

Helixor-M Suppresses Immunostimulatory Activity through TLR4-Dependent NF-κB Pathway in RAW 264.7 Cells

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Measurement of Cell Viability
2.4. Immunofluorescence Assay
2.5. Western Blot
2.6. Real Time PCR (RT-PCR)
2.7. Determination of Nitric Oxide (NO) Production
2.8. Measurement of Phagocytic Activity
2.9. Statistical Analysis
3. Results
3.1. Effects of Helixor-M on RAW 264.7 Cell Viability and Phagocytosis Activity
3.2. Helixor-M Inhibited NO Production and Cyclooxygenase-2 (COX2) Expression in LPS-Induced Raw 264.7 Cells
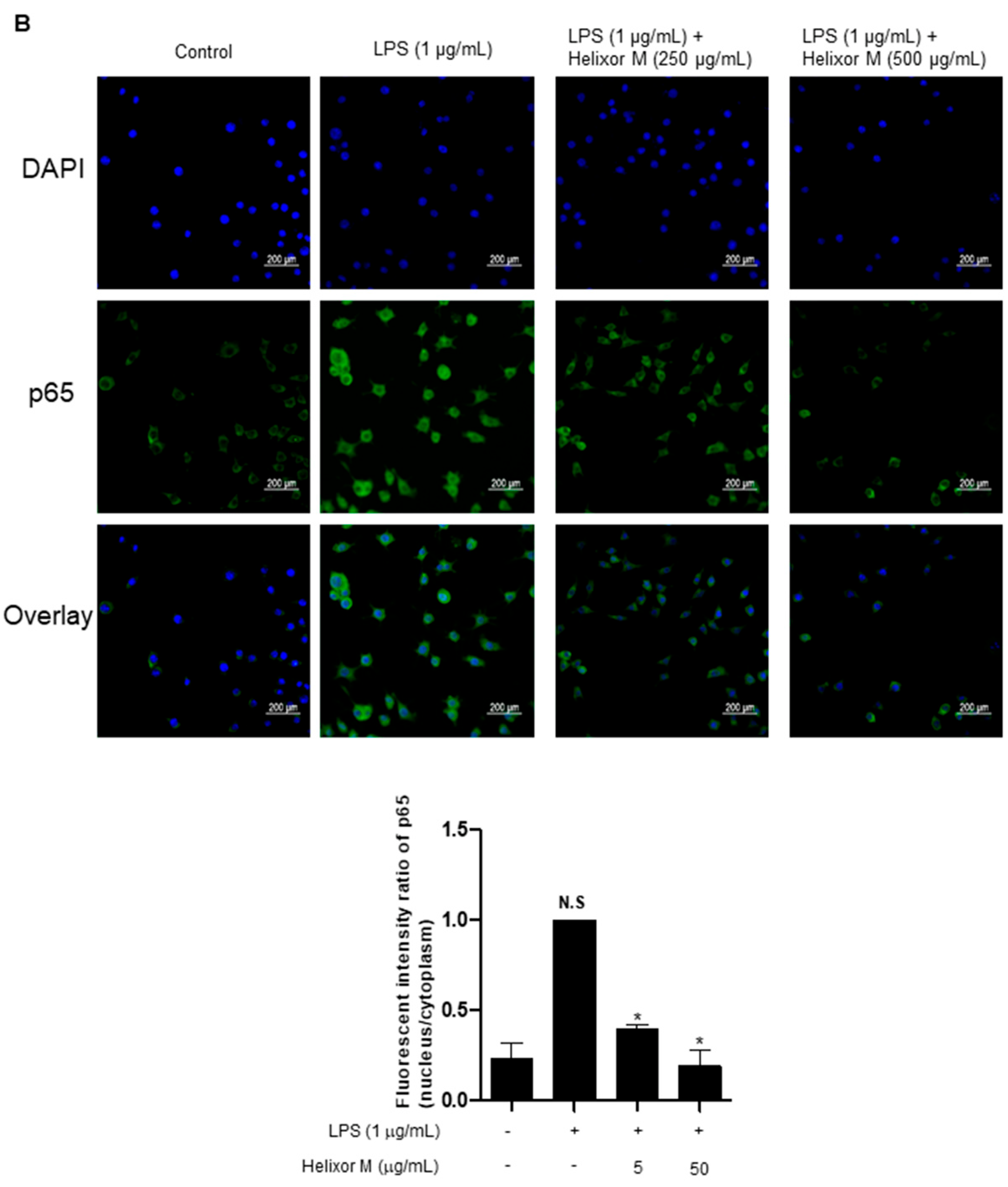
3.3. Effect of Helixor-M on NF-κB Signaling Pathway in LPS-Induced RAW 264.7 Cells
3.4. Effect of Helixor-M on MAPKs in LPS-Induced RAW 264.7 Cells
3.5. Helixor-M Inhibited Pro-Inflammatory Cytokine Mediators in RAW 264.7 Cells
3.6. Effect of Helixor-M on PI3K/AKT Pathway in RAW 264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerriero, J.L. Macrophages: Their Untold Story in T Cell Activation and Function. Int. Rev. Cell Mol. Biol. 2019, 342, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gu, J.; Zong, M.; Zhang, Q.; Li, H.; Li, D.; Mou, X.; Liu, P.; Liu, Y.; Qiu, F.; et al. Anti-inflammatory action of physalin A by blocking the activation of NF-κB signaling pathway. J. Ethnopharmacol. 2021, 267, 113490. [Google Scholar] [CrossRef] [PubMed]
- Pudla, M.; Srisaowakarn, C.; Utaisincharoen, P. NLRP12 negatively modulates inducible nitric oxide synthase (iNOS) expression and tumor necrosis factor-α production in Porphyromonas gingivalis LPS-treated mouse macrophage cell line (RAW264.7). Inflamm. Res. 2019, 68, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, C.; Tang, K.; Dong, X.; Xue, L.; Su, G.; Zhang, W.; Jin, Y. Aquaporin-1 attenuates macrophage-mediated inflammatory responses by inhibiting p38 mitogen-activated protein kinase activation in lipopolysaccharide-induced acute kidney injury. Inflamm. Res. 2019, 68, 1035–1047. [Google Scholar] [CrossRef]
- Hirayama, D.; Iida, T. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef]
- Daliri, E.B.; Choi, S.I.; Cho, B.Y.; Jo, H.Y.; Kim, S.H.; Chelliah, R.; Rubab, M.; Kim, J.H.; Oh, H.T.; Lee, O.H.; et al. Biological activities of a garlic-Cirsium setidens Nakai blend fermented with Leuconostoc mesenteroides. Food Sci. Nutr. 2019, 7, 2024–2032. [Google Scholar] [CrossRef]
- Bordet, E.; Frétaud, M.; Crisci, E.; Bouguyon, E.; Rault, S.; Pezant, J.; Pleau, A.; Renson, P.; Giuffra, E.; Larcher, T.; et al. Macrophage-B Cell Interactions in the Inverted Porcine Lymph Node and Their Response to Porcine Reproductive and Respiratory Syndrome Virus. Front. Immunol. 2019, 10, 953. [Google Scholar] [CrossRef]
- Craxton, A.; Magaletti, D.; Ryan, E.J.; Clark, E.A. Macrophage- and dendritic cell--dependent regulation of human B-cell proliferation requires the TNF family ligand BAFF. Blood 2003, 101, 4464–4471. [Google Scholar] [CrossRef]
- Liu, S.; Yang, T.; Ming, T.W.; Gaun, T.K.W.; Zhou, T.; Wang, S.; Ye, B. Isosteroid alkaloids with different chemical structures from Fritillariae cirrhosae bulbus alleviate LPS-induced inflammatory response in RAW 264.7 cells by MAPK signaling pathway. Int. Immunopharmacol. 2020, 78, 106047. [Google Scholar] [CrossRef]
- Hu, T.Y.; Ju, J.M.; Mo, L.H.; Ma, L.; Hu, W.H.; You, R.R.; Chen, X.Q.; Chen, Y.Y.; Liu, Z.Q.; Qiu, S.Q.; et al. Anti-inflammation action of xanthones from Swertia chirayita by regulating COX-2/NF-κB/MAPKs/Akt signaling pathways in RAW 264.7 macrophage cells. Phytomedicine 2019, 55, 214–221. [Google Scholar] [CrossRef]
- Zheng, Y.; Tian, C.; Fan, C.; Xu, N.; Xiao, J.; Zhao, X.; Lu, Z.; Cao, H.; Liu, J.; Yu, L. Sheng-Mai Yin exerts anti-inflammatory effects on RAW 264.7 cells and zebrafish. J. Ethnopharmacol. 2021, 267, 113497. [Google Scholar] [CrossRef]
- Fang, Y.; Yang, L.; He, J. Plantanone C attenuates LPS-stimulated inflammation by inhibiting NF-κB/iNOS/COX-2/MAPKs/Akt pathways in RAW 264.7 macrophages. Biomed. Pharmacother. = Biomed. Pharmacother. 2021, 143, 112104. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Hua, K.-F.; Lin, C.-C.; Lin, C.-H.; Hsu, J.; Wong, C.-H. Extract of Reishi polysaccharides induces cytokine expression via TLR4-modulated protein kinase signaling pathways. J. Immunol. 2004, 173, 5989–5999. [Google Scholar] [CrossRef]
- Wang, Y.-Q.; Mao, J.-B.; Zhou, M.-Q.; Jin, Y.-W.; Lou, C.-H.; Dong, Y.; Shou, D.; Hu, Y.; Yang, B.; Jin, C.-Y.; et al. Polysaccharide from Phellinus Igniarius activates TLR4-mediated signaling pathways in macrophages and shows immune adjuvant activity in mice. Int. J. Biol. Macromol. 2019, 123, 157–166. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Dong, C.; Davis, R.J.; Flavell, R.A. MAP kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef]
- Li, Y.; Meng, T.; Hao, N.; Tao, H.; Zou, S.; Li, M.; Ming, P.; Ding, H.; Dong, J.; Feng, S.; et al. Immune regulation mechanism of Astragaloside IV on RAW264.7 cells through activating the NF-κB/MAPK signaling pathway. Int. Immunopharmacol. 2017, 49, 38–49. [Google Scholar] [CrossRef]
- Ren, J.; Li, L.; Wang, Y.; Zhai, J.; Chen, G.; Hu, K. Gambogic acid induces heme oxygenase-1 through Nrf2 signaling pathway and inhibits NF-κB and MAPK activation to reduce inflammation in LPS-activated RAW264.7 cells. Biomed. Pharmacother. 2019, 109, 555–562. [Google Scholar] [CrossRef]
- Huber, R.; Schlodder, D.; Effertz, C.; Rieger, S.; Tröger, W. Safety of intravenously applied mistletoe extract-results from a phase I dose escalation study in patients with advanced cancer. BMC Complement. Altern. Med. 2017, 17, 465. [Google Scholar] [CrossRef]
- Lev, E.; Ephraim, M.; Ben-Arye, E. European and Oriental mistletoe: From mythology to contemporary integrative cancer care. Eur. J. Integr. Med. 2011, 3, e133–e137. [Google Scholar] [CrossRef]
- Tu, Y. Artemisinin—A gift from traditional Chinese medicine to the world (Nobel lecture). Angew. Chem. Int. Ed. 2016, 55, 10210–10226. [Google Scholar] [CrossRef] [PubMed]
- Horneber, M.A.; Bueschel, G.; Huber, R.; Linde, K.; Rostock, M. Mistletoe therapy in oncology. Cochrane Database Syst. Rev. 2008, 2008, Cd003297. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.L.; Axtner, J. Adverse Drug Reactions and Expected Effects to Therapy with Subcutaneous Mistletoe Extracts (Viscum album L.) in Cancer Patients. Evid. -Based Complement. Altern. Med. 2014, 2014, 724258. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Na, K.J.; Kim, D.W.; Choi, Y.E.; Ma, J.S.; Jeong, I.S. Chemical pleurodesis using a Viscum album extract in infants with congenital chylothorax. Eur. J. Pediatr. 2014, 173, 823–826. [Google Scholar] [CrossRef]
- Lee, Y.-G.; Jung, I.; Koo, D.-H.; Kang, D.-Y.; Oh, T.Y.; Oh, S.; Lee, S.-S. Efficacy and safety of Viscum album extract (Helixor-M) to treat malignant pleural effusion in patients with lung cancer. Support. Care Cancer 2019, 27, 1945–1949. [Google Scholar] [CrossRef]
- An, E.J.; Kim, Y.; Lee, S.H.; Ko, H.M.; Chung, W.S.; Jang, H.J. Anti-Cancer Potential of Oxialis obtriangulata in Pancreatic Cancer Cell through Regulation of the ERK/Src/STAT3-Mediated Pathway. Molecules 2020, 25, 2301. [Google Scholar] [CrossRef]
- Park, D.; Ko, H.M.; Jee, W.; Jung, J.H.; Kwon, S.W.; Jung, W.S.; Jang, H.-J. Identification of allergens in Acorus gramineus using protein analysis. Mol. Cell. Toxicol. 2022. [Google Scholar] [CrossRef]
- Ko, H.M.; Choi, S.H.; Kim, Y.; An, E.J.; Lee, S.H.; Kim, K. Effect of Rosa laevigata on PM10-Induced Inflammatory Response of Human Lung Epithelial Cells. Evid. Based Complement. Altern. Med. 2020, 2020, 2893609. [Google Scholar] [CrossRef]
- Park, J.; Kim, H.; Lee, I.S.; Kim, K.H.; Kim, Y.; Na, Y.C.; Lee, J.H.; Jang, H.J. The therapeutic effects of Yongdamsagan-tang on autoimmune hepatitis models. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 94, 244–255. [Google Scholar] [CrossRef]
- Chiu, C.-H.; Tong, Y.-W.; Yu, J.-F.; Lei, K.F.; Chen, A.C.-Y. Osteogenesis and Chondrogenesis of Primary Rabbit Periosteal Cells under Non-uniform 2-Axial Tensile Strain. Biochip J. 2020, 14, 438–446. [Google Scholar] [CrossRef]
- Yoo, Y.-J.; Huh, S.-E.; Kim, Y.; Jang, H.-J. Anti-cancer activity of Boswellia carterii extract alters the stress functional gene expression in the pancreatic cancer cell. Biochip J. 2019, 13, 191–201. [Google Scholar] [CrossRef]
- Yeom, S.-J.; Lee, S.-Y.; Tas, I.; Lee, M.; Park, S.-Y.; Jung, S.-C.; Kim, K.K.; Kim, H. Microquantitation of van gogh-like protein 1 by using antibody-conjugated magnetic beads. Biochip J. 2019, 13, 151–157. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, M. A Simple and Multiplex Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid Detection of SARS-CoV. BioChip J. 2019, 13, 341–351. [Google Scholar] [CrossRef]
- Liu, C.; Chen, J.; Chen, L.; Huang, X.; Cheung, P.C. Immunomodulatory activity of polysaccharide–protein complex from the mushroom sclerotia of Polyporus rhinocerus in murine macrophages. J. Agric. Food Chem. 2016, 64, 3206–3214. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef]
- Guo, C.; Yang, L.; Luo, J.; Zhang, C.; Xia, Y.; Ma, T.; Kong, L. Sophoraflavanone G from Sophora alopecuroides inhibits lipopolysaccharide-induced inflammation in RAW264.7 cells by targeting PI3K/Akt, JAK/STAT and Nrf2/HO-1 pathways. Int. Immunopharmacol. 2016, 38, 349–356. [Google Scholar] [CrossRef]
- Chu, A.J. Antagonism by bioactive polyphenols against inflammation: A systematic view. Inflamm. Allergy Drug Targets 2014, 13, 34–64. [Google Scholar] [CrossRef]
- Paramonov, A.S.; Lyukmanova, E.N.; Tonevitsky, A.G.; Arseniev, A.S.; Shenkarev, Z.O. Spatial structure and oligomerization of viscotoxin A3 in detergent micelles: Implication for mechanisms of ion channel formation and membrane lysis. Biochem. Biophys. Res. Commun. 2021, 585, 22–28. [Google Scholar] [CrossRef]
- Orrù, S.; Scaloni, A.; Giannattasio, M.; Urech, K.; Pucci, P.; Schaller, G. Amino acid sequence, SS bridge arrangement and distribution in plant tissues of thionins from Viscum album. Biol. Chem. 1997, 378, 989–996. [Google Scholar] [CrossRef]
- Schaller, G.; Urech, K.; Giannattasio, M. Cytotoxicity of different viscotoxins and extracts from the European subspecies of Viscum album L. Phytother. Res. 1996, 10, 473–477. [Google Scholar] [CrossRef]
- Tabiasco, J.; Pont, F.; Fournié, J.J.; Vercellone, A. Mistletoe viscotoxins increase natural killer cell-mediated cytotoxicity. Eur. J. Biochem. 2002, 269, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K.; Taylor, M.E. Biology of animal lectins. Annu. Rev. Cell Biol. 1993, 9, 237–264. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.L.; Megson, I.L.; Haslett, C.; Rossi, A.G. Nitric oxide: A key regulator of myeloid inflammatory cell apoptosis. Cell Death Differ. 2003, 10, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.; Fernando, I.P.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef]
- Welsh, P.; Grassia, G.; Botha, S.; Sattar, N.; Maffia, P. Targeting inflammation to reduce cardiovascular disease risk: A realistic clinical prospect? Br. J. Pharmacol. 2017, 174, 3898–3913. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene 2010, 29, 781–788. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, X.H.; Chen, H.; Guo, J.H.; Tsang, L.L.; Yu, M.K.; Xu, W.M.; Chan, H.C. CFTR negatively regulates cyclooxygenase-2-PGE2 positive feedback loop in inflammation. J. Cell. Physiol. 2012, 227, 2759–2766. [Google Scholar] [CrossRef]
- Debprasad, C.; Hemanta, M.; Paromita, B.; Durbadal, O.; Kumar, K.A.; Shanta, D.; Kumar, H.P.; Tapan, C.; Ashoke, S.; Sekhar, C. Inhibition of, TNF-and iNOS EXpression by Shorea robusta L.: An Ethnomedicine Used for Anti-Inflammatory and Analgesic Activity. Evid. Based Complement. Altern. Med. 2012, 2012, 254849. [Google Scholar] [CrossRef]
- Hernandez, A.; Patil, N.K.; Stothers, C.L.; Luan, L.; McBride, M.A.; Owen, A.M.; Burelbach, K.R.; Williams, D.L.; Sherwood, E.R.; Bohannon, J.K. Immunobiology and application of toll-like receptor 4 agonists to augment host resistance to infection. Pharmacol. Res. 2019, 150, 104502. [Google Scholar] [CrossRef]
- Doyle, S.L.; O’Neill, L.A. Toll-like receptors: From the discovery of NFκB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006, 72, 1102–1113. [Google Scholar] [CrossRef]
- Rigoglou, S.; Papavassiliou, A.G. The NF-κB signalling pathway in osteoarthritis. Int. J. Biochem. Cell Biol. 2013, 45, 2580–2584. [Google Scholar] [CrossRef]
- Schuliga, M. NF-kappaB signaling in chronic inflammatory airway disease. Biomolecules 2015, 5, 1266–1283. [Google Scholar] [CrossRef]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef]
- LoRusso, P.M. Inhibition of the PI3K/AKT/mTOR pathway in solid tumors. J. Clin. Oncol. 2016, 34, 3803. [Google Scholar] [CrossRef]
- Tokuhira, N.; Kitagishi, Y.; Suzuki, M.; Minami, A.; Nakanishi, A.; Ono, Y.; Kobayashi, K.; Matsuda, S.; Ogura, Y. PI3K/AKT/PTEN pathway as a target for Crohn’s disease therapy. Int. J. Mol. Med. 2015, 35, 10–16. [Google Scholar] [CrossRef]
- Zheng, T.; Shi, Y.; Zhang, J.; Peng, J.; Zhang, X.; Chen, K.; Chen, Y.; Liu, L. MiR-130a exerts neuroprotective effects against ischemic stroke through PTEN/PI3K/AKT pathway. Biomed. Pharmacother. 2019, 117, 109117. [Google Scholar] [CrossRef]
- Liu, F.; Wen, Y.; Kang, J.; Wei, C.; Wang, M.; Zheng, Z.; Peng, J. Regulation of TLR4 expression mediates the attenuating effect of erythropoietin on inflammation and myocardial fibrosis in rat heart. Int. J. Mol. Med. 2018, 42, 1436–1444. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′=3′) | Reverse Primer Sequence (5′=3′) |
|---|---|---|
| TNF-α | (F) 5′-GCAGGTCTACTTTGGGTCATTG-3′ | (R) 5′-GCGTTTGGGAAGGTTGGA-3′ |
| IL-1β | (F) 5′-TCAGCCAATCTTCATTGCTCAA-3′ | (R) 5′-TGGCGAGCTCAGGTAC-TTCTG-3′ |
| IL-6 | (F) 5′-AGGGCTCTTCGGCAAATGTA-3′ | (R) 5′-GAAGGAATGCCCATTAACAACAA-3′ |
| TLR4 | (F) 5′-TAGCCATTGCTGCCAACATC-3′ | (R) 5′-GCCTCAGCAGGGACTTCT CAA-3′ |
| GAPDH | (F) 5′-GCCACATCGCTCAGACACC-3′ | (R) 5′-CCCAATACGACCAAATCCGT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, D.; Ko, H.M.; Jee, W.; Park, S.M.; Park, Y.R.; Jung, J.H.; Kim, H.S.; Chung, W.S.; Kim, S.K.; Chung, J.S.; et al. Helixor-M Suppresses Immunostimulatory Activity through TLR4-Dependent NF-κB Pathway in RAW 264.7 Cells. Life 2023, 13, 595. https://doi.org/10.3390/life13020595
Park D, Ko HM, Jee W, Park SM, Park YR, Jung JH, Kim HS, Chung WS, Kim SK, Chung JS, et al. Helixor-M Suppresses Immunostimulatory Activity through TLR4-Dependent NF-κB Pathway in RAW 264.7 Cells. Life. 2023; 13(2):595. https://doi.org/10.3390/life13020595
Chicago/Turabian StylePark, Doil, Hyun Min Ko, Wona Jee, So Mi Park, Ye Rin Park, Ji Hoon Jung, Hyung Suk Kim, Won Seok Chung, Sang Ki Kim, Jong Sup Chung, and et al. 2023. "Helixor-M Suppresses Immunostimulatory Activity through TLR4-Dependent NF-κB Pathway in RAW 264.7 Cells" Life 13, no. 2: 595. https://doi.org/10.3390/life13020595
APA StylePark, D., Ko, H. M., Jee, W., Park, S. M., Park, Y. R., Jung, J. H., Kim, H. S., Chung, W. S., Kim, S. K., Chung, J. S., & Jang, H. J. (2023). Helixor-M Suppresses Immunostimulatory Activity through TLR4-Dependent NF-κB Pathway in RAW 264.7 Cells. Life, 13(2), 595. https://doi.org/10.3390/life13020595