
Acetylcholinesterase Inhibition Activity of Hippeastrum papilio (Ravenna) Van Scheepen (Amaryllidaceae) Using Zebrafish Brain Homogenates

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction
2.3. GC-MS Analysis
2.4. Alkaloid Identification
2.5. Alkaloid Quantification
2.6. Determination of AChE Activity
2.7. Computational Experiments
2.7.1. Comparative Modeling
2.7.2. Molecular Docking Protocol
2.8. Ethical and Legal Considerations
3. Results and Discussion
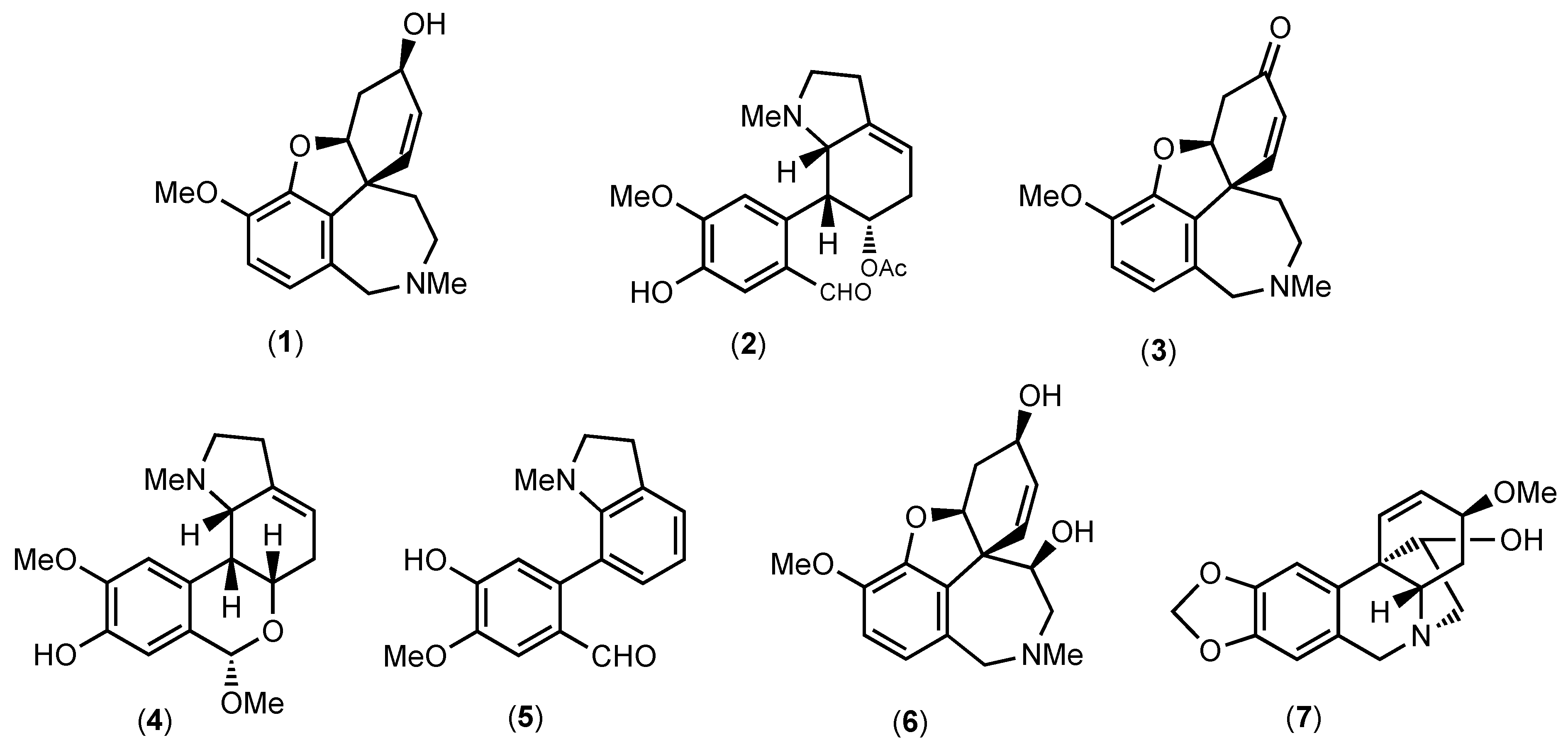
3.1. Chemical Profile
3.2. AChE Inhibitory Activity
3.3. Computational Results
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardy, K. Paleomedicine and the Evolutionary Context of Medicinal Plant Use. Rev. Bras. Farmacogn. 2021, 31, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Cordell, G.A.; Quinn-Beattie, M.L.; Farnsworth, N.R. The potential of alkaloids in drug discovery. Phytother. Res. 2001, 15, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Feher, M.; Schmidt, J.M. Property Distributions: Differences between Drugs, Natural Products, and Molecules from Combinatorial Chemistry. J. Chem. Inf. Model. 2002, 43, 218–227. [Google Scholar] [CrossRef]
- Lu, J.-J.; Bao, J.-L.; Chen, X.-P.; Huang, M.; Wang, Y.-T. Alkaloids Isolated from Natural Herbs as the Anticancers Agents. Evid. Based Complement. Altern. Med. 2012, 2012, 485042. [Google Scholar] [CrossRef]
- Tewari, D.; Stankiewicz, A.M.; Mocan, A.; Sah, N.A.; Tzvetkov, N.T.; Huminiecki, L.; Horbanczuk, J.O.; Atanasov, A.G. Ethnopharmacological Approaches for Dementia Therapy and Significance of Natural Products and Herbal Drugs. Front. Aging Neurosci. 2018, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Masondo, N.A.; Stafford, G.I.; Aremu, A.O.; Makunga, N.P. Acetylcholinesterase inhibitors from southern African plants: An overview of ethnobotanical, pharmacological potential and phytochemical research including and beyond Alzheimer’s disease treatment. S. Afr. J. Bot. 2019, 120, 39–64. [Google Scholar] [CrossRef]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus alkaloids. In The Alkaloids; Cordell, G.A., Ed.; Elsevier Scientific Publishing: Amsterdam, The Netherlands, 2006; pp. 87–179. ISSN 1099-4831. [Google Scholar]
- The Angiosperm Phylogeny (APG). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of AA. In The Alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 83, pp. 113–185. ISSN 1099-4831. [Google Scholar]
- Konrath, E.L.; Passos, C.S.; Klein-Junior, L.C.; Henriques, A.T. Alkaloids as a source of potential anticholinesterase inhibitors for the treatment of Alzheimer’s disease. J. Pharm. Pharmacol. 2013, 65, 1701–1725. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.; Gill, K.D.; Mahdi, A.A. Therapeutics of Alzheimer’s disease: Past, present and future. Neuropharmacology 2014, 76, 27–50. [Google Scholar] [CrossRef]
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Amaryllidaceae alkaloids: Absolute configuration and biological activity. Chirality 2017, 29, 486–499. [Google Scholar] [CrossRef]
- Panula, P.; Sallinen, V.; Sundvik, M.; Kolehmainen, J.; Torkko, V.; Tiittula, A.; Moshnyakov, M.; Podlasz, P. Modulatory Neurotransmitter Systems and Behavior: Towards Zebrafish Models of Neurodegenerative Diseases. Zebrafish 2006, 3, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R.; Fernandes, Y.; Pereira, T. Zebrafish (Danio rerio) responds to the animated image of a predator: Towards the development of an automated aversive task. Behav. Brain Res. 2009, 201, 318–324. [Google Scholar] [CrossRef]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueff, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef]
- Marinho, C.S.; Matias, M.V.F.; Brandao, I.G.F.; Santos, E.L.; Machado, S.S.; Zanta, C.L.P.S. Characterization and kinetic study of the brain and muscle acetylcholinesterase from Danio rerio. Comp. Biochem. Phys. C 2019, 222, 11–18. [Google Scholar] [CrossRef]
- Nasevicius, A.; Ekker, S.C. Effective targeted gene ‘knockdown’ in zebrafish. Nat. Genet. 2000, 26, 216–220. [Google Scholar] [CrossRef]
- Gerlai, R. High-Throughput Behavioral Screens: The First Step towards Finding Genes Involved in Vertebrate Brain Function Using Zebrafish. Molecules 2010, 15, 2609–2622. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Santana, S.; Rico, E.P.; Burgos, J.S. Can zebrafish be usede as animal model to study Alzheimr’s disease? Am. J. Neurodegener. Dis. 2012, 1, 32–48. [Google Scholar]
- Berkov, S.; Bastida, J.; Nikolova, M.; Viladomat, F.; Codina, C. Rapid TLC/GC-MS identification of acetylcholinesterase inhibitors in alkaloid extracts. Phytochem. Anal. 2008, 19, 411–419. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Šali, A.; Blundell, T.L. Comparative Protein Modelling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Shen, M.-Y.; Sali, A. Statistical potential for assessment and prediction of protein structures. Protein Sci. 2006, 15, 2507–2524. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with pyrx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC–MS combined with k-means cluster analysis. Ind. Crops Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- De Andrade, J.P.; Berkov, S.; Viladomat, F.; Codina, C.; Zuanazzi, J.A.S.; Bastida, J. Alkaloids from Hippeastrum papilio. Molecules 2011, 16, 7097–7104. [Google Scholar] [CrossRef]
- Guo, Y.; de Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Tallini, L.R.; Borges, W.S.; Viladomat, F.; Nair, J.J.; Zuanazzi, J.A.S.; Bastida, J. New Alkaloids from Hippeastrum papilio (Ravenna) VanScheepen. Helv. Chim. Acta 2016, 99, 143–147. [Google Scholar] [CrossRef]
- Berkov, S.; Bastida, J.; Codina, C.; de Andrade, J.P.; Berbee, R.L.M. Extract of Hippeastrum papilio Rich in Galanthamine. EP2999480B1, 4 September 2019. [Google Scholar]
- Ríos, S.; Berkov, S.; Martínez-Francés, V.; Bastida, J. Biogeographical Patterns and Phenological Changes in Lapiedra martinezii Lag. Related to Its Alkaloid Diversity. Chem. Biodivers. 2013, 10, 1220–1238. [Google Scholar] [CrossRef]
- Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schmeda-Hirschmann, G. Cholinesterase Inhibition Activity, Alkaloid Profiling and Molecular Docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. [Google Scholar] [CrossRef]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Atanassov, A.; Viladomat, F.; Bastida, J.; Codina, C. Plant Sources of Galanthamine: Phytochemical and Biotechnological Aspects. Biotechnol. Biotechnol. Equip. 2009, 23, 1170–1176. [Google Scholar] [CrossRef]
- Salehi, S.M.H.; Azadi, B.; Amin, G.; Amini, M.; Sharifzadeh, M. The first phytochemical report of Galanthus transcaucasicus Fomin. DARU 2010, 18, 124–127. [Google Scholar]
- Torras-Claveria, L.; Berkov, S.; Jáuregui, O.; Caujapé, J.; Viladomat, F.; Codina, C.; Bastida, J. Metabolic profiling of bioactive Pancratium canariense extracts by GC-MS. Phytochem. Anal. 2010, 21, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Cahlíková, L.; Macáková, K.; Zavadil, S.; Jiros, P.; Opletal, L.; Urbanová, K.; Jahoodár, L. Analysis of Amaryllidaceae Alkaloids from Chlidanthus fragrans by GC-MS and their Cholinesterase Activity. Nat. Prod. Commun. 2011, 6, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Cahlíková, L.; Locárek, M.; Benesová, N.; Kucera, R.; Chlebek, J.; Novák, Z.; Opletal, L. Isolation and Cholinesterase Inhibitory Activity of Narcissus Extracts and Amaryllidaceae alkaloid. Nat. Prod. Commun. 2013, 8, 781–785. [Google Scholar] [CrossRef]
- De Andrade, J.P.; Giordani, R.B.; Torras-Claveria, L.; Pigni, N.B.; Berkov, S.; Font-Bardia, M.; Calvet, T.; Konrath, E.; Bueno, K.; Sachett, L.G.; et al. The Brazilian Amaryllidaceae as a source of acetylcholinesterase inhibitory alkaloids. Phytochem. Rev. 2016, 15, 147–160. [Google Scholar] [CrossRef]
- Sebben, C.; Giordani, R.B.; de Andrade, J.P.; Berkov, S.; Osorio, J.E.; Sobral, M.; Almeida, M.V.; Henriques, A.T.; Bastida, J.; Zuanazzi, J.A.S. New lycosinine derivative from Hippeastrum breviflorum. Rev. Bras. Farmacogn. 2015, 25, 353–355. [Google Scholar] [CrossRef]
- Greig, N.H.; Lahiri, D.K.; Sambamurti, K. Butyrylcholinesterase: An Important New Target in Alzheimer’s Disease Therapy. Int. Psychogeriatr. 2002, 14, 77–91. [Google Scholar] [CrossRef]
- Bertrand, C.; Chatonnet, A.; Takke, C.; Yan, Y.; Postlethwait, J.; Toutant, J.-P.; Cousin, X. Zebrafish Acetylcholinesterase Is Encoded by a Single Gene Localized on Linkage Group 7. J. Biol. Chem. 2001, 276, 464–474. [Google Scholar] [CrossRef]
- Pitchai, A.; Nagasundaram, N.; Prakash, V.S.G.; Kannan, R.R. Zebrafish bio-assay guided isolation of human acetylcholinesterase inhibitory trans-tephrostachin from Tephrosia purpurea (L.) Pers. Neurosci. Lett. 2018, 687, 268–275. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Garro, A.; Pigni, N.B.; Agüero, M.B.; Roitman, G.; Slanis, A.; Enriz, R.D.; Feresin, G.E.; Bastida, J.; Tapia, A. Cholinesterase-inhibitory effect and in silico analysis of alkaloids from bulbs of Hieronymiella species. Phytomedicine 2018, 39, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Bores, G.M.; Huger, F.P.; Petko, W.; Mutlib, A.E.; Camacho, F.; Rush, D.K.; Selk, D.E.; Wolf, V.; Kosley, R.W.; Davis, L.; et al. Pharmacological evaluation of novel Alzheimer’s disease therapeutics: Acetylcholinesterase inhibitors related to galanthamine. J. Pharmacol. Exp. Ther. 1996, 277, 728–738. [Google Scholar]
- Zarotsky, V.; Sramek, J.J.; Cutler, N.R. Galanthamine hydrobromide: An agent for Alzheimer’s disease. Am. J. Health-Syst. Pharm. 2003, 60, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Samochocki, M.; Höffle, A.; Fehrenbacher, A.; Jostock, R.; Ludwig, J.; Christner, C.; Radina, M.; Zerlin, M.; Ullmer, C.; Pereira, E.F.R.; et al. Galantamine Is an Allosterically Potentiating Ligand of Neuronal Nicotinic but Not of Muscarinic Acetylcholine Receptors. Pharmacol. Exp. Ther. 2003, 305, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Vera, J.d.C.; Buitrago-Díaz, A.A.; Possamai, L.M.; Timmers, L.F.S.M.; Tallini, L.R.; Bastida, J. Alkaloid profile and cholinesterase inhibition activity of five species of Amaryllidaceae family collected from Mérida state-Venezuela. S. Afr. J. Bot. 2021, 136, 126–136. [Google Scholar] [CrossRef]
- Rosenberry, T.L.; Brazzolotto, X.; Macdonald, I.R.; Wandhammer, M.; Trovaslet-Leroy, M.; Darvesh, S.; Nachon, F. Comparison of the Binding of Reversible Inhibitors to Human Butyrylcholinesterase and Acetylcholinesterase: A Crystallographic, Kinetic and Calorimetric Study. Molecules 2017, 22, 2098. [Google Scholar] [CrossRef] [PubMed]
- Stierand, K.; Maaß, P.; Rarey, M. Molecular complexes at a glance: Automated generation of two-dimensional complex diagrams. Bioinformatics 2006, 22, 1710–1716. [Google Scholar] [CrossRef] [PubMed]
- Fricker, P.; Gastreich, M.; Rarey, M. Automated Generation of Structural Molecular Formulas under Constraints. J. Chem. Inf. Comput. Sci. 2004, 44, 1065–1078. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkaloid | Rt | Fragmentation Pattern, m/z (Relative Intensity, %) | Amount a | % b |
|---|---|---|---|---|
| galanthamine (1) | 27.247 | 287 (78), 286 (100), 270 (12), 244 (23), 216 (32), 174 (26) | 1286.03 | 67.3 |
| papiline (2) | 27.715 | 286 (3), 177 (16), 110 (8), 109 (100), 108 (24), 94 (5) | 89.77 | 4.7 |
| narwedine (3) | 28.395 | 285 (80), 284 (100), 242 (19), 216 (21), 199 (22), 174 (31) | 61.31 | 3.2 |
| hippapiline (4) | 28.564 | 177 (15), 110 (9), 109 (100), 108 (23), 94 (7), 81 (5) | 38.25 | 2.0 |
| 9-O-demethyllycosinine B (5) | 29.881 | 283 (100), 255 (75), 254 (83), 240 (31), 222 (39), 194 (20) | 56.95 | 2.9 |
| 11β-hydroxygalanthamine (6) | 30.009 | 303 (24), 286 (10), 231 (22), 230 (100), 213 (34), 197 (10) | 106.57 | 5.7 |
| haemanthamine (7) | 30.657 | 301 (13), 273 (18), 272 (100), 240 (19), 181 (26), 153 (14) | 271.78 | 14.2 |
| Total | - | - | 1910.66 | 100.0 |
| Alkaloid | Estimated Free Binding Energy |
|---|---|
| galanthamine (1) | −9.3 |
| papiline (2) | −9.1 |
| narwedine (3) | −9.5 |
| hippapiline (4) | −9.6 |
| 9-O-demethyllycosinine B (5) | −9.0 |
| 11β-hydroxygalanthamine (6) | −9.7 |
| haemanthamine (7) | −8.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tallini, L.R.; da Silva, C.R.; Jung, T.; Alves, E.d.O.; Baldin, S.L.; Apel, M.; Timmers, L.F.S.M.; Rico, E.P.; Bastida, J.; Zuanazzi, J.A.S. Acetylcholinesterase Inhibition Activity of Hippeastrum papilio (Ravenna) Van Scheepen (Amaryllidaceae) Using Zebrafish Brain Homogenates. Life 2023, 13, 1721. https://doi.org/10.3390/life13081721
Tallini LR, da Silva CR, Jung T, Alves EdO, Baldin SL, Apel M, Timmers LFSM, Rico EP, Bastida J, Zuanazzi JAS. Acetylcholinesterase Inhibition Activity of Hippeastrum papilio (Ravenna) Van Scheepen (Amaryllidaceae) Using Zebrafish Brain Homogenates. Life. 2023; 13(8):1721. https://doi.org/10.3390/life13081721
Chicago/Turabian StyleTallini, Luciana R., Camila Rockenbach da Silva, Tatiana Jung, Elen de Oliveira Alves, Samira Leila Baldin, Miriam Apel, Luis F. S. M. Timmers, Eduardo Pacheco Rico, Jaume Bastida, and José Angelo S. Zuanazzi. 2023. "Acetylcholinesterase Inhibition Activity of Hippeastrum papilio (Ravenna) Van Scheepen (Amaryllidaceae) Using Zebrafish Brain Homogenates" Life 13, no. 8: 1721. https://doi.org/10.3390/life13081721
APA StyleTallini, L. R., da Silva, C. R., Jung, T., Alves, E. d. O., Baldin, S. L., Apel, M., Timmers, L. F. S. M., Rico, E. P., Bastida, J., & Zuanazzi, J. A. S. (2023). Acetylcholinesterase Inhibition Activity of Hippeastrum papilio (Ravenna) Van Scheepen (Amaryllidaceae) Using Zebrafish Brain Homogenates. Life, 13(8), 1721. https://doi.org/10.3390/life13081721