


Critical Assessment of the Anti-Inflammatory Potential of Usnic Acid and Its Derivatives—A Review

 , , and
, , and
Abstract
1. Introduction
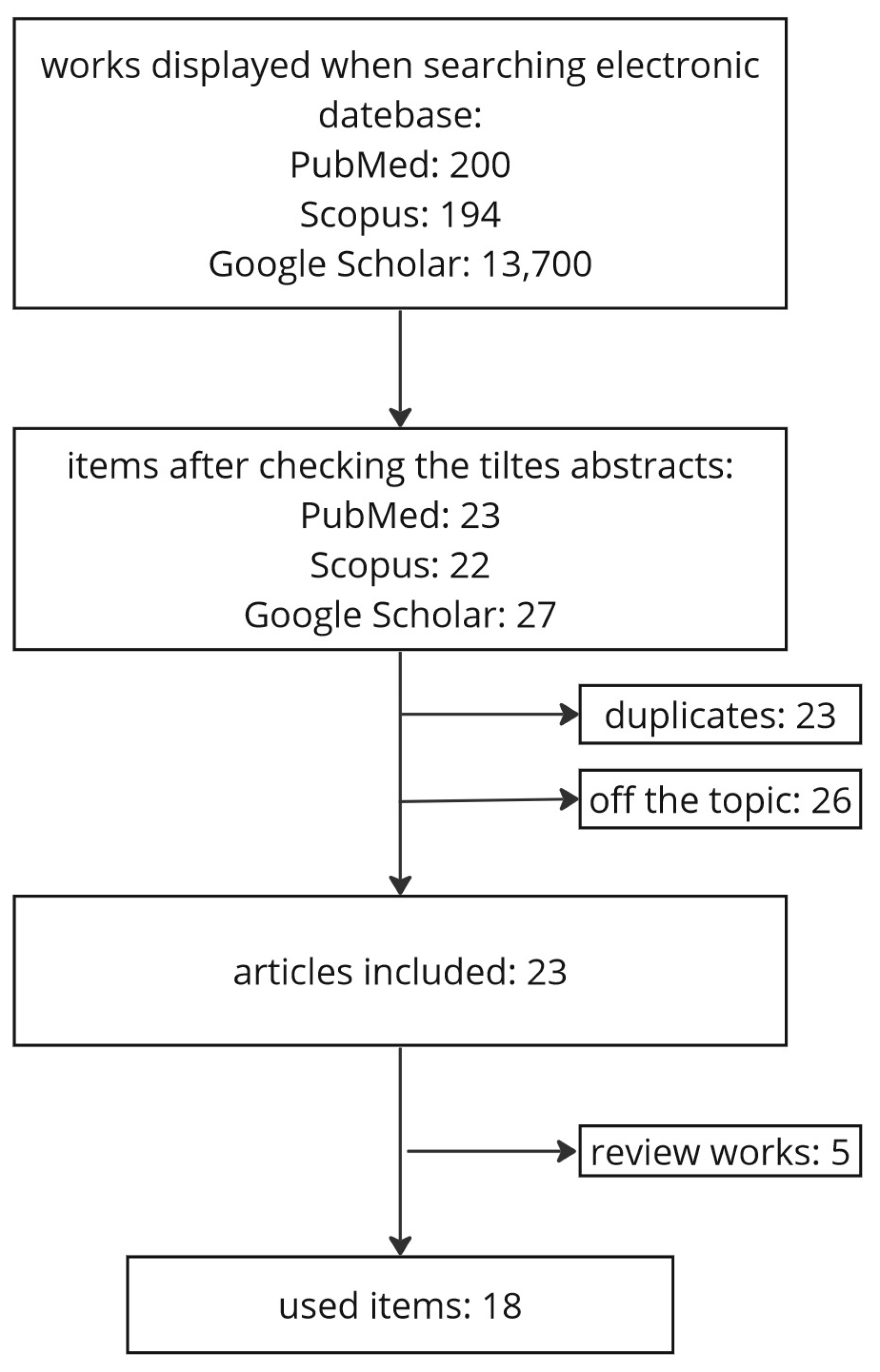
2. Materials and Methods
3. Anti-Inflammatory Potential of Usnic Acid
3.1. Results from the In Vitro Studies
3.2. Results from In Vivo Studies
4. Anti-Inflammatory Potential of Synthetic Usnic Acid Derivatives In Vitro and In Vivo
5. Limitations of the Studies Included in the Review
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchi, S.; Guilbaud, E.; Tait, S.W.; Yamazaki, T.; Galluzzi, L. Mitochondrial control of inflammation. Nat. Rev. Immunol. 2023, 23, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.D.R.; Barreto Arantes, M.; Menezes de Faria Pereira, S.; Leandro da Cruz, L.; de Souza Passos, M.; Pereira de Moraes, L.; Vieira, I.J.C.; Barros de Oliveira, D. Plants as Sources of Anti-Inflammatory Agents. Molecules 2020, 25, 3726. [Google Scholar] [CrossRef] [PubMed]
- de Cássia da Silveira e Sá, R.; Andrade, L.N.; de Sousa, D.P. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef] [PubMed]
- Karpel, E. Mediatory ogólnoustrojowej odpowiedzi zapalnej—Znaczenie w praktyce klinicznej intensywnej terapii. Anestezjol. Intensywna Ter. 2001, 3, 181–190. [Google Scholar]
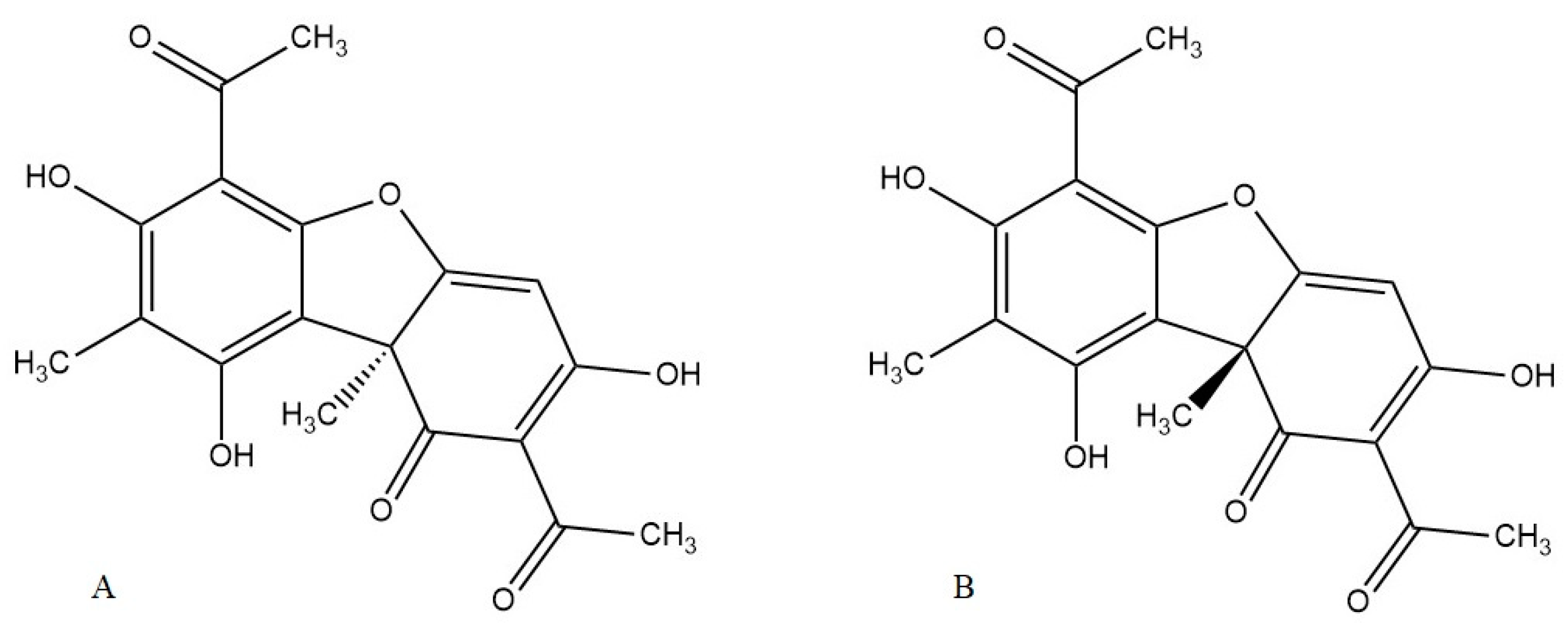
- Galanty, A.; Paśko, P.; Podolak, I. Enantioselective activity of usnic acid: A comprehensive review and future perspectives. Phytochem. Rev. 2019, 18, 527–548. [Google Scholar] [CrossRef]
- Kumar, S.; Muller, K. Lichen Metabolites. 2. Antiproliferative and cytotoxic activity of gyrophoric, usnic, and diffractaic acid on human keratinocyte growth. J. Nat. Prod. 1999, 62, 821–823. [Google Scholar] [CrossRef]
- Bucar, F.; Schneider, I.; Ogmundsdóttir, H.; Ingólfsdóttir, K. Anti-proliferative lichen compounds with inhibitory activity on 12(S)-HETE production in human platelets. J. Phytomed. 2004, 11, 602–606. [Google Scholar] [CrossRef]
- Jin, J.; Li, C.; He, L. Down-regulatory effect of usnic acid on nuclear factor-κB-dependent tumor necrosis factor-α and inducible nitric oxide synthase expression in lipopolysaccharide-stimulated macrophages RAW 264.7. Phytother. Res. 2008, 22, 1605–1609. [Google Scholar] [CrossRef]
- Huang, Z.; Tao, J.; Ruan, J.; Li, C.; Zheng, G. Anti-inflammatory effects and mechanisms of usnic acid, a compound firstly isolated from lichen Parmelia saxatilis. J. Med. Plant Res. 2014, 8, 197–207. [Google Scholar]
- Galanty, A.; Zagrodzki, P.; Gdula-Argasińska, J.; Grabowska, K.; Koczurkiewicz-Adamczyk, P.; Wróbel-Biedrawa, D.; Podolak, I.; Pękala, E.; Paśko, P. A comparative survey of anti-melanoma and anti-inflammatory potential of usnic acid enantiomers—A comprehensive in vitro approach. Pharmaceuticals 2021, 14, 945. [Google Scholar] [CrossRef]
- Yildirim, M.; Degirmenci, U.; Akkapulu, M.; Gungor, M.; Oztornacı, R.O.; Berkoz, M.; Comelekoglu, U.; Yalın, A.E.; Yalın, S. Anti-inflammatory effects of usnic acid in breast cancer. Russ. J. Bioorg. Chem. 2022, 14, 945. [Google Scholar] [CrossRef]
- Vijayakumar, C.S.; Viswanathan, S.; Reddy, M.K.; Parvathavarthini, S.; Kundu, A.B.; Sukumar, E. Anti-inflammatory activity of (+)-usnic acid. Fitoterapia 2000, 71, 564–568. [Google Scholar] [CrossRef]
- Nunes, P.S.; Albuquerque-Júnior, R.L.C.; Cavalcante, D.R.R.; Dantas, M.D.M.; Cardoso, J.C.; Bezerra, M.S.; Souza, J.C.C.; Russo Serafini, M.; Quitans, L.J., Jr.; Bonjardim, L.R.; et al. Collagen-based films containing liposome-loaded usnic acid as dressing for dermal burn healing. Biomed. Res. Int. 2011, 2011, 761593. [Google Scholar] [CrossRef]
- Nunes, P.S.; Rabelo, A.S.; Campos de Souza, J.C.; Vasconcelos Santana, B.; Monteiro Menezes da Silva, T.; Russo Serafini, M.; Dos Passos Menezes, P.; Dos Santos Lima, B.; Cordeiro Cardoso, J.; Santana Alves, J.C.; et al. Gelatin-based membrane containing usnic acid-loaded liposome improves dermal burn healing in a porcine model. Int. J. Pharm. 2016, 513, 473–482. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, Y.; Li, Y.; Bai, H.; Ma, T.; Song, X.; Zhao, J.; Gao, L. The effects of sodium usnic acid by topical application on skin wound healing in rats. Biomed. Pharmacother. 2018, 97, 587–593. [Google Scholar] [CrossRef]
- Erfani, S.; Valadbeigi, T.; Aboutaleb, N.; Karimi, N.; Moghimi, A.; Khaksari, M. Usnic acid improves memory impairment after cerebral ischemia/reperfusion injuries by anti-neuroinflammatory, anti-oxidant, and anti-apoptotic properties. Iran. J. Basic Med. Sci. 2020, 23, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, Y.; Ha, S.; Chung, H.Y.; Kim, H.; Hur, J.S.; Lee, J. Anti-inflammatory effects of usnic acid in an MPTP-induced mouse model of Parkinson’s disease. Brain Res. J. 2020, 1730, 146642. [Google Scholar] [CrossRef]
- Cazarin, C.A.; Dalmagro, A.P.; Gonçalves, A.E.; Boeing, T.; Mota da Silva, L.; Correa, R.; Klein-Júnior, L.C.; Carlesso Pinto, B.; Savoldi Lorenzett, T.; Uchoa da Costa Sobrinho, T.; et al. Usnic acid enantiomers restore cognitive deficits and neurochemical alterations induced by Aβ1-42 in mice. Behav. Brain Res. 2021, 397, 112945. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.J.; Peng, W.; Zhao, J.H.; Yang, H.J.; Qu, L.L.; Wang, C.; Kong, L.Y.; Wang, X.B. Usnic acid derivatives as tau-aggregation and neuroinflammation inhibitors. Eur. J. Med. Chem. 2020, 187, 111961. [Google Scholar] [CrossRef]
- Su, Z.-Q.; Mo, Z.Z.; Liao, J.-B.; Feng, X.-X.; Liang, Y.-Z.; Zhang, X.; Liu, Y.-H.; Chen, X.-Y.; Chen, Z.-W.; Su, Z.-R.; et al. Usnic acid protects LPS-induced acute lung injury in mice through attenuating inflammatory responses and oxidative stress. Int. Immunopharmacol. 2014, 22, 371–378. [Google Scholar] [CrossRef]
- Huang, X.Q.; Ai, G.X.; Zheng, X.H.; Liao, H.J. Usnic acid ameliorates bleomycin-induced pulmonary fibrosis in mice via inhibition of inflammatory responses and oxidative stress. Trop. J. Pharm. Res. 2019, 18, 2563–2569. [Google Scholar]
- Wang, H.; Xuan, M.; Huang, C.; Wang, C. Advances in research on bioactivity, toxicity, metabolism, and pharmacokinetics of usnic acid in vitro and in vivo. Molecules 2022, 27, 7469. [Google Scholar] [CrossRef] [PubMed]
- da Silva Santos, N.P.; Nascimento, S.C.; Wanderley, M.S.O.; Pontes-Filho, N.T.; da Silva, J.F.; de Castro, C.M.M.B.; Santos-Magalhaes, N.S. Nanoencapsulation of usnic acid: An attempt to improve antitumour activity and reduce hepatotoxicity. Eur. J. Pharm. Biopharm. 2006, 64, 154–160. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Kaproń, B.; Plech, T.; Żarowski, M.; Cielecka-Piontek, J. Lichen-derived compounds and extracts as biologically active substances with anticancer and neuroprotective properties. Pharmaceuticals 2021, 14, 1293. [Google Scholar] [CrossRef]
- Gaweł, M.; Potulska-Chromik, A. Neurodegenerative diseases: Alzheimer’s and Parkinson’s disease. Postępy Nauk Med. 2015, 28, 468–472. [Google Scholar]
- Parekh, D.; Dancer, R.C.; Thickett, D.R. Acute lung injury. Clin. Med. 2011, 11, 615–618. [Google Scholar] [CrossRef]
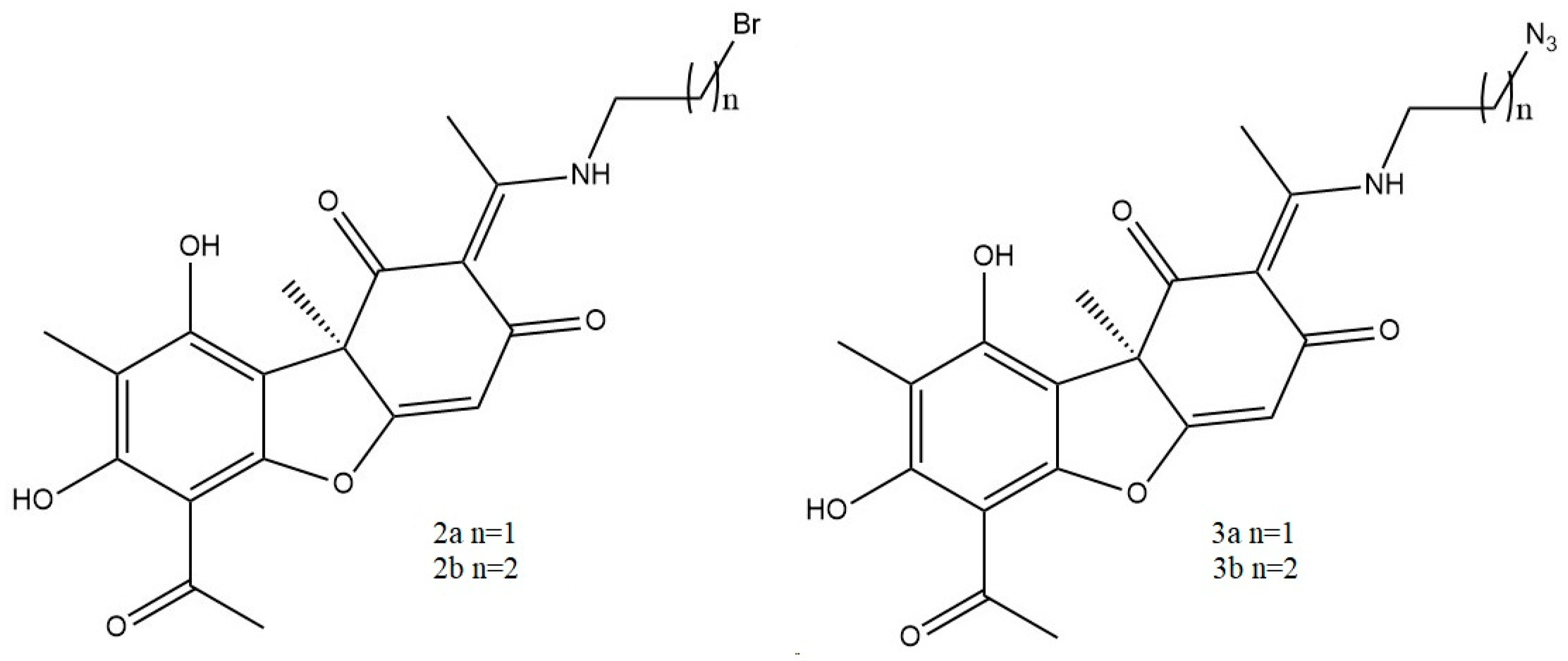
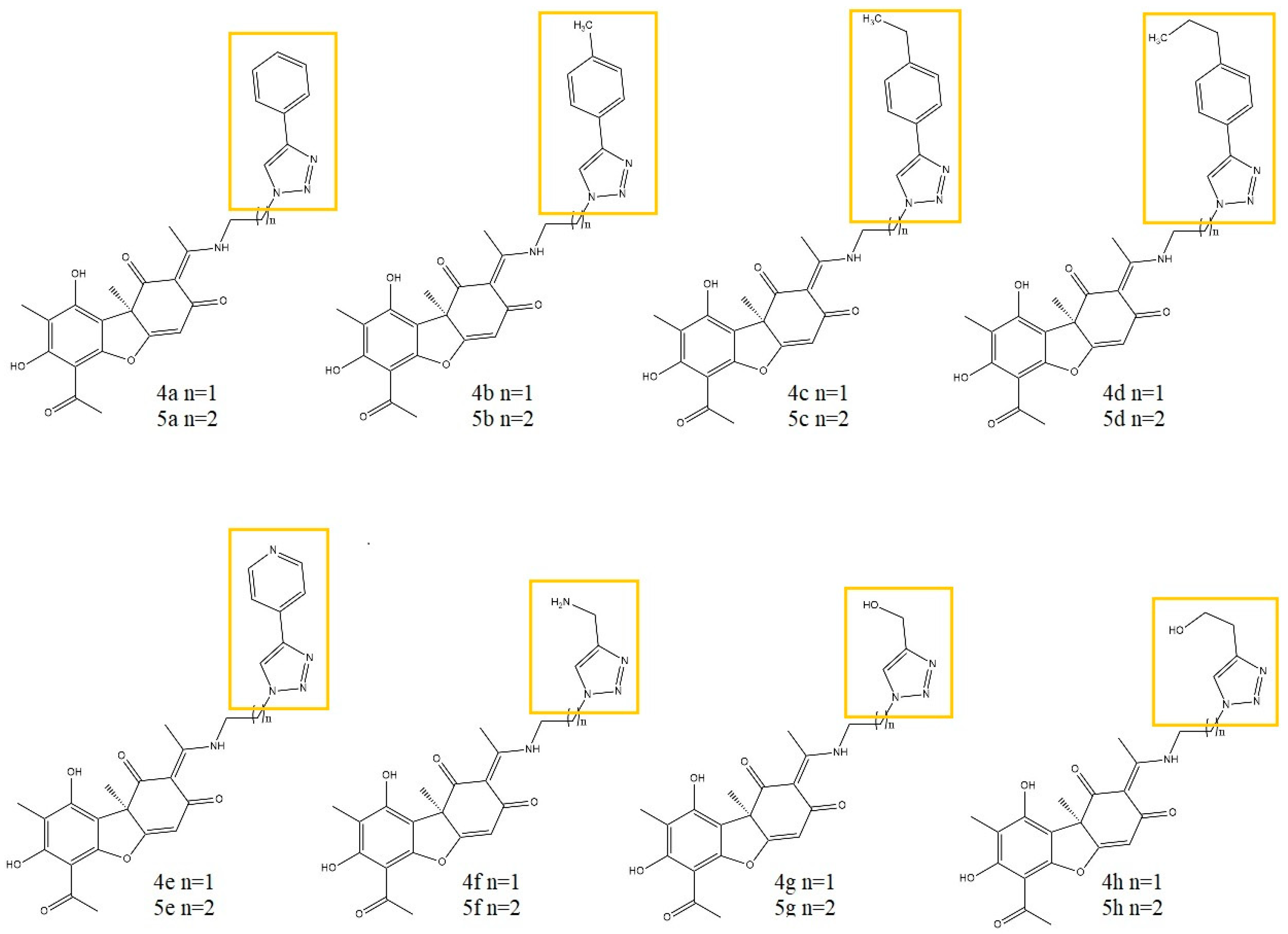
- Vanga, N.R.; Kota, A.; Sistla, R. Synthesis and anti-inflammatory activity of novel triazole hybrids of (+)-usnic acid, the major dibenzofuran metabolite of the lichen Usnea longissima. Mol. Divers. 2017, 21, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Somasekhar, T.; Javadi, M.; Sistla, R. Synthesis of novel anti-inflammatory usnic acid-based imidazolium salts. Eur. Chem. Bull. 2021, 10, 67–72. [Google Scholar] [CrossRef]
- Guo, L.; Shi, Q.; Fang, J.L.; Mei, N.; Ali, A.A.; Lewis, S.M.; Frankos, V.H. Review of usnic acid and Usnea barbata toxicity. J. Environ. Health Part C 2008, 26, 317–338. [Google Scholar] [CrossRef]
- Croce, N.; Pitaro, M.; Gallo, V.; Antonini, G. Toxicity of Usnic Acid: A Narrative Review. J. Toxicol. 2022, 2022, 8244340. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Vitro Model | Experimental Conditions | Effects | Ref. |
|---|---|---|---|
| bovine polymorphonuclear leukocytes (PMNL) | (+)-UA Reference: nordihydroguaiaretic acid, anthralin Groups: Ca-ionophore A23187-stimulated cells Methods: RP-HPLC (inhibition of LTB4 biosynthesis) |
| [6] |
| human platelets | (+)-UA (3.33–100 μg/mL) Reference: baicalein (IC50 = 24.6 μM) Methods: optical density, RP-HPLC (inhibition of platelet-type 12(S)-LOX) |
| [7] |
| RAW 264.7 macrophages | UA (0.5–400 μM) Reference: none Groups: LPS-stimulated cells, untreated cells Methods: Griess reagent (NO), ELISA assay (TNF-α, iNOS, NF-κB, I-κB). |
| [8] |
| RAW 264.7 cells | UA (1, 5, 10 μg/mL) Reference: dexamethasone 0.5 μg/mL Groups: LPS-stimulated cells, untreated cells (control) Methods: ELISA assay (TNF-α, IL-1β, IL-6, IL-10), Griess reagent (NO), RT-PCR (TNF-α mRNA, COX2 mRNA, iNOS mRNA, HO-1 mRNA), immunocytochemical assay (NF-κB), Western Blot (COX-2, HO-1) |
| [9] |
| RAW 264.7 cells | (+)-UA, (−)-UA (10, 25 µg/mL) Reference: dexamethasone 0.5 μg/mL Groups: LPS-stimulated cells, untreated cells (control) Methods: ELISA assay (TNF-α, IL-6) Griess reagent (NO), Western Blot (TLR4, cPLA2, COX-1, COX-2). |
| [10] |
| MCF-7 breast cancer cells | UA (0.623–15,638 µM) Reference: none Groups: untreated (control) Methods: biochemical analysis (MDA, GSH), Griess reagent (NO), ELISA assay (PGE2, IL-2, IL-6, TNF-α), Bio-Plex assay (VEGF), RT-PCR (COX-2, iNOS) |
| [11] |
| In Vivo Model | Experimental Conditions | Effects | Ref. |
|---|---|---|---|
| Induced chronic and acute inflammation in Wistar rats (n = 30) | (+)-UA: 25, 50, 100 mg/kg orally (p.o.) Reference: ibuprofen 100 mg/kg Different groups: untreated control Methods: volume of paw edema, weight of cotton pellets. |
| [12] |
| Wound healing models | |||
| Burn wound in male Wistar rats (n = 45) | Collagen film with liposomal UA: 330 mg/4 cm2, dermal application for 7, 14, and 21 days. Reference: no data Different groups: collagen film, collagen film with empty liposomes. Methods: histological assessment of inflammatory profile, epithelization rates, collagen deposition, mean of myofibroblasts for histological field. |
| [13] |
| Burn wound in a porcine model (n = 9) | Gelatin-based membranes with liposomal UA: 127.02 mg/7 cm2, dermal application for 8, 18, and 30 days. Reference: ointment with silver sulfadiazine. Different groups: duoDerme® dressing. Methods: histological assessment of burn healing grading, collagen deposition. |
| [14] |
| Healing of wound in 8-week-old male Wistar rats (n = 64) | SUA: 38.4 mg/L in DMSO, daily dermal application for 21 days. Reference: gentamicin sulfate 0.01%. Different groups: untreated control, pure DMSO. Methods: wound area measured at 3, 7, 10, and 14 days after wounding. Histological assessment, immunohistochemistry analysis (VEGF). |
| [15] |
| Neurodegenerative diseases models | |||
| Model of cerebral ischemia/reperfusion by 20-min occlusion of the carotid arteries in male Wistar rats (n = 42) | UA: 25 mg/kg in DMSO, intraperitoneally (i.p.), 20 min of ischemia, and 48 h of reperfusion. Reference: no data Different groups: sham-operated, untreated control. Methods: Morris water maze task, spatial training test, spatial probe test, immunohistochemistry analysis (caspase-3, GFAP, Iba-1), biochemical assessment (SOD, GSH, MDA). |
| [16] |
| MPTP-induced Parkinson’s disease model in mice C57BL/6 (n = 40) | UA: 5 and 25 mg/kg intraperitoneally (i.p.) used daily for 10 days before MPTP-induced Parkinson’s disease. Reference: no data Different groups: sham control, untreated control. Methods: motor performance testing (rota-rod), immunocytochemical and immunochemical tests (Iba-1, GFAP, iNOS). |
| [17] |
| Aβ1-42-induced Alzheimer’s disease model in female mice (n = 81) | (R)-(+)- and (S)-(−)-UA: 25, 50, and 100 mg/kg, orally (p.o.) for 24 days. Reference: donepezil 2 mg/kg. Different groups: naïve, untreated control, sham-operated, Methods: open field test, novel object recognition test, Morris water maze task, Inhibitory-avoidance test, biochemical analysis (SOD, GSH, LOOH, MPO, IL-1β). |
| [18] |
| Okadaic acid-induced memory impairment in male rats SD (n = 32) | UA derivative No 30 *: 5 and 10 mg/kg, intraperitoneally (i.p.) for 7 days after okadaic acid injection. Reference: no data Different groups: sham-operated, natrium chloratum 0.9%. Methods: Morris water maze task. |
| [19] |
| Lung diseases models | |||
| LPS-induced acute lung injury (ALI) in mice (n = no data) | UA: 25, 50, or 100 mg/kg used daily for 5 days intratracheal. Reference: dexamethasone 5 mg/kg. Different groups: naïve, untreated control. Methods: histological assessment of BALF, immunochemical analysis (MPO, MDA, TNF-α, IL-6, IL-10, IL-8, MIP-2, GSH, SOD). |
| [20] |
| Bleomycin-induced lung fibrosis in mice (n = no data) | UA: 25, 50, or 100 mg/kg with bleomycin 15 mg/kg used daily for 21 days i.p. Reference: prednisone acetate 5 mg/kg. Different groups: natrium chloratum 0.9%, untreated control. Methods: histological assessments, immunochemical analysis (TGF-β1, TNF-α, IL-1β,IL-6, SOD, MDA). |
| [21] |
| Cellular Model | Experimental Conditions | Effects | Ref. |
|---|---|---|---|
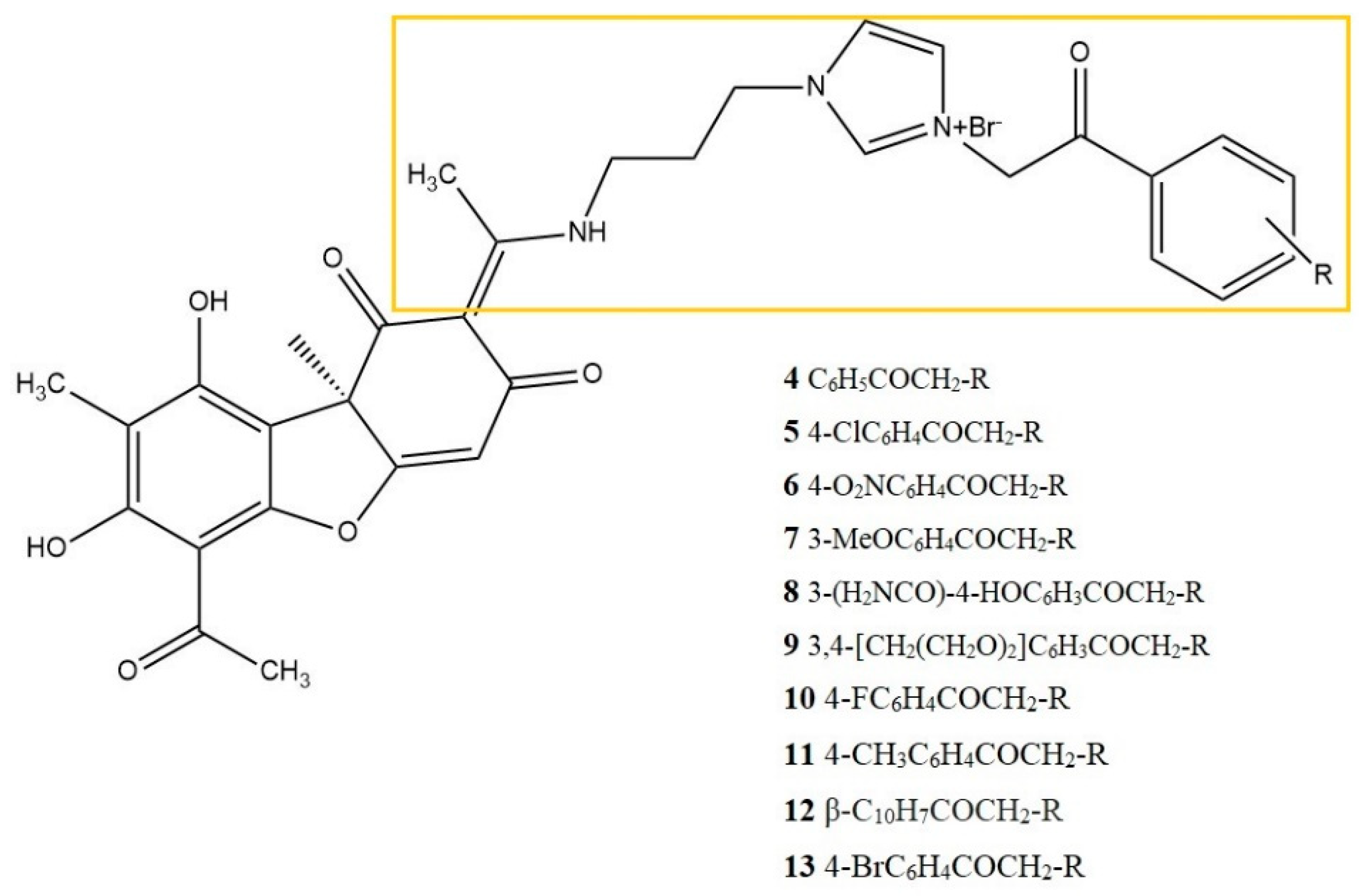
| lymphoma U937 cells | 16 derivatives of UA (10 μM) Reference: prednisolone 10 μM groups: LPS-stimulated cells Methods: ELISA assay (IL-1β, TNF-α) |
| [27] |
| lymphoma U937 cells | UA derivatives No 4-13 (10 μM) Reference: dexamethasone 10 μM. Different groups: LPS-stimulated cells, Methods: ELISA assay (IL-1β, TNF-α) |
| [28] |
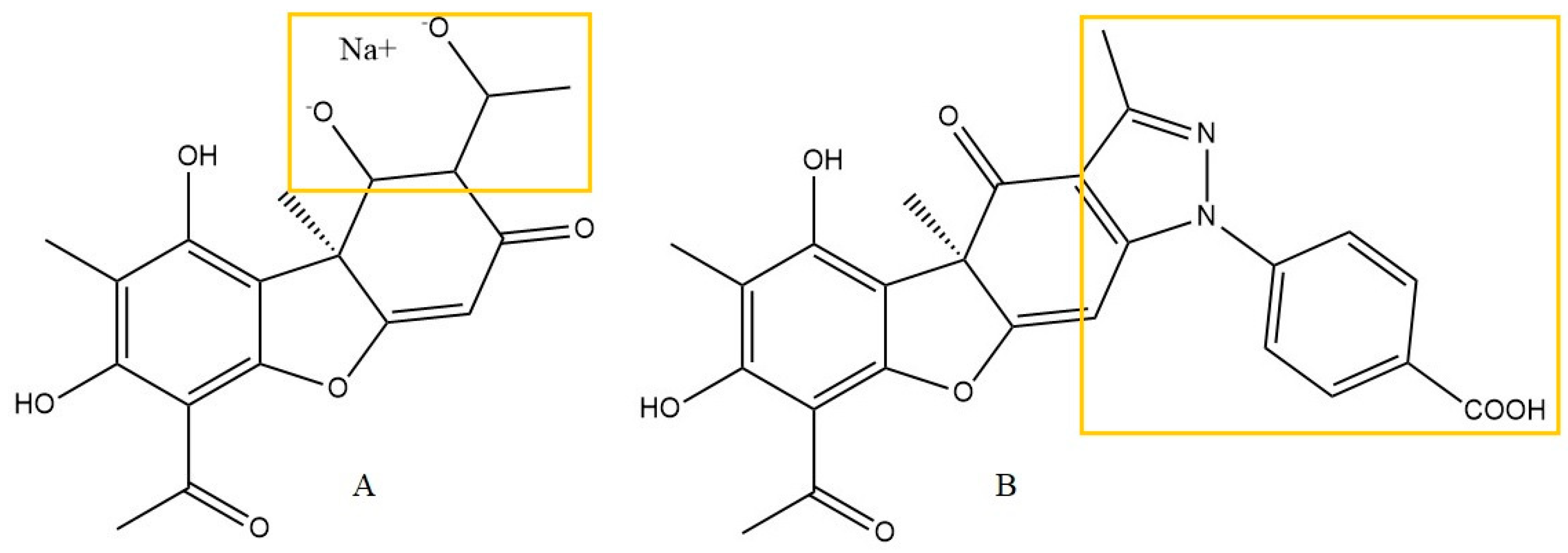
| microglia BV2 cells | UA derivative No 30 (2.5, 5, 10 μM) Reference: sodium usnate 10 μM Different groups: LPS-stimulated cells Methods: Griess reagent (NO) |
| [19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paździora, W.; Podolak, I.; Grudzińska, M.; Paśko, P.; Grabowska, K.; Galanty, A. Critical Assessment of the Anti-Inflammatory Potential of Usnic Acid and Its Derivatives—A Review. Life 2023, 13, 1046. https://doi.org/10.3390/life13041046
Paździora W, Podolak I, Grudzińska M, Paśko P, Grabowska K, Galanty A. Critical Assessment of the Anti-Inflammatory Potential of Usnic Acid and Its Derivatives—A Review. Life. 2023; 13(4):1046. https://doi.org/10.3390/life13041046
Chicago/Turabian StylePaździora, Wojciech, Irma Podolak, Marta Grudzińska, Paweł Paśko, Karolina Grabowska, and Agnieszka Galanty. 2023. "Critical Assessment of the Anti-Inflammatory Potential of Usnic Acid and Its Derivatives—A Review" Life 13, no. 4: 1046. https://doi.org/10.3390/life13041046
APA StylePaździora, W., Podolak, I., Grudzińska, M., Paśko, P., Grabowska, K., & Galanty, A. (2023). Critical Assessment of the Anti-Inflammatory Potential of Usnic Acid and Its Derivatives—A Review. Life, 13(4), 1046. https://doi.org/10.3390/life13041046