Hydrosols—In Vivo and In Vitro. Life 2022, 12, 1452. [Google Scholar] [CrossRef]
- Abidi, S.; Shaheen, N.; Azher, I.; Mahmood, Z.A. Photoprotective and antioxidant activities along with phytochemical investigation of rose water. Int. J. Pharm. Sci. Res. 2018, 9, 5320–5326. [Google Scholar] [CrossRef]
- Künzel, G.; Nicoloff, H. Further results on karyotype reconstruction in barley. Biol. Zentrbl. 1979, 98, 587–592. [Google Scholar]
- Evans, E. Handbook of Mutagenicity Test Procedures; Kilbey, N., Legator, M., Nicols, W., Ramel, C., Eds.; Elsevier Science Publishers BV: Amsterdam, The Netherlands, 1984; pp. 405–427. [Google Scholar]
- Jovtchev, G.; Stergios, M.; Schubert, I. A comparison of N-methyl-N-nitrosourea-induced chromatid aberrations and micronuclei in barley meristems using FISH techniques. Mutat. Res. Gen. Toxicol. Environ. Mutag. 2002, 517, 47–51. [Google Scholar] [CrossRef]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef]
- Rieger, R.; Michaelis, A.; Schubert, I.; Doebel, P.; Jank, H.W. Non-random intrachromosomal distribution of chromatid aberrations induced by X-rays, alkylating agents and ethanol in Vicia faba. Mutat. Res. 1975, 27, 69–79. [Google Scholar] [CrossRef]
- Jovtchev, G.; Gateva, S.; Stergios, M.; Kulekova, S. Cytotoxic and genotoxic effects of paraquat in Hordeum vulgare and human lymphocytes in vitro. Environ. Toxicol. 2010, 25, 294–303. [Google Scholar] [CrossRef]
- Živković, L.; Čabarkapa, A.; Marčetić, M.; Kovačević, N.; Bajić, V.; Jovičić, S.; Spremo-Potparević, B. Evaluation of genotoxic and antigenotoxic properties of essential oils of Seseli rigidum Waldst. & Kit. (Apiaceae). Arch. Biol. Sci. Belgrade 2016, 68, 135–144. [Google Scholar]
- Yonis, N.S.; Abduldaium, M.S.; Mohamed, M.E. Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways. Antioxidants 2020, 9, 977. [Google Scholar] [CrossRef]
- Farhath, M.S.S.; Vijaya, P.P.; Vimal, M. Antioxidant activity of Geraniol, Geranial acetate, Gingerol and Eugenol. Res Pharm. 2013, 3, 1–6. [Google Scholar]
- Gateva, S.; Jovtchev, G.; Stankov, A.; Georgieva, A.; Dobreva, A.; Mileva, M. The potential of geraniol to reduce cytotoxic and genotoxic effects of MNNG in plant and human lymphocyte test-systems. S. Afr. J. Bot. 2019, 123, 170–179. [Google Scholar] [CrossRef]
- Ammar, B.R. Potential Effects of Geraniol on Cancer and Inflammation-Related Diseases: A Review of the Recent Research Findings. Molecules 2023, 28, 3669. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Azimullah, S.; Parekh, K.A.; Ojha, S.K.; Beiram, R. Effect of citronellol on oxidative stress, neuroinflammation and autophagy pathways in an in vivo model of Parkinson’s disease. Heliyon 2022, 8, e11434. [Google Scholar] [CrossRef]
- Gunaseelan, S.; Balupillai, A.; Govindasamy, K.; Ramasamy, K.; Muthusamy, G.; Shanmugam, M.; Thangaiyan, R.; Robert, B.M.; Nagarajan, R.P.; Ponniresan, V.K.; et al. Linalool prevents oxidative stress activated protein kinases in single UVB-exposed human skin cells. PLoS ONE 2017, 12, e0176699. [Google Scholar] [CrossRef]
- Tatke, P.; Patil, P.; Gabhe, S. In vitro antioxidant and free radical scavenging activity of extracts of Rosa damascena flower petals. Am. J. Phytomed. Clin. Ther. 2015, 3, 589–601. [Google Scholar]
- Serdal, O.; Kilinik, D.; Yaman, H.; Koc, E. Effects of rose water containing essential rose oil against toxicological effects of chlorpyrifos (Dursban 4) ethyl in Rats. Fresen Environ. Bull 2015, 24, 1009–1015. [Google Scholar]
- Gichner, T.; Velemínský, J. Genetic effects of N-methyl-N′-nitro-N-nitrosoguanidine and its homologs. Mutat. Res.-Rev. Genet. Toxicol. 1982, 99, 129–242. [Google Scholar] [CrossRef]
- Künzel, G.; Gecheff, K.I.; Schubert, I. Different chromosomal distribution patterns of radiation-induced interchange breakpoints in barley: First post-treatment mitosis versus viable offspring. Genome 2001, 44, 128–132. [Google Scholar] [CrossRef]
- Drabløs, F.; Feyzi, E.; Aas, P.A.; Vaagbø, C.B.; Kavli, B.; Bratlie, M.S.; Peña-Diaz, J.; Otterlei, M.; Slupphaug, G.; Krokan, H.E. Alkylation damage in DNA and RNA—Repair mechanisms and medical significance. DNA Repair. 2004, 3, 1389–1407. [Google Scholar] [CrossRef]
- Gateva, S.; Jovtchev, G.; Chanev, C.; Georgieva, A.; Stankov, A.; Dobreva, A.; Mileva, M. Assessment of anti-cytotoxic, anti-genotoxic and antioxidant potential of Bulgarian Rosa alba L. essential oil. Caryologia Int. J. Cytol. Cytosyst. Cytogen. 2020, 73, 371–788. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gateva, S.; Jovtchev, G.; Angelova, T.; Gerasimova, T.; Dobreva, A.; Mileva, M. Cytogenetic Studies on Genoprotective Effect of Rosa damascena Mill. Hydrosol in Plant and Lymphocyte Test Systems. Life 2023, 13, 1753. https://doi.org/10.3390/life13081753
Gateva S, Jovtchev G, Angelova T, Gerasimova T, Dobreva A, Mileva M. Cytogenetic Studies on Genoprotective Effect of Rosa damascena Mill. Hydrosol in Plant and Lymphocyte Test Systems. Life. 2023; 13(8):1753. https://doi.org/10.3390/life13081753
Chicago/Turabian StyleGateva, Svetla, Gabriele Jovtchev, Tsveta Angelova, Tsvetelina Gerasimova, Ana Dobreva, and Milka Mileva. 2023. "Cytogenetic Studies on Genoprotective Effect of Rosa damascena Mill. Hydrosol in Plant and Lymphocyte Test Systems" Life 13, no. 8: 1753. https://doi.org/10.3390/life13081753
APA StyleGateva, S., Jovtchev, G., Angelova, T., Gerasimova, T., Dobreva, A., & Mileva, M. (2023). Cytogenetic Studies on Genoprotective Effect of Rosa damascena Mill. Hydrosol in Plant and Lymphocyte Test Systems. Life, 13(8), 1753. https://doi.org/10.3390/life13081753