


Antioxidant and Antidiabetic Activity of Algae

 ,
,  ,
,  and
and
Abstract
1. Introduction
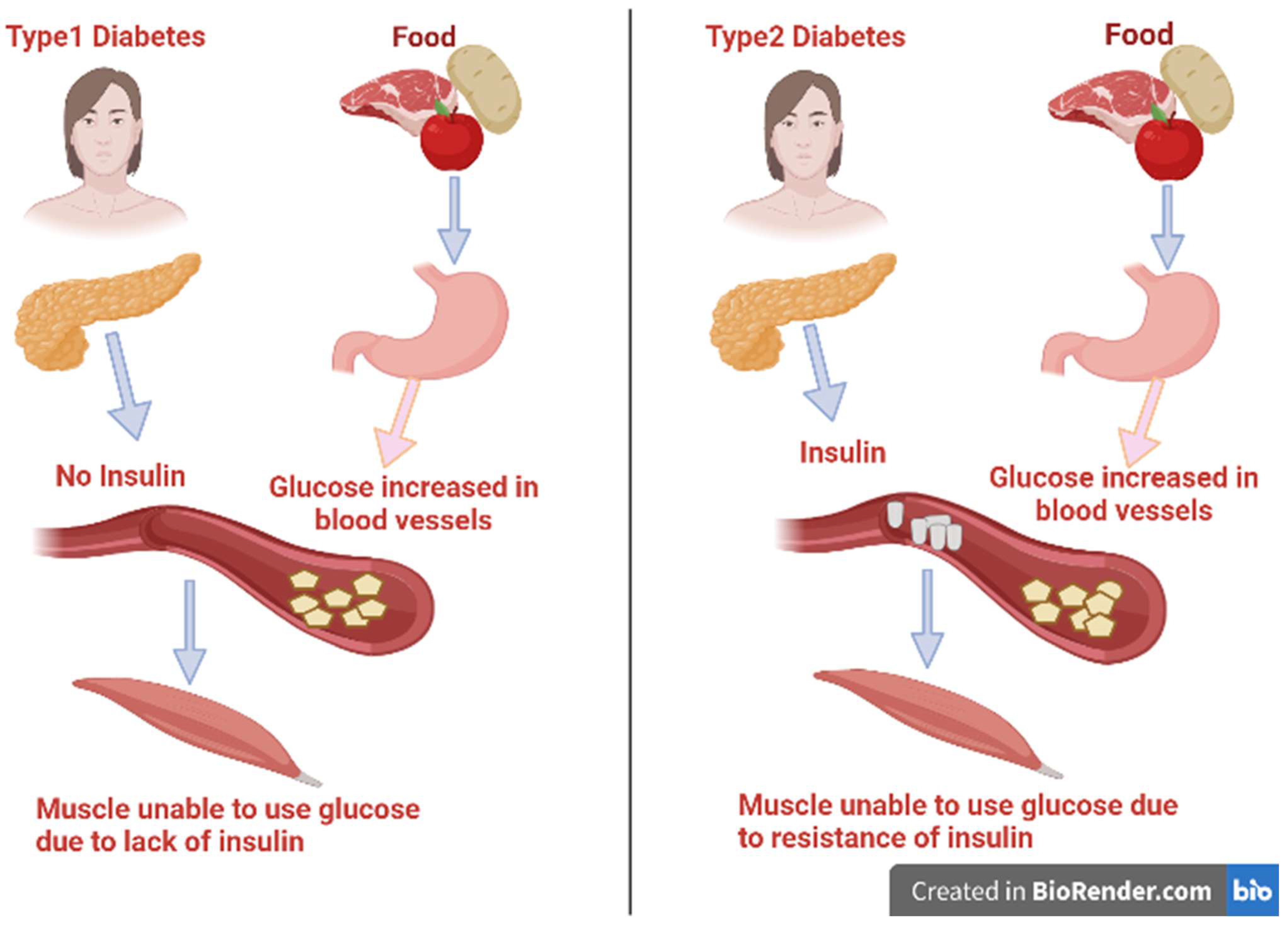
2. Factors That Contribute to Diabetes and Its Complications
Oxidative Stress Linked to Diabetes
3. Treatment of Diabetes
3.1. Antioxidants
3.2. Algal Extracts and Their Bioactive Components Linked to Diabetes Treatment
3.3. Antioxidant and Antihyperglycemic Activity of Different Algal Extracts
3.4. The Inhibitory Activity of Different Algal Extracts on the Carbohydrate Hydrolyzing Enzymes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algal Species | Division | Extract Type | α-Amylase Inhibition % | IC50 Value | α-Glucosidase Inhibition % | IC50 Value | Ref. |
|---|---|---|---|---|---|---|---|
| Spirulina sp. | Cyanobacteria | Butanol crude extract | - | - | - | 23 μg/mL | [61] |
| Pseudanabaena sp. | Cyanobacteria | Exopolysaccharides extract | - | - | 14.02% | - | [88] |
| Fischerella sp. | Cyanobacteria | Aqueous crude extract | - | - | 7.5% | - | [90] |
| Phaeodactylum tricornutum | Bacillariophyta | Fucoxanthin extract. | - | 0.68 mmol/L | - | 4.75 mmol/L | [91] |
| Nannochloropsis oculata | Ochrophyta | Ethyl acetate crude extract | 78.52% | 121.96 μg/mL | 80.42% | 178.53 μg/mL | [92] |
| Arthrospira platensis | Cyanobacteria | Methanol crude extract | 96.46% | 13.31 mg/mL | 97.42% | 9.56 mg/mL | [57] |
| Porphyridium sp. | Rhodophyta | Methanol crude extract | - | - | 12.63%. | - | [93] |
| Algal Species | Division | Extract Type | α-Amylase Inhibition % | IC50 | α-Glucosidase Inhibition % | IC50 | Ref. |
|---|---|---|---|---|---|---|---|
| Ulva reticulata | Chlorophyta | Ethanol crude extract | 89.1% | - | 79.55% | - | [25] |
| Sargassum polycystum | Phaeophyta | Ethyl acetate | - | 438.5 μg/mL | - | 289.7 μg/mL | [28] |
| Sargassum wightii | Phaeophyta | Acetone crude extract | - | 438.5 μg/mL | - | 289.7 μg/mL | [28] |
| Hormophysa cuneiformis | Phaeophyta | Methanol crude extract | - | - | 53% | 676.9 μg/mL | [100] |
| Fucus vesiculosus | Phaeophyta | Semi-purified phlorotannin fraction | - | 2.8 μg/mL | - | 0.82 μg/mL | [101] |
| Turbinaria decurrens | Phaeophyta | Methanol crude extract | - | - | - | 11 μg/mL | [75] |
| Turbinaria decurrens | Phaeophyta | Acetone crude extract | 96.1% | 4.37 mg/mL | 97.4% | 2.84 mg/mL | [58] |
| Taonia atomaria | Phaeophyta | Ethanol crude extract | 66.3% | - | - | - | [77] |
| Laurencia dendroidea | Rhodophyta | Ethyl acetate fraction | - | - | - | 8.14 μg/mL | [81] |
| Halymenia durvillei | Rhodophyta | Aqueous crude extract | - | - | - | 4.34 mg/mL | [103] |
3.5. In Vivo Antihyperglycemic Activity of Different Algal Extracts
3.5.1. Effect of Different Algal Extracts as Treatment on the Blood Glucose Levels of Diabetic-Induced Animals
3.5.2. Effect of Different Algal Extracts as Treatment on the Body Weight of Diabetic-Induced Animals
3.5.3. Effect of Different Algal Extracts as Treatment on the Hemoglobin (Hb) Levels of Diabetic-Induced Animals
3.5.4. Effect of Different Algal Extracts as Treatment on the Total Bilirubin of Diabetic-induced Animals
3.5.5. Effect of Different Algal Extracts as Treatment on the Liver Enzymes of Diabetic-Induced Animals
3.5.6. Effect of Different algal Extracts as Treatment on the Urea and Creatinine of Diabetic-Induced Animals
3.5.7. Effect of Different Algal Extracts as Treatment on the Total Protein of Diabetic-Induced Animals
3.5.8. Effect of Different Algal Extracts as Treatment on the Albumin Level of Diabetic-Induced Animals
3.5.9. Effect of Different Algal Extracts as Treatment on the Lipid Profile of Diabetic-Induced Animals
3.5.10. Effect of Different Algal Extracts as Treatment on the Histological Profile of the Liver and Pancreas of the Diabetic-Induced Animals
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Diabetes Federation (IDF). IDF Diabetes Atlas. 2022. Available online: https://www.diabetesatlas.org/ (accessed on 31 January 2023).
- WHO. W.H.O. Diabetes. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 31 January 2023).
- Ritu, M.; Nandini, J. Nutritional composition of Stevia rebaudiana, a sweet herb, and its hypoglycaemic and hypolipidaemic effect on patients with non-insulin dependent diabetes mellitus. J. Sci. Food Agric. 2016, 96, 4231–4234. [Google Scholar] [CrossRef] [PubMed]
- Akpaso, M.I.; Igiri, A.O.; Odey, P.A. A comparative study on the effect of combined methanolic leaf extracts of Vernonia amygdalina and Gongronema latifolium and metformin on the pancreatic beta cells of streptozocin induced diabetic wistar rats. Asian J. Pharm. Nurs. Med. Sci. 2017, 5, 42–48. [Google Scholar] [CrossRef]
- American Diabetes Association. Classification and diagnosis of diabetes: Standards of medical care in diabetes. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C. Standards of Medical Care for Patients with Diabetes; American Family Physician: Leawood, KS, USA, 2006; Volume 74. [Google Scholar]
- Fox, I.S. Human Physiology, 8th ed.; McGraw-Hill: New York, NY, USA, 2004. [Google Scholar]
- Vaidyanathan, K.; Vasudevan, D.M.; Sreekumari, S. Regulation of blood glucose, insulin and diabetes mellitus. In Textbook of Biochemistry for Medical Students; Jaypee Brothers Medical Publishers: New Delhi, India, 2011; pp. 285–290. [Google Scholar]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef]
- Franks, P.W.; McCarthy, M.I. Exposing the exposures responsible for type 2 diabetes and obesity. Science 2016, 354, 69–73. [Google Scholar] [CrossRef]
- Sørensen, L.E.; Jeppesen, P.B.; Christiansen, C.B.; Hermansen, K.; Gregersen, S. Nordic seaweed and diabetes prevention: Exploratory studies in KK-Ay mice. Nutrients 2019, 11, 1435. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Tiwari, A. Imbalance in antioxidant defense and human disease: Multiple approach of natural antioxidant therapy. J. Curr. Sci. 2001, 81, 1179–1187. [Google Scholar]
- Pirian, K.; Moein, S.; Sohrabipour, J.; Rabiei, R.; Blomster, J. Antidiabetic and antioxidant activities of brown and red macroalgae from the Persian Gulf. J. Appl. Phycol. 2017, 29, 3151–3159. [Google Scholar] [CrossRef]
- Poljsak, B.; Fink, R. The protective role of antioxidants in the defence against ROS/RNS-mediated environmental pollution. Oxidative Med. Cell. Longev. 2014, 2014, 22–28. [Google Scholar] [CrossRef]
- Shin, C.S.; Moon, B.S.; Park, K.S.; Kim, S.Y.; Park, S.J.; Chung, M.H.; Lee, H.K. Serum 8-hydroxy-guanine levels are increased in diabetic patients. Diabetes Care 2001, 24, 733–737. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.Y.; Fung, P.L.; Xiaoqiang, Y.; Zhen-Yu, C.; Yu, H. Reactive oxygen species in vascular wall. Cardiovasc. Hematol. Disord. -Drug Targets 2006, 6, 1–19. [Google Scholar]
- Styskal, J.; Van Remmen, H.; Richardson, A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free. Radic. Biol. Med. 2012, 52, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, S.; Khan, A. Antioxidants and diabetes. Indian J. Endocrinol. Metab. 2012, 16, S267–S271. [Google Scholar] [CrossRef] [PubMed]
- Desco, M.-C.; Asensi, M.; Márquez, R.; Martínez-Valls, J.; Vento, M.; Pallardó, F.V.; Sastre, J.; Viña, J. Xanthine oxidase is involved in free radical production in type 1 diabetes. Diabetes 2002, 51, 1118–1124. [Google Scholar] [CrossRef]
- Maiese, K. New insights for oxidative stress and diabetes mellitus. Oxidative Med. Cell. Longev. 2015, 2015, 875961. [Google Scholar] [CrossRef]
- Xin, Y.; Yuan, B.; Yu, B.; Wang, Y.; Wu, J.; Zhou, W.; Qiu, Z. Tet1-mediated DNA demethylation regulates neuronal cell death induced by oxidative stress. Sci. Rep. 2015, 5, 7645–7650. [Google Scholar] [CrossRef]
- Maiese, K. MTOR: Driving apoptosis and autophagy for neurocardiac complications of diabetes mellitus. World J. Diabetes 2015, 6, 217–224. [Google Scholar] [CrossRef]
- Reka, P.; Banu, T.; Seethalakshmi, M. Alpha amylase and α glucosidase inhibition activity of selected edible seaweeds from South Coast area of India. Int. J. Pharm. Pharm. Sci. 2017, 9, 64–68. [Google Scholar] [CrossRef]
- Rang, H.; Dale, M.; Ritter, J.; Moore, P. Pharmacology, 5th ed.; Churchill Livingstone: London, UK, 2003. [Google Scholar]
- Kwon, Y.I.; Apostolidis, E.; Kim, Y.C.; Shetty, K. Health benefits of traditional corn, beans, and pumpkin: In vitro studies for hyperglycemia and hypertension management. J. Med. Food 2007, 10, 266–275. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Antidiabetic potential of marine algae by inhibiting key metabolic enzymes. Front. Life Sci. 2015, 8, 148–159. [Google Scholar] [CrossRef]
- Bhandari, M.R.; Jong-Anurakkun, N.; Hong, G.; Kawabata, J. α-Glucosidase and α-amylase inhibitory activities of Nepalese medicinal herb Pakhanbhed (Bergenia ciliata, Haw.). Food Chem. 2008, 106, 247–252. [Google Scholar] [CrossRef]
- Fernando, M.R.; Wickramasinghe, S.M.D.N.; Thabrew, M.I.; Ariyananda, P.L.; Karunanayake, E.H. Effect of Artocarpus heterophyllus and Asteracanthus longifolia on glucose tolerance in normal human subjects and in maturity-onset diabetic patients. J. Ethnopharmacol. 1991, 31, 277–282. [Google Scholar] [CrossRef]
- Lee, M.Y.; Choi, D.S.; Lee, M.K.; Lee, H.W.; Park, T.S.; Kim, D.M.; Chung, C.H.; Kim, D.K.; Kim, I.J.; Jang, H.C.; et al. Comparison of acarbose and voglibose in diabetes patients who are inadequately controlled with basal insulin treatment: Randomized, parallel, open-label, active-controlled study. J. Korean Med. Sci. 2014, 29, 90–97. [Google Scholar] [CrossRef]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. Inhibitory potential of wine and tea against α-amylase and α-glucosidase for management of hyperglycemia linked to type 2 diabetes. J. Food Biochem. 2008, 32, 15–31. [Google Scholar] [CrossRef]
- Lee, C.W.; Han, J.S. Hypoglycemic effect of Sargassum ringgoldianum extract in STZ-induced diabetic mice. Prev. Nutr. Food Sci. 2012, 17, 8–13. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Hama, Y.; Miyasaki, T.; Ito, M.; Nakamura, T. Extracellular secretion of phenolic substances from living brown algae. J. Appl. Phycol. 2006, 18, 787–794. [Google Scholar] [CrossRef]
- Desai, K.; Sivakami, S. Spirulina: The wonder food of the 21st Century. Asia-Pac. Biotech News 2004, 08, 1298–1302. [Google Scholar] [CrossRef]
- Wanasundara, P.; Shahidi, F. Antioxidants: Science, Technology, and Applications, 6th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; Volume 6. [Google Scholar]
- Wijesekara, I.; Senevirathne, M.; Li, Y.; Kim, S. Functional ingredients from marine algae as potential antioxidants in the food industry. In Handbook of Marine Macroalgae; Wiley Online Books: Hoboken, NJ, USA, 2011; pp. 398–402. [Google Scholar]
- Goiris, K.; De Vreese, P.; De Cooman, L.; Muylaert, K. Rapid screening and guided extraction of antioxidants from microalgae using voltammetric methods. J. Agric. Food Chem. 2012, 60, 7359–7366. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.; David, B.M. Food Lipids: Chemistry, Nutrition, Biotechnology; Marcel Dekker Inc.: New York, NY, USA, 2002. [Google Scholar]
- Chandini, S.K.; Ganesan, P.; Bhaskar, N. In vitro antioxidant activities of three selected brown seaweeds of India. Food Chem. 2008, 107, 707–713. [Google Scholar] [CrossRef]
- Samaraweera, A.M.; Vidanarachchi, J.K.; Kurukulasuriya, M.S. Industrial Applications of Macroalgae; John Wiley & Sons Ltd: West Sussex, UK, 2012; pp. 500–521. [Google Scholar]
- Motshakeri, M.; Ebrahimi, M.; Goh, Y.M.; Matanjun, P.; Mohamed, S. Sargassum polycystum reduces hyperglycaemia, dyslipidaemia and oxidative stress via increasing insulin sensitivity in a rat model of type 2 diabetes. J. Sci. Food Agric. 2013, 93, 93–1772. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.M.; Shrivastava, R.K.; Singh, N. Status of exogenous antioxidant, total antioxidant capacity and oxidative stress in SCA patients. Indian J. Appl. Res. 2018, 8, 112–118. [Google Scholar]
- Abbott, I.A.; Hollenberg, G.J. Marine Algae of California; Stanford University Press: Redwood City, CA, USA, 1976. [Google Scholar]
- Namikoshi, M.; Rinehart, K.L. Bioactive compounds produced by cyanobacteria. J. Indian Microbiol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Raven, J.A.; Mario, G. Algae. Curr. Biol. 2014, 24, 590–595. [Google Scholar] [CrossRef]
- Dhargalkar, V.K.; Verlecar, X.N. Southern Ocean seaweeds: A resource for exploration in food and drugs. Aquaculture 2009, 287, 229–242. [Google Scholar] [CrossRef]
- Manivannan, K.; Thirumaran, G.; Devi, G.K.; Anantharaman, P.; Balasubramanian, T. Proximate composition of different group of seaweeds from Vedalai Coastal Waters (Gulf of Mannar). Middle-East J. Sci. Res. 2009, 4, 72–77. [Google Scholar]
- Paul, J.P.J. Phytochemical analysis of Padina distromatica Hauck. Indo Am. J. Pharm. Res. 2013, 3, 5290–5297. [Google Scholar]
- Solanki, R.; Khanna, M.; Lal, R. Bioactive compounds from marine actinomycetes. Indian J. Microbiol. 2008, 48, 410–431. [Google Scholar] [CrossRef]
- Valls, R.; Piovetti, L.; Banaigs, B.; Archavlis, A.; Pellegrini, M. (S)-13-hydroxygeranylgeraniol-derived furanoditerpenes from Bifurcaria bifurcate. Phytochemistry 1995, 39, 145–149. [Google Scholar] [CrossRef]
- Okwu, D.E. Improving the nutritive value of Cassava tapioca meal with local spices. J. Nutraceuticals Funct. Med. Foods 2001, 3, 43–50. [Google Scholar] [CrossRef]
- Rivière, C.; Hong, V.N.T.; Pieters, L.; Dejaegher, B.; Heyden, Y.V.; Van, M.C.; Quetin-Leclercq, J. Polyphenols isolated from antiradical extracts of Mallotus metcalfianus. Phytochemistry 2009, 70, 86–94. [Google Scholar] [CrossRef]
- Polterait, O. Antioxidants and free radical scavengers of natural origin. Curr. Org. Chem. 1997, 1, 415–440. [Google Scholar]
- Mandal, P.; Sinha Babu, S.P.; Mandal, N.C. Antimicrobial activity of saponins from Acacia auriculiformis. Fitoterapia 2005, 76, 462–465. [Google Scholar] [CrossRef]
- Gheda, S.F.; Abo-Shady, A.M.; Abdel-Karim, O.H.; Ismail, G.A. Antioxidant and Antihyperglycemic Activity of Arthrospira platensis (Spirulina platensis) Methanolic Extract: In vitro and In vivo Study. Egypt. J. Bot. 2021, 61, 71–93. [Google Scholar] [CrossRef]
- Ismail, G.A.; Gheda, S.F.; Abo-Shady, A.M.; Abdel-Karim, O.H. In vitro potential activity of some seaweeds as antioxidants and inhibitors of diabetic enzymes. Food Sci. Technol. 2020, 40, 681–691. [Google Scholar] [CrossRef]
- Souza, M.M.d.; Prietto, L.; Ribeiro, A.C.; Souza, T.D.d.; Badiale-Furlong, E. Assessment of the antifungal activity of Spirulina platensis phenolic extract against Aspergillus flavus. Ciência e Agrotecnologia 2011, 35, 1050–1058. [Google Scholar] [CrossRef]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar Drugs 2021, 19, 549. [Google Scholar] [CrossRef]
- Gouda, K.G.M.; Kavitha, M.D.; Sarada, R. Antihyperglycemic, antioxidant and antimicrobial activities of the butanol extract from Spirulina platensis. J. Food Biochem. 2015, 39, 594–602. [Google Scholar] [CrossRef]
- Jayshree, A.; Jayashree, S.; Nallamuthu, T. Chlorella vulgaris and Chlamydomonas reinhardtii: Effective antioxidant, antibacterial and anticancer mediators. Indian J. Pharm. Sci. 2016, 78, 575–581. [Google Scholar] [CrossRef]
- Scaglioni, P.T.; Quadros, L.; de Paula, M.; Furlong, V.B.; Abreu, P.C.; Badiale-Furlong, E. Inhibition of enzymatic and oxidative processes by phenolic extracts from Spirulina sp. and Nannochloropsis sp. Food Technol. Biotechnol. 2018, 56, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Karim, O.H.; Gheda, S.F.; Ismail, G.A.; Abo-Shady, A.M. Phytochemical screening and antioxidant activity of Chlorella vulgaris. Delta J. Basic Appl. Sci. 2020, 41, 76–86. [Google Scholar] [CrossRef]
- Santhar, D.T.; Haq, M.A.B.; Marudhupandi, T.; Vaseeharan, B.; Rajan, D.K.; Moovendhan, M. Evaluation of chemical compositions and antioxidant potential of marine microalgae of the genus Nannochloropsis. Biomass Convers. Biorefinery 2021. [Google Scholar] [CrossRef]
- Jaffer, M.; Ashraf, H.; Shaheen, S. Phytochemical, antioxidant and antimicrobial activity of biological important algae Hydrodictyon reticulatum L. Pak. J. Sci. Ind. Res. Ser. b Biol. Sci. 2021, 64, 244–250. [Google Scholar] [CrossRef]
- Moaveni, S.; Salami, M.; Khodadadi, M.; McDougall, M.; Emam-Djomeh, Z. Investigation of S.limacinum microalgae digestibility and production of antioxidant bioactive peptides. LWT 2022, 154, 112468. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C.; Liu, B.; Lin, L.; Sarker, S.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Food Sci. Technol. 2017, 72, 1–12. [Google Scholar] [CrossRef]
- Gunathilaka, T.L.; Keertihirathna, L.R.; Peiris, D. Advanced pharmacological uses of marine algae as an anti-diabetic therapy. In Medicinal Plants from Nature; IntechOpen: London, UK, 2021. [Google Scholar]
- Elangovan, M.; Noorjahan, A.; Anantharaman, P. Extraction Of Metabolites And Screening Their Antioxidant Potential From Marine Macro Algae. Int. J. Sci. Technol. Res. 2019, 8, 1059–1064. [Google Scholar]
- Al-Azzawie, H.F.; Alhamdani, M.-S.S. Hypoglycemic and antioxidant effect of Oleuropein in alloxan-diabetic rabbits. Life Sci. 2006, 78, 1371–1377. [Google Scholar] [CrossRef]
- Ibrahim, R.Y.M.; Saber, A.A.; Hammad, H.B.I. The possible role of the seaweed Ulva fasciata on ameliorating hyperthyroidism-associated heart inflammations in a rat model. Environ. Sci. Pollut. Res. 2021, 28, 6830–6842. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Angiotensin I-Converting Enzyme (ACE) Inhibitory Activity, Antioxidant Properties, Phenolic Content and Amino Acid Profiles of Fucus spiralis L. Protein Hydrolysate Fractions. Mar. Drugs 2017, 15, 311. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, R.; Chitra, L.; Penislusshiyan, S.; Palvannan, T. Exploring bioactive fraction of Sargassum wightii: In vitro elucidation of Angiotensin 1-converting enzyme inhibition and antioxidant potential. Int. J. Food Prop. 2018, 21, 674–684. [Google Scholar] [CrossRef]
- Arguelles, E.D.; Sapin, A.B. In vitro antioxidant, alpha-glucosidase inhibition and antibacterial properties of Turbinaria decurrens Bory (Sargassaceae, Ochrophyta). Asia-Pac. J. Sci. Technol. 2020, 25, 1–9. [Google Scholar]
- El-Sheekh, M.M.; El-Shenody, R.A.E.K.; Bases, E.A.; El Shafay, S.M. Comparative assessment of antioxidant activity and biochemical composition of four seaweeds, Rocky Bay of Abu Qir in Alexandria, Egypt. Food Science and Technology 2021, 41, 29–40. [Google Scholar] [CrossRef]
- Shafay, S.E.L.; El-Sheekh, M.; Bases, E.; El-Shenody, R. Antioxidant, antidiabetic, anti-inflammatory and anticancer potential of some seaweed extracts. Food Sci. Technol. 2021, 42, 1–12. [Google Scholar] [CrossRef]
- Abhishek, D.; Sanjay, S.; Jadeja, B.A. Cytotoxicity, Antioxidant And Antimicrobial Activity Of Marine Macro Algae (Iyengaria Stellata And Padina Boryana) From The Gujarat Coast. J. Maharaja Sayajirao Univ. Baroda 2021, 55, 130–140. [Google Scholar]
- Qian, W.-W.; Yang, S.-Q.; Hu, S.-M.; Wang, X.-L.; Zhu, Y.; Zhou, T. Enzymatic degradation, antioxidant and immunoregulatory activities of polysaccharides from brown algae Sargassum fusiforme. J. Food Meas. Charact. 2021, 15, 1960–1972. [Google Scholar] [CrossRef]
- Čagalj, M.; Skroza, D.; Tabanelli, G.; Özogul, F.; Šimat, V. Maximizing the antioxidant capacity of Padina pavonica by choosing the right drying and extraction methods. Processes 2021, 9, 587. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Nguyen, T.H.; Nguyen, V.M.; Nguyen, T.L.P.; Tran, T.V.A.; Do, A.D.; Kim, S.M. Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 2019, 9, 501–509. [Google Scholar] [CrossRef]
- El Nur, E.E.; Ali, L.I.; Fadul, E.; Mohamed, l.E. Antioxidant, Antibacterial and Cytotoxic Potential of Selected Macroalgae from the Red Sea, Sudan Coast. Int. Res. J. Biol. Sci. 2021, 10, 1–11. [Google Scholar]
- Hmani, I.; Ktari, L.; Ismail, A.; M’dallel, C.; El Bour, M. Assessment of the antioxidant and antibacterial properties of red algae (Rhodophyta) from the north coast of Tunisia. Euro-Mediterr. J. Environ. Integr. 2021, 6, 13. [Google Scholar] [CrossRef]
- Alkhalaf, M.I. Chemical composition, antioxidant, anti-inflammatory and cytotoxic effects of Chondrus crispus species of red algae collected from the Red Sea along the shores of Jeddah city. J. King Saud Univ. Sci. 2021, 33, 101210. [Google Scholar] [CrossRef]
- Murugesan, S.; Vinoth-Kumar, R.; Kotteswari, M.; Shanthi, N. In vitro antioxidant activity of marine red alga Gymnogongrus pygmaeus J. Agardh. Int. J. Pharm. Res. 2021, 13, 892–899. [Google Scholar] [CrossRef]
- Bocanegra, A.; Macho-Gonzalez, A.; Garcimartin, A.; Benedi, J.; Sanchez-Muniz, F.J. Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 3816. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Torrella, J.R.; Pages, T.; Viscor, G.; Torres, J.L. Edible Microalgae and Their Bioactive Compounds in the Prevention and Treatment of Metabolic Alterations. Nutrients 2021, 13, 563. [Google Scholar] [CrossRef]
- Priatni, S.; Budiwati, T.A.; Ratnaningrum, D.; Kosasih, w.; Andryani, R.; Susanti, H.; Susilaningsih, D. Antidiabetic screening of some Indonesian marine cyanobacteria collection. Bio Diversit AS 2016, 17, 642–646. [Google Scholar] [CrossRef]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef]
- Ahmed, B.E.; Badawi, M.H.; Mostafa, S.S.; Higazy, A.M. Human Anticancers and Antidiabetic Activities of the Cyanobacterium Fischerella sp. BS1-EG Isolated from River Nile, Egypt. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 3473–3485. [Google Scholar] [CrossRef]
- Kawee-Ai, A.; Kim, A.T.; Kim, S.M. Inhibitory activities of microalgal fucoxanthin against α-amylase, α-glucosidase, and glucose oxidase in 3T3-L1 cells linked to type 2 diabetes. J. Oceanol. Limnol. 2019, 37, 928–937. [Google Scholar] [CrossRef]
- Deepa, P.K.; Subramanian, A.; Manjusha, W.A. Phytochemical Screening and Evaluation of Antidiabetic Activity of the Marine Microalgae: Nannochloropsis sp. Int. J. Life Sci. Pharma Res. (IJLPR) 2020, 10, 36–40. [Google Scholar] [CrossRef]
- Priatni, S.; Ratnaningrum, D.; Kosasih, W. The Screening of Antidiabetic Activity and The Cultivation Study of Local Marine Microalgae. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1011, 012066. [Google Scholar] [CrossRef]
- Rashad, S.; El-Chaghaby, G.A. Marine Algae in Egypt distribution, phytochemical composition and biological uses as bioactive resources (a review). Egypt. J. Aquat. Biol. Fish. 2020, 24, 147–160. [Google Scholar] [CrossRef]
- Satpati, G.G.; Mal, N.; Pal, R. Seaweed-based interventions for diabetic complications: An analytical discourse. Syst. Biosci. Eng. 2021, 1, 53–66. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Alpha-amylase inhibition and antioxidant activity of marine green algae and its possible role in diabetes management. Pharmacogn. Mag. 2015, 11, S511–S515. [Google Scholar]
- Mohapatra, L.; Bhattamisra, S.; Panigrahy, R.; Parida, S. Evaluation of the antioxidant, hypoglycaemic and antidiabetic activities of some seaweed collected from the East Coast of India. J. Biomed. Pharmacol. 2016, 9, 365–375. [Google Scholar] [CrossRef]
- Pandithurai, M.; Murugesan, S.; Bhuvaneswari, S.; Thennarasan, S. In vitro α-amylase and α-glucosidase inhibition activity of methanolic extract of marine brown alga Spatoglossum asperum. Int. J. Adv. Pharm. 2015, 4, 83–87. [Google Scholar]
- Osman, N.A.H.K.; Siam, A.A.; El-Manawy, I.M.; Jeon, Y.-J. Anti-microbial and Anti-diabetic Activity of Six Seaweeds Collected from the Red Sea, Egypt. Int. J. Environ. Sci. 2019, 19, 55–60. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of Phlorotannins Extraction from Fucus vesiculosus and Evaluation of Their Potential to Prevent Metabolic Disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef]
- Tessema, H.A. Evaluation of the in vitro α-amylase enzyme inhibition potential of commercial dried laver (Porphyra species) seaweed protein hydrolysate. Turk. J. Fish. Aquat. Sci. 2018, 18, 547–556. [Google Scholar]
- Sanger, G.; Rarung, L.K.; Damongilala, L.J.; Kaseger, B.E.; Montolalu, L.A.D.Y. Phytochemical constituents and antidiabetic activity of edible marine red seaweed (Halymenia durvilae). IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 012069. [Google Scholar] [CrossRef]
- Ripa, S.A.; Aziz, F.B.; Islam, R.; Hasan, M.M.; Parvez, M.M.M.; Lipi, T.; Jubayar, M.; Roy, M.C. Antidiabetic effect of Spirulina (Spirulina platensis) in alloxan induced rabbit model. Int. J. Nat. Soc. Sci. 2018, 5, 48–53. [Google Scholar]
- Hussaini, S.; Hossain, M.I.; Islam, M.S.; Rafiq, K. Effects of Spirulina platensis on alloxan induced diabetic rats. Progress. Agric. 2018, 29, 139–146. [Google Scholar] [CrossRef]
- Nasirian, F.; Dadkhah, M.; Moradi-Kor, N.; Obeidavi, Z. Effects of Spirulina platensis microalgae on antioxidant and anti-inflammatory factors in diabetic rats. Diabetes Metab. Syndr. Obes. 2018, 11, 375–380. [Google Scholar] [CrossRef]
- Emami, S.; Olfati, A.O. Effects of dietary supplementing of Spirulina platensis and Chlorella vulgaris microalgae on hematologic parameters in streptozotocin-induced diabetic rats. Iran. J. Pediatr. Hematol. Oncol. 2017, 7, 163–170. [Google Scholar]
- Nasirian, F.; Sarir, H.; Moradi-Kor, N. Antihyperglycemic and antihyperlipidemic activities of Nannochloropsis oculata microalgae in Streptozotocin-induced diabetic rats. Biomol Concepts 2019, 10, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, L.; Bhattamisra, S.; Panigrahy, R.; Parida, S.; Pati, P. Antidiabetic effect of Sargassum wightii and Ulva fasciata in high fat diet and multi low dose streptozotocin induced type 2 diabetic mice. UK J. Pharm. Biosci. 2016, 4, 13–23. [Google Scholar] [CrossRef]
- Kumar, N.; Sharunetha, T. Ulva reticulata, a marine alga normalize streptozotocin induced lipid peroxidation in experimental diabetic rats. J. Drug Deliv. Ther. 2018, 8, 12–15. [Google Scholar]
- Labbaci, F.Z.; Boukortt, F.O. Beneficial Effects of Algerian Green Alga Ulva lactuca and Its Hydroethanolic Extract on Insulin Resistance and Cholesterol Reverse Transport in High-Fat/Streptozotocin Diabetic Rats. Prev. Nutr. Food Sci. 2020, 25, 353–361. [Google Scholar] [CrossRef]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic Potential of Marine Brown Algae-a Mini Review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef]
- Husni, A.; Pawestri, S.; Isnansetyo, A. Blood glucose level and lipid profile of alloxan–induced diabetic rats treated with Na-alginate from seaweed Turbinaria ornata (Turner) J.agardh. J. Teknol. (Sci. Eng.) 2016, 78, 7–14. [Google Scholar] [CrossRef]
- Kumar, P.S.; Sudha, S.; Prakash, S. Antidiabetic activity of aqueous extract of Padina boergesenii in streptozotocin-induced diabetic rats. Int. J. Pharm. Pharm. Sci. 2014, 6, 418–422. [Google Scholar]
- Kumar, R.V. Antidiabetic Potential of Marine Red Alga Champia Parvula (C. Agardh) by Inhibiting Key Metabolic Enzymes. World J. Pharm. Res. 2017, 6, 1466–1474. [Google Scholar] [CrossRef]
- Gabbia, D.; Carrara, M.; Martin, S.D. The brown alga Ascophyllum nodosum as a nutraceutical useful for the control of type II diabetes. Curr. Res. Diabetes Obes. J. 2018, 8, 57–60. [Google Scholar] [CrossRef]
- Gotama, T.L.; Husni, A. Antidiabetic Activity of Sargassum hystrix Extracts in Streptozotocin-Induced Diabetic Rats. Prev. Nutr. Food Sci. 2018, 23, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, S.; Gholampour, H.; Farzadinia, P.; Daneshi, A.; Ramavandi, B.; Moazzeni, A.; Keshavarz, M.; Bargahi, A. Anti-diabetic effects of Sargassum oligocystum on Streptozotocin-induced diabetic rat. Iran J. Basic Med. Sci. 2018, 21, 342–346. [Google Scholar] [CrossRef]
- Abdel-Karim, O.H.; Abo-Shady, A.M.; Ismail, G.A.; Gheda, S.F. Potential effect of Turbinaria decurrens acetone extract on the biochemical and histological parameters on alloxan-induced diabetic rats. Int. J. Environ. Health Res. 2021, 32, 1447–1468. [Google Scholar] [CrossRef]
- Murugesan, S.; Anand Babu, M.; Bhuvaneswari, S.; Kotteswari, M.; Thennarasan, S. In vitro antidiabetic activity of methanolic extracts of selected marine algae. Eur. J. Pharm. Med. Res. 2015, 2, 256–260. [Google Scholar]
- Radhika, D.; Priya, R. Assessment of antidiabetic activity of some selected seaweeds. Eur. J. Biomed. Pharm. Sci. 2015, 2, 151–154. [Google Scholar]
- Yang, T.H.; Yao, H.T.; Chiang, M.T. Red algae (Gelidium amansii) reduces adiposity via activation of lipolysis in rats with diabetes induced by streptozotocin-nicotinamide. J. Food Drug Anal. 2015, 23, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Rayapu, L.; Makkar, F.; Anandan, S.K.; Maneesh, A.; Chakraborty, K.; Valluru, L. Protective role of marine macroalgae extracts against STZ induced diabetic rats. J. Coast. Life Med. 2017, 5, 521–530. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Aly, H.F.; El-Sayed, A.B.; Amal, A.M. Role of Spirulina Platensis in the control of glycemia in DM2 rats. Int. J. Sci. Eng. Res. 2013, 4, 1731–1740. [Google Scholar]
- Nam, J.; Lee, W.; Yoon, I.; Kang, M.; Jang, H.; Youn, J.; Kim, B.; Kong, H.; Kim, K.-H.; Kim, Y.-h.; et al. Effect of a Brown Algae Extract on Postprandial Glucose Control in Neonatal Diabetic and Obese Rats. FASEB J. 2007, 21, A1085. [Google Scholar] [CrossRef]
- Selvaraj, S.; Palanisamy, S. Investigations on the anti-diabetic potential of novel marine seaweed Sargassum longiotom against alloxan-induced diabetes mellitus: A pilot study. Bangladesh J. Pharmacol. 2018, 9, 194–197. [Google Scholar] [CrossRef]
- Nagy, M.A. Biochemical and histopathological analysis of Hydroclathrus clathratus aqueous extract on alloxan induced diabetic rats. BioChem. Indian J. 2015, 9, 70–80. [Google Scholar]
- Chatterjea, M.N.; Shinde, R. Textbook of Medical Biochemistry; Jaypee Brothers Medical Publishers: New Delhi, India, 2002; Volume 58. [Google Scholar]
- Layam, A.; Reddy, C.L. Antidiabetic property of Spirulina. Diabetol. Croat. 2007, 35, 29–33. [Google Scholar]
- Pandey, J.; Tiwari, A.; Mishra, G.; Mishra, R. Role of Spirulina maxima in the control of blood glucose levels and body weight in streptozotocin induced diabetic male Wistar rats. J. Algal Biomass Util. 2011, 2, 35–37. [Google Scholar]
- Aissaoui, O.; Amiali, M.; Bouzid, N.; Belkacemi, K.; Bitam, A. Effect of Spirulina platensis ingestion on the abnormal biochemical and oxidative stress parameters in the pancreas and liver of alloxan-induced diabetic rats. Pharm. Biol. 2017, 55, 1304–1312. [Google Scholar] [CrossRef]
- Banu, A.T.; Mageswari, S. Nutritional status and effect of seaweed chocolate on anemic adolescent girls. Food Sci. Hum. Wellness 2015, 4, 28–34. [Google Scholar] [CrossRef]
- Yakubu, M.; Akanji, M.; Oladiji, A. Aphrodisiac potentials of the aqueous extract of Fadogia agrestis (Schweinf. Ex Hiern) stem in male albino rats. Asian J. Androl. 2005, 7, 399–404. [Google Scholar] [CrossRef]
- Dey, P.; Saha, M.; Chowdhuri, S.; Sen, A.; Sarkar, M.; Haldar, B.; Chaudhuri, T. Assessment of anti-diabetic activity of an ethnopharmacological plant Nerium oleander through alloxan induced diabetes in mice. J. Ethnopharmacol. 2015, 161, 128–137. [Google Scholar] [CrossRef]
- Kuriakose, G.; Kurup, M. Hepatoprotective effect of Spirulina lonar on paracetamol induced liver damage in rats. Asian J. Exp. Biol. Sci. 2010, 1, 614–623. [Google Scholar]
- Huang, X.J.; Choi, Y.K.; Im, H.S.; Yarimaga, O.; Yoon, E.; Kim, H.S. Aspartate Aminotransferase (AST/GOT) and Alanine Aminotransferase (ALT/GPT) Detection Techniques. Sensors 2006, 6, 756–782. [Google Scholar] [CrossRef]
- Mori, D.; Baviera, A.; de Oliveira Ramalho, L.; Vendramini, R.; Brunetti, I.; Pepato, M. Temporal response pattern of biochemical analytes in experimental diabetes. Biotechnol. Appl. Biochem. 2003, 38, 183–191. [Google Scholar] [CrossRef]
- Setorki, M.; Asgary, S.; Eidi, A.; Rohani, A.; Khazaei, M. Acute effects of vinegar intake on some biochemical risk factors of atherosclerosis in hypercholesterolemic rabbits. Lipids Health Dis. 2010, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Felig, P.; Marliss, E.; Ohman, J.; Cahill, C. Plasma amino acid levels in diabetic ketoacidosis. Diabetes 1970, 19, 727–729. [Google Scholar] [CrossRef]
- Asayama, K.; Nakane, T.; Uchida, N.; Hayashibe, H.; Dobashi, K.; Nakazawa, S. Serum antioxidant status in streptozotocin-induced diabetic rat. Horm. Metab. Res. Horm. Und Stoffwechs. Horm. Et Metab. 1994, 26, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Ohaeri, O. Effect of garlic oil on the levels of various enzymes in the serum and tissue of streptozotocin diabetic rats. Biosci. Rep. 2001, 21, 19–24. [Google Scholar] [CrossRef]
- Panigrahi, B.B.; Panda, P.K.; Patro, V.J. Comparative hepatoprotective activity of different extracts of spirulina against ccl 4 induced liver damage in rats. Int. J. Pharm. Sci. Rev. Res. 2010, 4, 200–202. [Google Scholar]
- Salem, S.I.; Ibrahim, A.K.; El-Olemy, K.A.a.M. Clinicopathological studies on the use of Spirulina platensis as a modern food supplement in alloxan-induced diabetic rats. Egyptain J. Comp. Pathol. Clin. Pathol. 2014, 27, 55–72. [Google Scholar]
- Abbas, A.S.; Shazly, M.o.; Ahmed, K.; Abdel-Mawla, E.; Ibrahi, A.K. Therapeutic effects of Spirulina platensis on streptozotocin-induced diabetic rats. Egypt. J. Comp. Path Clinic Path 2015, 28, 18–31. [Google Scholar]
- Brito, V.; da Rocha, J.; Puntel, G.; da Luz, S.; Barbosa, N.; de Carvalho, N.; Folmer, V. Inhibition of δ-aminolevulinate dehydratase is not closely related to the development of hyperglycemia in alloxan-induced diabetic mice. Exp. Toxicol. Pathol. Off. J. Ges. Fur Toxikol. Pathol. 2011, 63, 443–451. [Google Scholar] [CrossRef]
- Khan, M.; Shobha, J.; Mohan, I.; Rao Naidu, M.; Prayag, A.; Kutala, V. Spirulina attenuates cyclosporine-induced nephrotoxicity in rats. J. Appl. Toxicol. JAT 2006, 26, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Avdagić, N.; Cosović, E.; Nakas-Ićindić, E.; Mornjaković, Z.; Zaciragić, A.; Hadzović-Dzuvo, A. Spirulina platensis protects against renal injury in rats with gentamicin-induced acute tubular necrosis. Bosn. J. Basic Med. Sci. 2008, 8, 331. [Google Scholar] [CrossRef] [PubMed]
- Doumas, B.; Bayse, D.; Carter, R.; Peters, T.; Schaffer, R. A candidate reference method for determination of total protein in serum. I. Development and validation. Clin. Chem. 1981, 27, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Arise, R.O.; Ganiyu, A.I.; Oguntibeju, O. Lipid Profile, Antidiabetic and Antioxidant Activity of Acacia ataxacantha Bark Extract in Streptozotocin-Induced Diabetic Rats. In Antioxidant-Antidiabetic Agents and Human Health; Oguntibeju, O., Ed.; IntechOpen: London, UK, 2014. [Google Scholar]
- Alderson, N.; Chachich, M.; Frizzell, N.; Canning, P.; Metz, T.; Januszewski, A.; Youssef, N.; Stitt, A.; Baynes, J.; Thorpe, S. Effect of antioxidants and ACE inhibition on chemical modification of proteins and progression of nephropathy in the streptozotocin diabetic rat. Diabetologia 2004, 47, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, R.; John, S.A. Hypoglycemic activity of marine cyanobacteria in alloxan-induced diabetic rats. Pharmacologyonline 2008, 2, 704–714. [Google Scholar]
- Venkataraman, L.V. Spirulina: Global Reach of a Health Care product, 4th ed.; Souvenir: Noida, India, 1998; Volume 98, p. 175. [Google Scholar]
- Belay, A. The Potential Application of Spirulina (Arthrospira) as a Nutritional Health and Therapeutic Supplement in Health Management. J. Am. Nutraceutical Assoc. 2002, 5, 27–48. [Google Scholar]
- Parameshwar, S.; Srinivasan, K.K.; Rao, C.M. Oral antidiabetic activities of different extracts of Caesalpinia bonducella Seed Kernels. Pharm. Biol. 2002, 40, 590–595. [Google Scholar] [CrossRef]
- Soon, Y.; Tan, B. Evaluation of the hypoglycemic and anti-oxidant activities of Morinda officinalis in streptozotocin-induced diabetic rats. Singap. Med. J. 2002, 43, 77–85. [Google Scholar]
- Abdel-Raouf, N.; Al-Enazi, N.M.; Ibraheem, I.B.M.; Al-Harbie, R.M. Antibacterial and anti-hyperlipidemic activities of the brown alga Hormophysa cuneiformis from Ad Dammam Seashore. J. Appl. Pharm. Sci. 2015, 5, 114–125. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2010, 33, S62–S69. [Google Scholar] [CrossRef] [PubMed]
- Valado, A.; Pereira, M.; Amaral, M.; Cotas, J.; Pereira, L. Bioactivity of carrageenans in metabolic syndrome and cardiovascular diseases. Nutraceuticals 2022, 2, 441–454. [Google Scholar] [CrossRef]
- Shirwaikar, A.; Rajendran, K.; Dinesh Kumar, C.; Bodla, R. Antidiabetic activity of aqueous leaf extract of Annona squamosa in streptozotocin-nicotinamide type 2 diabetic rats. J. Ethnopharmacol. 2004, 91, 171–175. [Google Scholar] [CrossRef]
- Mir, S.H.; Baqui, A.; Baghat, R.C.; Darzi, M.M.; Shah, A.W. Biochemical and histomorphological study of streptozotocin-induced diabetes mellitus in rabbits. Pak. J. Nutr. 2008, 7, 359–364. [Google Scholar] [CrossRef]
- Ebuehi, O.A.T.; Ajuluchukwu, A.E.; Afolabi, O.T.; Akinwande, A.I. Oxidative stress in Alloxan--induced diabetes in female and male rats. Adv. Med. Dent. Sci. 2010, 3, 71–75. [Google Scholar]
- Cardoso, S.; Pereira, O.; Seca, A.; Pinto, D.; Silva, A. Seaweeds as preventive agents for cardiovascular diseases: From nutrients to functional foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhou, S.; Zhang, K.; Tang, J.; Guang, L.; Ying, Y.; Xu, Y.; Zhang, L.; Li, D. Chronic effects of berberine on blood, liver glucolipid metabolism and liver PPARs expression in diabetic hyperlipidemic rats. Biol. Pharm. Bull. 2008, 31, 1169–1176. [Google Scholar] [CrossRef]
- Aboonabi, A.; Rahmat, A.; Othman, F. Effect of pomegranate on histopathology of liver and kidney on generated oxidative stress diabetic induced rats. J. Cytol. Histol. 2015, 6, 1000294. [Google Scholar] [CrossRef]
- Nuhu, A.A. Spirulina (Arthrospira): An Important Source of Nutritional and Medicinal Compounds. J. Mar. Biol. 2013, 2013, 325636. [Google Scholar] [CrossRef]
- Abdel-Daim, M.; El-Bialy, B.; Rahman, H.; Radi, A.; Hefny, H.; Hassan, A. Antagonistic effects of Spirulina platensis against sub-acute deltamethrin toxicity in mice: Biochemical and histopathological studies. Biomed. Pharmacother. Biomed. Pharmacother. 2016, 77, 79–85. [Google Scholar] [CrossRef] [PubMed]
- El-Baky, H.H.A.; El-Baz, F.K.; El-Baroty, G.S. Production of phenolic compounds from Spirulina maxima microalgae and its protective effects in vitro toward hepatotoxicity model. Afr. J. Pharm. Pharmacol. 2009, 3, 133–139. [Google Scholar]
- Abdel-Daim, M.M. Pharmacodynamic interaction of Spirulina platensis with erythromycin in Egyptian Baladi bucks (Capra hircus). Small Rumin. Res. 2014, 120, 234–241. [Google Scholar] [CrossRef]
- Nugent, D.A.; Smith, D.M.; Jones, H.B. A Review of Islet of Langerhans Degeneration in Rodent Models of Type 2 Diabetes. Toxicol. Pathol. 2008, 36, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.; Farouk, S.; Madkour, F.; Azab, S. Anti-inflammatory and immunomodulatory effects of Spirulina platensis in comparison to Dunaliella salina in acetic acid-induced rat experimental colitis. Immunopharmacol. Immunotoxicol. 2015, 37, 126–139. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Shady, A.M.; Gheda, S.F.; Ismail, G.A.; Cotas, J.; Pereira, L.; Abdel-Karim, O.H. Antioxidant and Antidiabetic Activity of Algae. Life 2023, 13, 460. https://doi.org/10.3390/life13020460
Abo-Shady AM, Gheda SF, Ismail GA, Cotas J, Pereira L, Abdel-Karim OH. Antioxidant and Antidiabetic Activity of Algae. Life. 2023; 13(2):460. https://doi.org/10.3390/life13020460
Chicago/Turabian StyleAbo-Shady, Atef Mohamed, Saly Farouk Gheda, Gehan Ahmed Ismail, João Cotas, Leonel Pereira, and Omnia Hamdy Abdel-Karim. 2023. "Antioxidant and Antidiabetic Activity of Algae" Life 13, no. 2: 460. https://doi.org/10.3390/life13020460
APA StyleAbo-Shady, A. M., Gheda, S. F., Ismail, G. A., Cotas, J., Pereira, L., & Abdel-Karim, O. H. (2023). Antioxidant and Antidiabetic Activity of Algae. Life, 13(2), 460. https://doi.org/10.3390/life13020460