


Human Serum Mediated Bacteriophage Life Cycle Switch in Aggregatibacter actinomycetemcomitans Is Linked to Pyruvate Dehydrogenase Complex

Abstract
1. Introduction
2. Materials and Methods
2.1. Phage and Bacterial Strain Preparation
2.2. Growth Comparison after Mitomycin C Exposure
2.3. Transmission Electron Microscopy (TEM) Analysis
2.4. Bacteriophage Plaque Assays
2.5. Blue Native-Polyacrylamide Gel Electrophoresis (BN-PAGE)
2.6. Liquid Chromatography/Mass Spectrometry (LC/MS) Analysis
3. Results
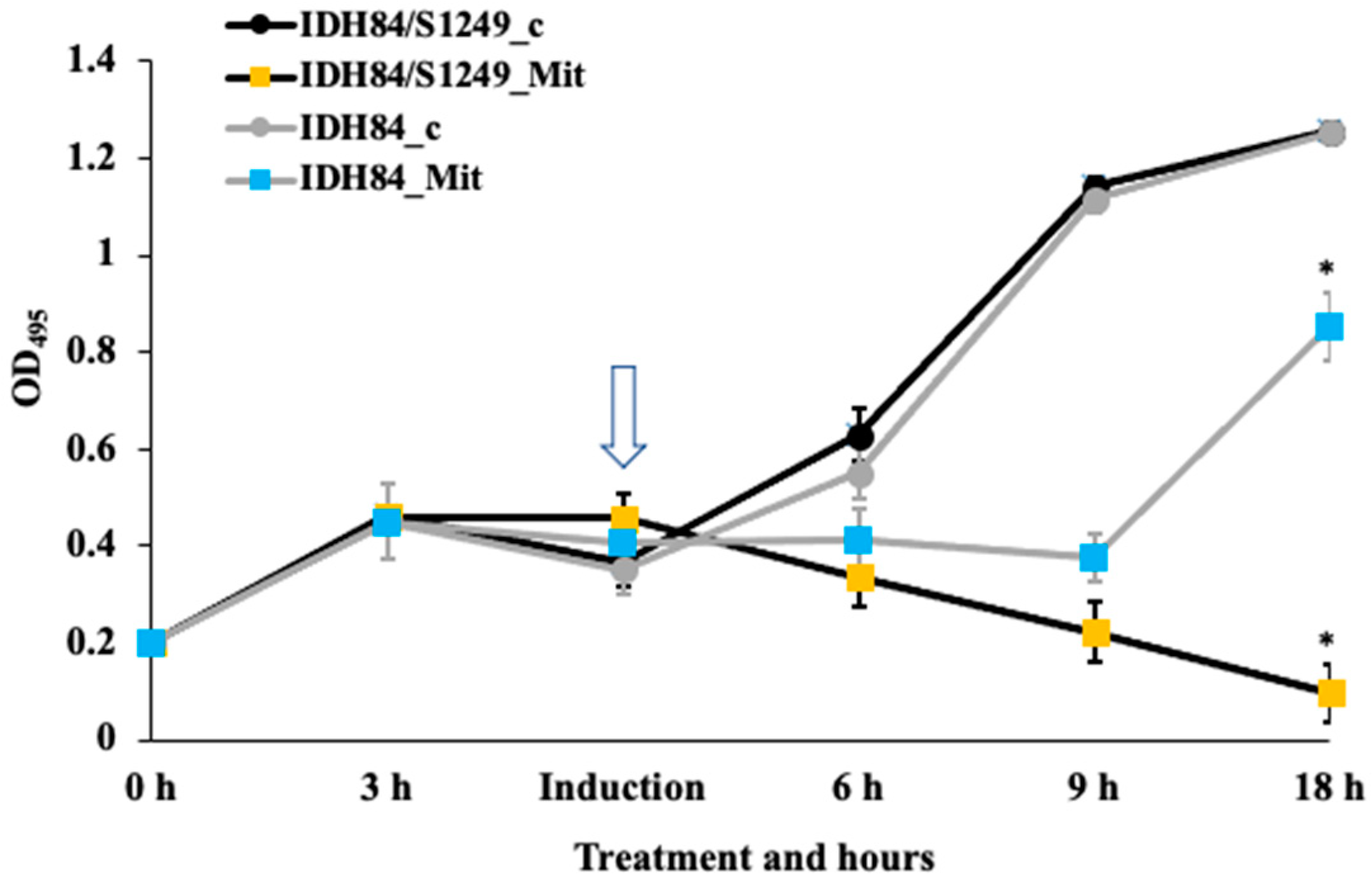
3.1. Differential Responses of the Lysogen and Its Isogenic, Phage-Naïve Strain to Mitomycin C
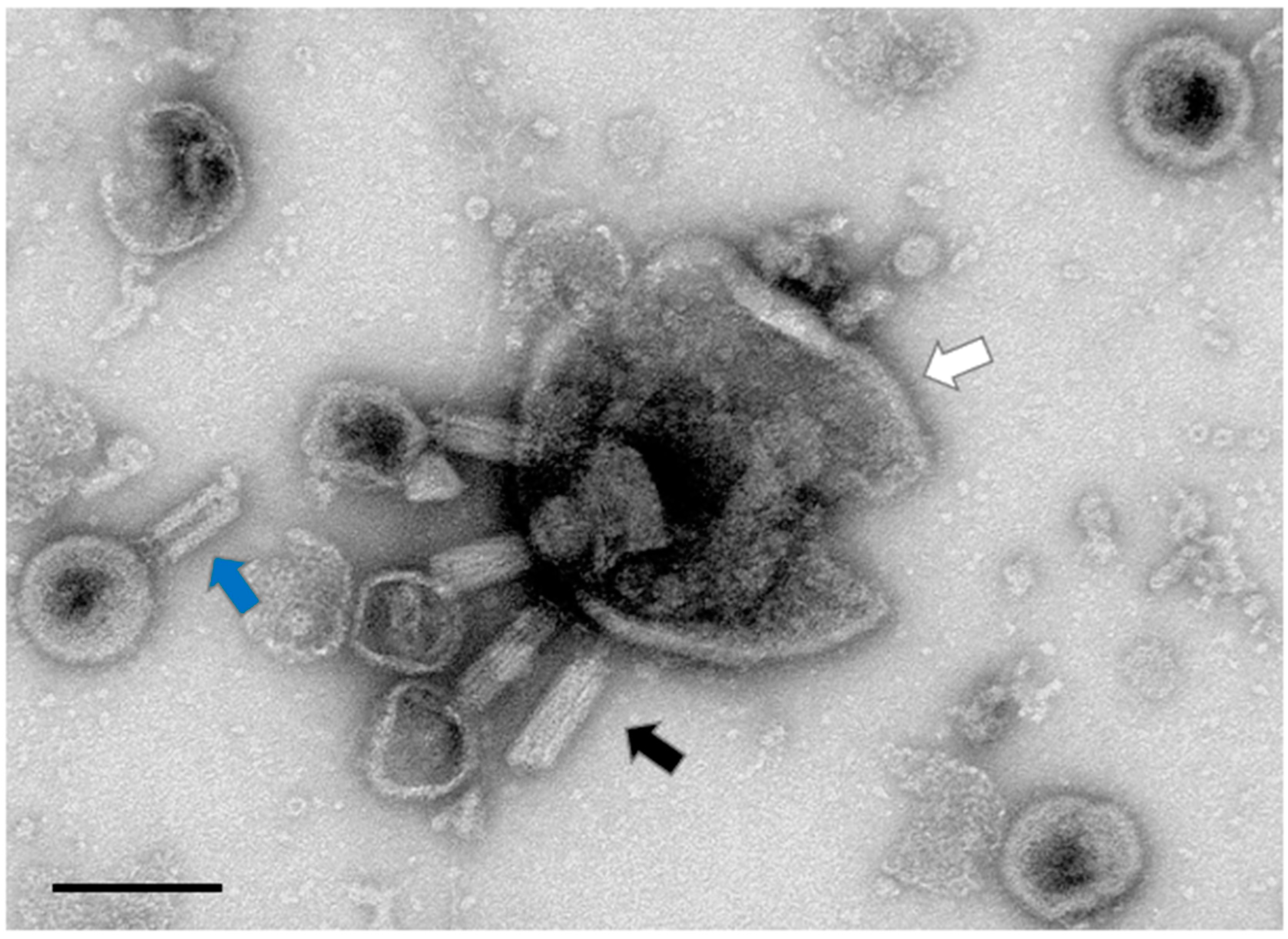
3.2. Aggregatibacter Phage S1249 in the Lytic State Demonstrated under TEM
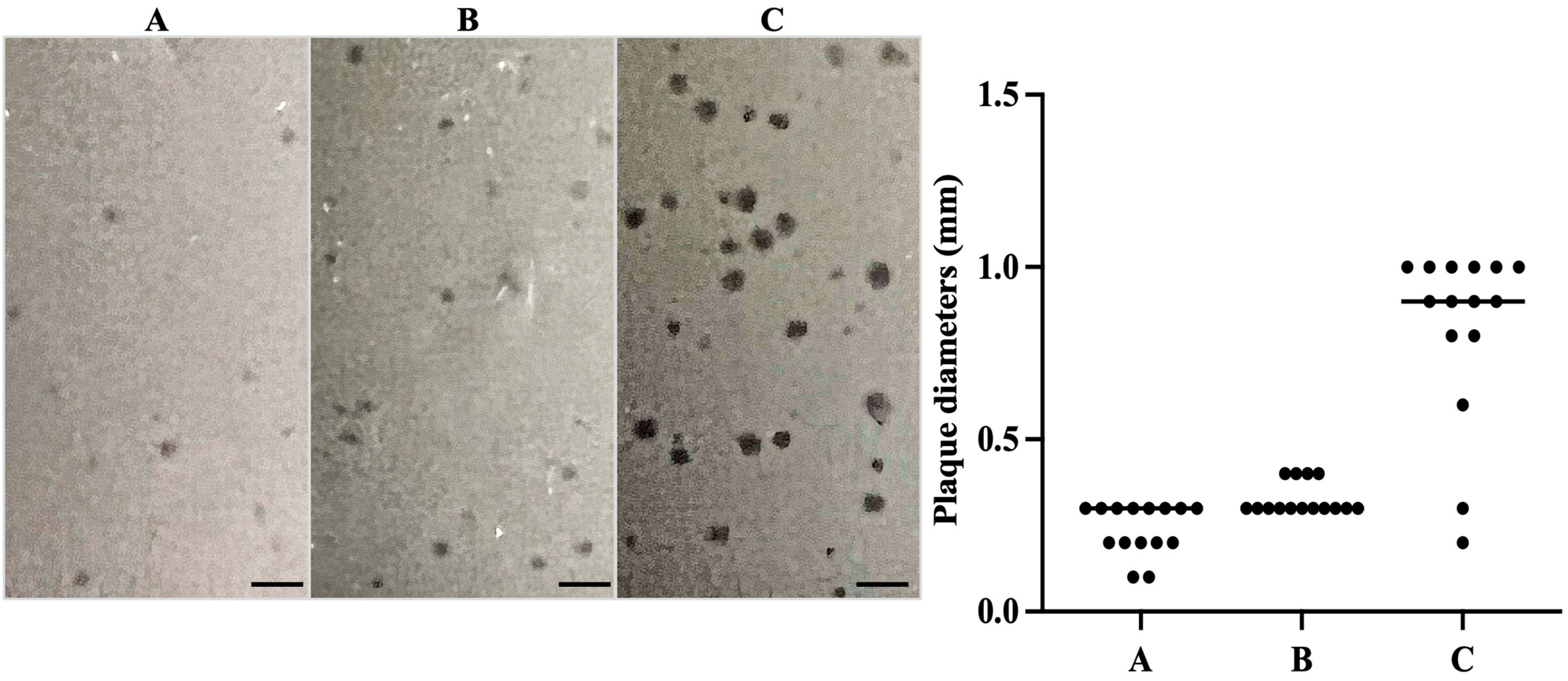
3.3. Plaque Assays Demonstrated Lytic Phage S1249 in the Presence of Human Serum
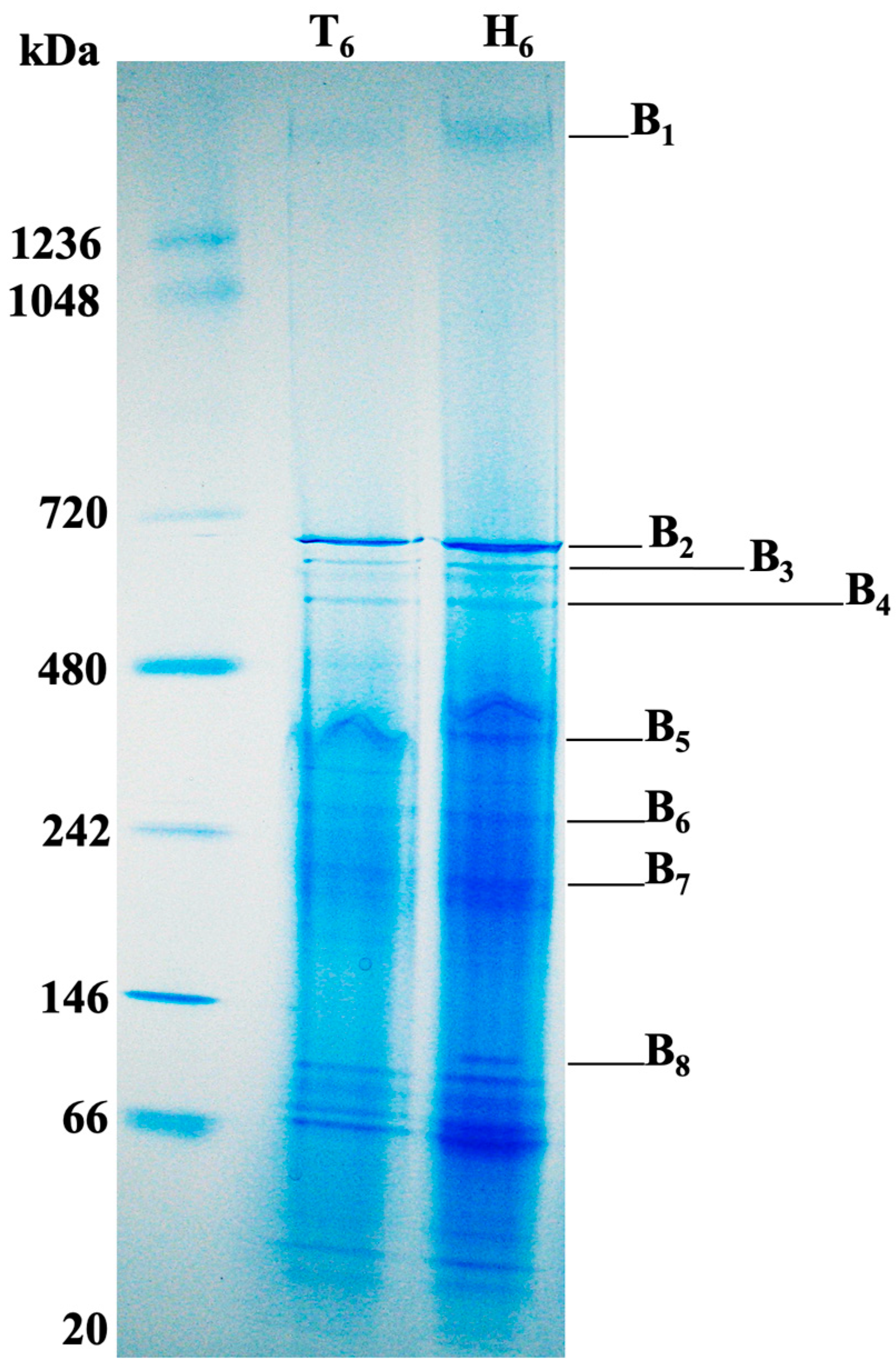
3.4. Membrane Associated Proteins of Aggregatibacter Phage S1249 Lysogen Grown in Human Serum
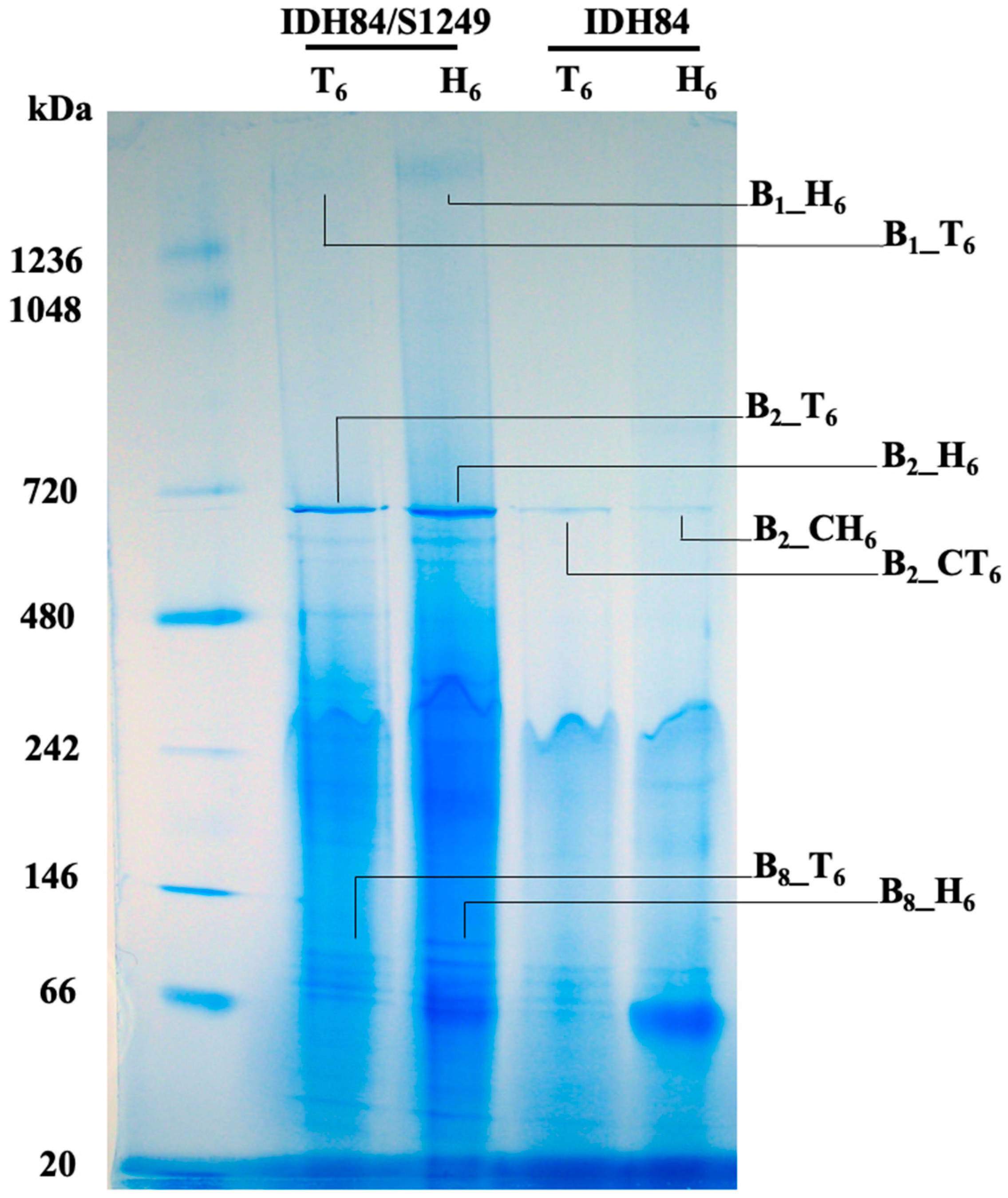
3.5. Bacterial Membrane Associated Protein Comparison between the Phage-Naïve Versus Its Isogenic Lysogenic Strain after Exposure to Human Serum
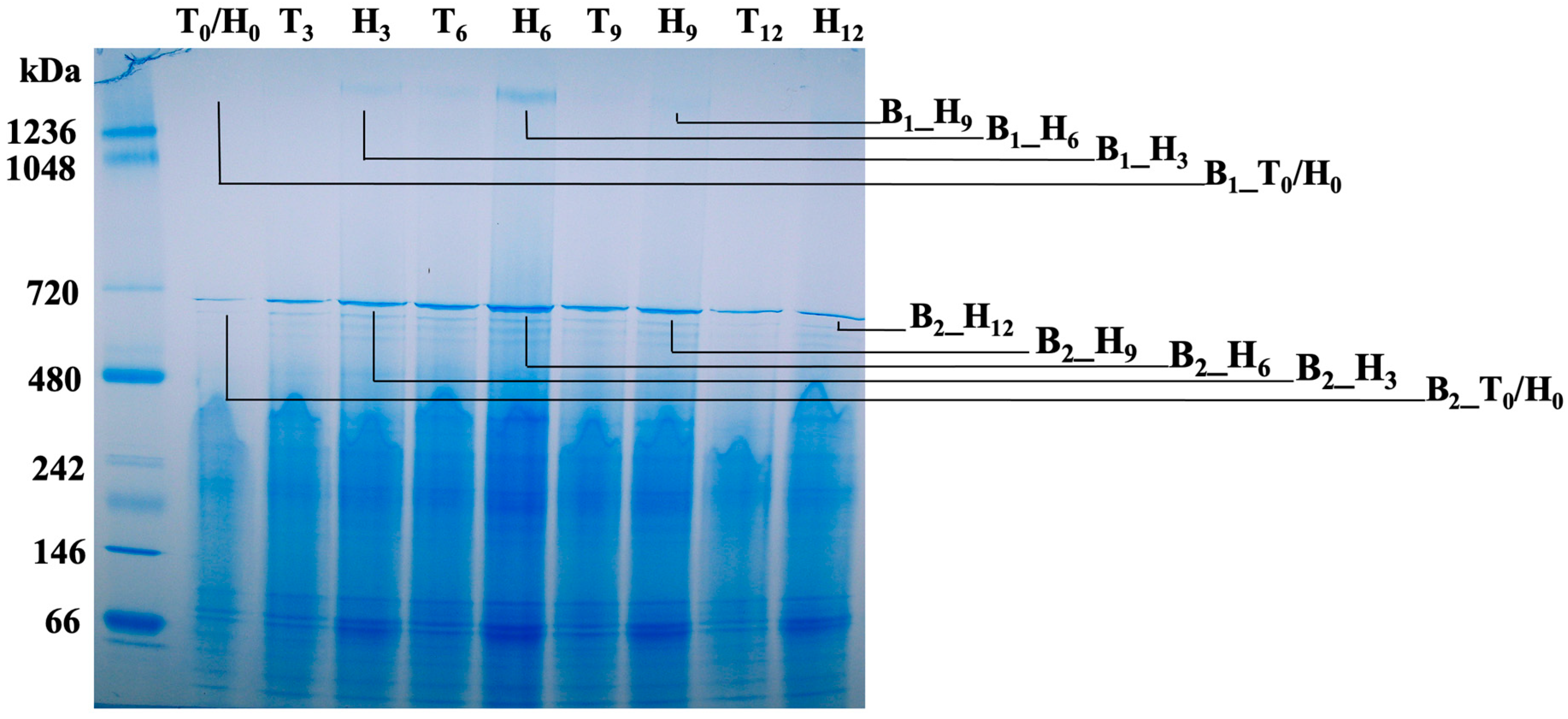
3.6. Membrane Protein Change Kinetics of the Lysogenic Bacterium Grown in Human Serum
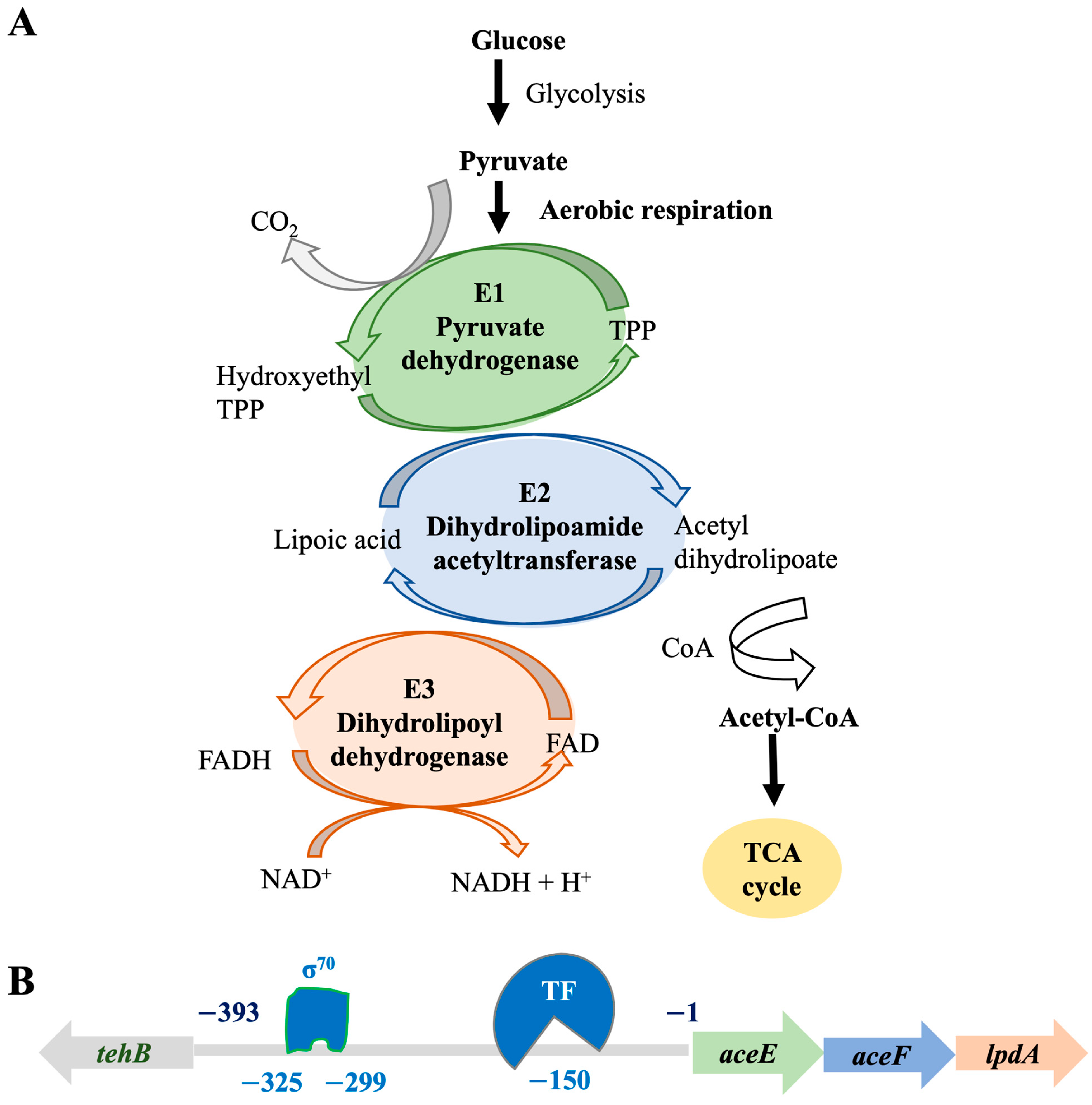
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaborators, A.R. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar]
- Caselli, E.; Fabbri, C.; D’Accolti, M.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the oral microbiome by whole-genome sequencing and resistome analysis: The complexity of the healthy picture. BMC Microbiol. 2020, 20, 120. [Google Scholar] [CrossRef] [PubMed]
- Ardila, C.M.; Bedoya-García, J.A. Antimicrobial resistance of Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis and Tannerella forsythia in periodontitis patients. J. Glob. Antimicrob. Resist. 2020, 22, 215–218. [Google Scholar] [CrossRef]
- Akrivopoulou, C.; Green, I.M.; Donos, N.; Nair, S.P.; Ready, D. Aggregatibacter actinomycetemcomitans serotype prevalence and antibiotic resistance in a UK population with periodontitis. J. Glob. Antimicrob. Resist. 2017, 10, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Roe, D.E.; Weinberg, A.; Roberts, M.C. Mobile rRNA methylase genes coding for erythromycin resistance in Actinobacillus actinomycetemcomitans. J. Antimicrob. Chemother. 1996, 37, 457–464. [Google Scholar] [CrossRef]
- Narayanan, A.M.; Ramsey, M.M.; Stacy, A.; Whiteley, M. Defining genetic fitness determinants and creating genomic resources for an oral pathogen. Appl. Environ. Microbiol. 2017, 83, e00797-17. [Google Scholar] [CrossRef] [PubMed]
- Crosby, J.A.; Kachlany, S.C. TdeA, a TolC-like protein required for toxin and drug export in Aggregatibacter (Actinobacillus) actinomycetemcomitans. Gene 2007, 388, 83–92. [Google Scholar] [CrossRef]
- Cortelli, J.R.; Aquino, D.R.; Cortelli, S.C.; Fernandes, C.B.; de Carvalho-Filho, J.; Franco, G.C.; Costa, F.O.; Kawai, T. Etiological analysis of initial colonization of periodontal pathogens in oral cavity. J. Clin. Microbiol. 2008, 46, 1322–1329. [Google Scholar] [CrossRef]
- Fine, D.H.; Patil, A.G.; Loos, B.G. Classification and diagnosis of aggressive periodontitis. J. Periodontol. 2018, 89 (Suppl. 1), S103–S119. [Google Scholar] [CrossRef]
- Chambers, S.T.; Murdoch, D.; Morris, A.; Holland, D.; Pappas, P.; Almela, M.; Fernandez-Hidalgo, N.; Almirante, B.; Bouza, E.; Forno, D.; et al. HACEK infective endocarditis: Characteristics and outcomes from a large, multi-national cohort. PLoS ONE 2013, 8, e63181. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef]
- Haase, E.; Zmuda, J.; Scannapieco, F. Identification and molecular analysis of rough-colony-specific outer membrane proteins of Actinobacillus actinomycetemcomitans. Infect Immun. 1999, 67, 2901–2908. [Google Scholar] [CrossRef]
- Tomich, M.; Planet, P.J.; Figurski, D.H. The tad locus: Postcards from the widespread colonization island. Nat. Rev. Microbiol. 2007, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Ruiz, T.; Barrantes-Reynolds, R.; Mintz, K.P. Molecular heterogeneity of EmaA, an oligomeric autotransporter adhesin of Aggregatibacter (Actinobacillus) actinomycetemcomitans. Microbiology 2007, 153, 2447–2457. [Google Scholar] [CrossRef]
- Tang, G.; Kitten, T.; Munro, C.L.; Wellman, G.C.; Mintz, K.P. EmaA, a potential virulence determinant of Aggregatibacter actinomycetemcomitans in infective endocarditis. Infect Immun. 2008, 76, 2316–2324. [Google Scholar] [CrossRef]
- Tang, G.; Mintz, K.P. Glycosylation of the collagen adhesin EmaA of Aggregatibacter actinomycetemcomitans is dependent upon the lipopolysaccharide biosynthetic pathway. J. Bacteriol. 2010, 192, 1395–1404. [Google Scholar] [CrossRef]
- Tang-Siegel, G.G.; Danforth, D.R.; Tristano, J.; Ruiz, T.; Mintz, K.P. The serotype a-EmaA adhesin of Aggregatibacter actinomycetemcomitans does not require O-PS synthesis for collagen binding activity. Microbiology 2022, 168, 001191. [Google Scholar] [CrossRef]
- Tang-Siegel, G.G.; Radermacher, M.; Mintz, K.P.; Ruiz, T. Serotype-specific sugars impact structure but not functions of the trimeric autotransporter adhesin EmaA of Aggregatibacter actinomycetemcomitans. J. Bacteriol. 2022, 204, e0021522. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.; Kaplan, J.; Furgang, D.; Mansfield, K.; Fine, D. A second Aggregatibacter actinomycetemcomitans autotransporter adhesin exhibits specificity for buccal epithelial cells in humans and old world primates. Infect Immun. 2007, 75, 4440–4448. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.E.; Meyer, D.H.; Fives-Taylor, P.M. Aae, an autotransporter involved in adhesion of Actinobacillus actinomycetemcomitans to epithelial cells. Infect Immun. 2003, 71, 2384–2393. [Google Scholar] [CrossRef]
- Lally, E.; Golub, E.; Kieba, I.; Taichman, N.; Rosenbloom, J.; Rosenbloom, J.; Gibson, C.; Demuth, D. Analysis of the Actinobacillus actinomycetemcomitans leukotoxin gene. Delineation of unique features and comparison to homologous toxins. J. Biol. Chem. 1989, 264, 15451–15456. [Google Scholar] [CrossRef]
- Tang, G.; Kawai, T.; Komatsuzawa, H.; Mintz, K. Lipopolysaccharides mediate leukotoxin secretion in Aggregatibacter actinomycetemcomitans. Mol. Oral Microbiol. 2012, 27, 70–82. [Google Scholar] [CrossRef] [PubMed]
- DiRienzo, J.M. Breaking the gingival epithelial barrier: Role of the Aggregatibacter actinomycetemcomitans cytolethal distending toxin in oral Infectious disease. Cells 2014, 3, 476–499. [Google Scholar] [CrossRef]
- Zambon, J.J.; Slots, J.; Genco, R.J. Serology of oral Actinobacillus actinomycetemcomitans and serotype distribution in human periodontal disease. Infect Immun. 1983, 41, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.; MacLean, L.; Gmur, R.; Wilson, M. Characterization of the O-polysaccharide structure of lipopolysaccharide from Actinobacillus actinomycetemcomitans serotype b. Infect Immun. 1996, 64, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.B.; MacLean, L.M.; Brisson, J.R.; Wilson, M.E. Structures of the antigenic O-polysaccharides of lipopolysaccharides produced by Actinobacillus actinomycetemcomitans serotypes a, c, d and e. Eur. J. Biochem. 1996, 242, 682–688. [Google Scholar] [CrossRef]
- Kaplan, J.; Perry, M.; MacLean, L.; Furgang, D.; Wilson, M.; Fine, D. Structural and genetic analyses of O polysaccharide from Actinobacillus actinomycetemcomitans serotype f. Infect Immun. 2001, 69, 5375–5384. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Saito, M.; Tsuzukibashi, O.; Kawashima, Y.; Ishida, S.; Hirasawa, M. Characterization of a new serotype g isolate of Aggregatibacter actinomycetemcomitans. Mol. Oral Microbiol. 2010, 25, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, G.; Johansson, E. Bactericidal effect of pooled human serum on Bacteroides melaninogenicus, Bacteroides asaccharolyticus and Actinobacillus actinomycetemcomitans. Scand J. Dent. Res. 1982, 90, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, R.; Komatsuzawa, H.; Kawai, T.; Yamada, S.; Goncalves, R.B.; Izumi, S.; Fujiwara, T.; Nakano, Y.; Suzuki, N.; Uchida, Y.; et al. Outer membrane protein 100, a versatile virulence factor of Actinobacillus actinomycetemcomitans. Mol. Microbiol. 2003, 50, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Permpanich, P.; Kowolik, M.J.; Galli, D.M. Resistance of fluorescent-labelled Actinobacillus actinomycetemcomitans strains to phagocytosis and killing by human neutrophils. Cell Microbiol. 2006, 8, 72–84. [Google Scholar] [CrossRef]
- Tang-Siegel, G.; Bumgarner, R.; Ruiz, T.; Kittichotirat, W.; Chen, W.; Chen, C. Human serum-specific activation of alternative sigma factors, the stress responders in Aggregatibacter actinomycetemcomitans. PLoS ONE 2016, 11, e0160018. [Google Scholar] [CrossRef]
- Tang-Siegel, G.G.; Chen, C.; Mintz, K.P. Phage induction by human serum in Aggregatibacter actinomycetemcomitans. J. Dent. Res. 2021, 100, Abstract 2048. Available online: https://iadr.abstractarchives.com/abstract/21iags-3567371/phage-induction-by-human-serum-in-aggregatibacter-actinomycetemcomitans (accessed on 22 December 2022).
- Tang-Siegel, G.G. Characteriz ation of Aggregatibacter phage S1249: Lysogenic and lytic phenotype switches. J. Dent. Res. 2022, 101, Abstract 0298. Available online: https://iadr.abstractarchives.com/abstract/51am-3652616/characterization-of-aggregatibacter-phage-s1249-lysogenic-and-lytic-phenotype-switches (accessed on 22 December 2022).
- Tang-Siegel, G.G.; Chen, C.; Mintz, K.P. Increased sensitivity of Aggregatibacter actinomycetemcomitans to human serum is mediated by induction of a bacteriophage. Mol. Oral Microbiol. 2022, 38, 1–13. [Google Scholar] [CrossRef]
- Suh, G.A.; Lodise, T.P.; Tamma, P.D.; Knisely, J.M.; Alexander, J.; Aslam, S.; Barton, K.D.; Bizzell, E.; Totten, K.M.C.; Campbell, J.L.; et al. Considerations for the use of phage therapy in clinical practice. Antimicrob. Agents Chemother. 2022, 66, e0207121. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Krukiewicz, K.; Yahya, G.; Haq, M.U.; Maryam, S.; Mosbah, R.A.; Saber, S.; Alrouji, M. The breadth of bacteriophages contributing to the development of the phage-based vaccines for COVID-19: An ideal platform to design the multiplex vaccine. Int. J. Mol. Sci. 2023, 24, 1536. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Nemeria, N.S.; Furey, W.; Jordan, F. The pyruvate dehydrogenase complexes: Structure-based function and regulation. J. Biol. Chem. 2014, 289, 16615–16623. [Google Scholar] [CrossRef]
- Garrett, R.H.; Grisham, C.M. Biochemistry, 6th ed.; Cengage Learning: Boston, MA, USA, 2016; pp. 643–677. [Google Scholar]
- Škerlová, J.; Berndtsson, J.; Nolte, H.; Ott, M.; Stenmark, P. Structure of the native pyruvate dehydrogenase complex reveals the mechanism of substrate insertion. Nat. Commun. 2021, 12, 5277. [Google Scholar] [CrossRef]
- White, D. (Ed.) Electron Transport, in The Physiology and Biochemistry of Prokarytes; Oxford University Press: New York, NY, USA, 2007; pp. 120–148. [Google Scholar]
- Chen, C.; Kittichotirat, W.; Si, Y.; Bumgarner, R. Genome sequence of Aggregatibacter actinomycetemcomitans serotype c strain D11S-1. J. Bacteriol. 2009, 191, 7378–7379. [Google Scholar] [CrossRef]
- Ellis, E.L.; Delbrück, M. The growth of bacteriophage. J. Gen. Physiol. 1939, 22, 365–384. [Google Scholar] [CrossRef]
- Jones, D.; Krueger, A.P. A rapid slide plaque technic for bacteriophage assay. J. Gen. Physiol. 1951, 34, 347–357. [Google Scholar] [CrossRef]
- Le, R.K.; Harris, B.J.; Iwuchukwu, I.J.; Bruce, B.D.; Cheng, X.; Qian, S.; Heller, W.T.; O’Neill, H.; Frymier, P.D. Analysis of the solution structure of Thermosynechococcus elongatus photosystem I in n-dodecyl-β-D-maltoside using small-angle neutron scattering and molecular dynamics simulation. Arch. Biochem. Biophys. 2014, 550–551, 50–57. [Google Scholar] [CrossRef]
- Lee, Y.C.; Bååth, J.A.; Bastle, R.M.; Bhattacharjee, S.; Cantoria, M.J.; Dornan, M.; Gamero-Estevez, E.; Ford, L.; Halova, L.; Kernan, J.; et al. Impact of detergents on membrane protein complex isolation. J. Proteome Res. 2018, 17, 348–358. [Google Scholar] [CrossRef]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Klaenhammer, T.R. Phage resistance in a phage-insensitive Strain of Streptococcus lactis: Temperature-dependent phage development and host-controlled phage replication. Appl. Environ. Microbiol. 1984, 47, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Loś, J.M.; Golec, P.; Wegrzyn, G.; Wegrzyn, A.; Loś, M. Simple method for plating Escherichia coli bacteriophages forming very small plaques or no plaques under standard conditions. Appl. Environ. Microbiol. 2008, 74, 5113–5120. [Google Scholar] [CrossRef]
- Stetsenko, A.; Guskov, A. An overview of the top ten detergents used for membrane protein crystallization. Crystals 2017, 7, 197. [Google Scholar] [CrossRef]
- Carey, J.N.; Mettert, E.L.; Fishman-Engel, D.R.; Roggiani, M.; Kiley, P.J.; Goulian, M. Phage integration alters the respiratory strategy of its host. Elife 2019, 8, e49081. [Google Scholar] [CrossRef]
- Genest, O.; Wickner, S.; Doyle, S.M. Hsp90 and Hsp70 chaperones: Collaborators in protein remodeling. J. Biol. Chem. 2019, 294, 2109–2120. [Google Scholar] [CrossRef]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. Recent advances in understanding catalysis of protein folding by molecular chaperones. FEBS Lett. 2020, 594, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ang, D.; Snavely, M.; Georgopoulos, C. Isolation and characterization of point mutations in the Escherichia coli grpE heat shock gene. J. Bacteriol. 1994, 176, 6965–6973. [Google Scholar] [CrossRef]
- Georgopoulos, C. Toothpicks, serendipity and the emergence of the Escherichia coli DnaK (Hsp70) and GroEL (Hsp60) chaperone machines. Genetics 2006, 174, 1699–1707. [Google Scholar] [CrossRef]
- Barriot, R.; Latour, J.; Castanié-Cornet, M.P.; Fichant, G.; Genevaux, P. J-domain proteins in bacteria and their viruses. J. Mol. Biol. 2020, 432, 3771–3789. [Google Scholar] [CrossRef]
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 2008, 283, 18473–18477. [Google Scholar] [CrossRef] [PubMed]
- Garcie, C.; Tronnet, S.; Garénaux, A.; McCarthy, A.J.; Brachmann, A.O.; Pénary, M.; Houle, S.; Nougayrède, J.P.; Piel, J.; Taylor, P.W.; et al. The bacterial stress-responsive Hsp90 chaperone (HtpG) Is required for the production of the genotoxin colibactin and the siderophore Yersiniabactin in Escherichia coli. J. Infect Dis. 2016, 214, 916–924. [Google Scholar] [CrossRef]
- Hu, J.; Toft, D.O.; Seeger, C. Hepadnavirus assembly and reverse transcription require a multi-component chaperone complex which is incorporated into nucleocapsids. Embo J. 1997, 16, 59–68. [Google Scholar] [CrossRef]
- Taguwa, S.; Kambara, H.; Omori, H.; Tani, H.; Abe, T.; Mori, Y.; Suzuki, T.; Yoshimori, T.; Moriishi, K.; Matsuura, Y. Cochaperone activity of human butyrate-induced transcript 1 facilitates hepatitis C virus replication through an Hsp90-dependent pathway. J. Virol. 2009, 83, 10427–10436. [Google Scholar] [CrossRef] [PubMed]
- Burnett, B.J.; Altman, R.B.; Ferguson, A.; Wasserman, M.R.; Zhou, Z.; Blanchard, S.C. Direct evidence of an elongation factor-Tu/Ts·GTP·Aminoacyl-tRNA quaternary complex. J. Biol. Chem. 2014, 289, 23917–23927. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Bands * | Top Five Most Abundant Proteins Based on Total Spectrum Counts (PSMs **) | |
|---|---|---|
| A. actinomycetemcomitans | Bacteriophage S1249 *** | |
| B1 | Molecular chaperone GroEL (Hsp60, 463) E1 (252) **** E3 (165) E2 (149) Pilus assembly protein TadG (42) | D11S_2223 (1) |
| B2 | GroEL (Hsp60, 1651) DNA-directed RNA polymerase subunit β (RpoB, D11S_0063, 125) DNA-directed RNA polymerase (DdRP) subunit β (D11S_0064, 107) α-2-marcoglobulin (95) Acetaldehyde dehydrogenase (85) | D11S_2237 (4) D11S_2262 (1) D11S_2208 (1) D11S_2261 (1) |
| B3 | Malic enzyme (291) Citrate lysate (249) DdRP subunit β (D11S_0064, 159) RpoB (D11S_0063, 145) ATP synthase F0-F1 β (91) | D11S_2237 (6) |
| B4 | ATP synthase F0-F1 β (260) ATP synthase F0-F1 α (195) DdRP subunit β (D11S_0064, 183) RpoB (D11S_0063, 168) Malic enzyme (94) | D11S_2237 (8) D11S_2240 Endolysin protein (2) D11S_2239 Lytic protein Rz (2) |
| B5 | Fumarate hydratase (331) Pyruvate formate-lyase (PFL, 179) Na(+)-NQR subunit A (153) Alanyl-tRNA synthetase (133) Pyruvate kinase (120) | D11S_2267 (3) Endolysin protein (2) Lytic protein Rz (2) D11S_2242 (2) |
| B6 | PFL (240) Aspartate ammonia-lyase (170) Glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 129) DnaK (Hsp70, 115) Aspartyl-tRNA synthetase (102) | D11S_2223 (5) D11S_2267 (3) |
| B7 | PFL (553) Fumarate reductase (Frd, 262) GAPDH (205) Purine nucleoside phosphorylase (205) Preprotein translocase, SecA (178) | D11S_2267 (3) D11S_2260 (3) |
| B8 | Elongation factor Ts (EF-Ts, 235) Phosphoenolpyruvate carboxykinase (199) Elongation factor Tu (EF-Tu, 187) DnaK (Hsp70, 148) Heat shock protein 90 (Hsp90, 122) | D11S_2258 (12) D11S_2267 (12) D11S_2257 (7) D11S_2228 (2) |
| Protein Bands * | Top Five Identified Proteins of A. actinomycetemcomitans (PSMs **) |
|---|---|
| B1_T6 | GroEL (34) *** E1 (18) E3 (18) E2 (15) Porin OmpA (4) |
| B1_H6 | GroEL (463) E1 (252) E3 (165) E2 (149) TadG (42) |
| B2_CT6 | GroEL (246) TadG (72) α-2-marcoglobulin (58) EF-Tu (53) EF-Ts (50) |
| B2_CH6 | GroEL (230) TadG (70) Transaldolase (31) Malic enzyme (31) Sugar ABC transporter substrate binding protein (31) |
| B2_T6 | GroEL (1193) α-2-marcoglobulin (145) RpoB (D11S_0063, 122) DdRP subunit β (D11S_0064, 90) Malic enzyme (77) |
| B2_H6 | GroEL (1651) RpoB (D11S_0063, 125) DdRP subunit β (D11S_0064, 107) α-2-marcoglobulin (95) Acetaldehyde dehydrogenase (85) |
| B8_T6 | Phosphoenolpyruvate carboxykinase (PEPCK, 167) EF-Ts (131) Heat shock protein 90 (127) EF-Tu (116) Meth_synth # (115) |
| B8_H6 | EF-Ts (275) EF-Tu (226) PEPCK (195) Heat shock protein 90 (131) DnaK (122) |
| Protein Bands * | Top Five Identified Proteins (PSMs **) | |
|---|---|---|
| A. actinomycetemcomitans | Attached Human Serum Proteins | |
| B1_T0/H0 | GroEL (18) E1 (3) E3 (3) E2 (3) Hemolysin (3) | N/A |
| B1_H3 | E1 (94) GroEL (86) E3 (67) E2 (44) Hemolysin (14) | Keratin, type II cytoskeletal 1 (42) Keratin, type I cytoskeletal 9 (28) Keratin, type I cytoskeletal 10 (24) Keratin, type II cytoskeletal 2 epidermal (22) IGHG1 (22) |
| B1_H6 | GroEL (463) E1 (252) E3 (165) E2 (149) TadG (42) | Apolipoprotein B-100 (61) *** Apolipoprotein A-I (44) IGHG1 (38) IGHM (30) Keratin, type I cytoskeletal 9 (29) |
| B1_H9 | GroEL (102) E1 (71) E3 (42) E2 (39) Porin (23) | Apolipoprotein A-I (40) Keratin, type II cytoskeletal 2 epidermal (36) Apolipoprotein A-IV (32) Keratin, type II cytoskeletal 1 (30) Keratin, type I cytoskeletal 10 (29) |
| Protein Bands * | Top Five Identified Proteins (PSMs **) | ||
|---|---|---|---|
| A. actinomycetemcomitans1 | Attached Human Serum Proteins 2 | Phage S1249 *** | |
| B2_T0/H0 | Meth_synth (135) Elongation factor Tu (78) PEPCK # (68) Elongation factor Ts (65) Pilus assembly protein CpaF (43) | N/A | N/D |
| B2_H3 | GroEL (495) α-2-marcoglobulin (79) Citrate lyase subunit alpha (41) DdRP subunit β (D11S_0064, 39) Malic enzyme (39) | Keratin, type II cytoskeletal 1 (52) Haptoglobin (31) Keratin, type I cytoskeletal 10 (28) IGHG1 (25) Keratin, type I cytoskeletal 9 (24) | N/D |
| B2_H6 | GroEL (1651) **** RpoB (D11S_0063, 125) DdRP subunit β (D11S_0064, 107) α-2-marcoglobulin (95) Acetaldehyde dehydrogenase (85) | Complement C3 (29) IGHG1 (28) Serum albumin (21) Keratin, type II cytoskeletal 1 (21) Haptoglobin (20) | D11S_2237 (4) D11S_2262 (1) D11S_2208 (1) D11S_2261 (1) |
| B2_H9 | GroEL (502) α-2-marcoglobulin (97) TadG (49) Metallophosphatase (43) Malic enzyme (41) | Complement C3 (59) Keratin, type II cytoskeletal 1 (39) IGHG1 (37) Keratin, type I cytoskeletal 9 (29) Haptoglobin (26) | N/D |
| B2_H12 | GroEL (558) α-2-marcoglobulin (63) TadG (42) Acetaldehyde dehydrogenase (40) Malic enzyme (39) | Complement C3 (75) Keratin, type II cytoskeletal 1 (75) Keratin, type I cytoskeletal 9 (49) IGHG1 (49) Keratin, type II cytoskeletal 6C (42) | N/D |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang-Siegel, G.G. Human Serum Mediated Bacteriophage Life Cycle Switch in Aggregatibacter actinomycetemcomitans Is Linked to Pyruvate Dehydrogenase Complex. Life 2023, 13, 436. https://doi.org/10.3390/life13020436
Tang-Siegel GG. Human Serum Mediated Bacteriophage Life Cycle Switch in Aggregatibacter actinomycetemcomitans Is Linked to Pyruvate Dehydrogenase Complex. Life. 2023; 13(2):436. https://doi.org/10.3390/life13020436
Chicago/Turabian StyleTang-Siegel, Gaoyan Grace. 2023. "Human Serum Mediated Bacteriophage Life Cycle Switch in Aggregatibacter actinomycetemcomitans Is Linked to Pyruvate Dehydrogenase Complex" Life 13, no. 2: 436. https://doi.org/10.3390/life13020436
APA StyleTang-Siegel, G. G. (2023). Human Serum Mediated Bacteriophage Life Cycle Switch in Aggregatibacter actinomycetemcomitans Is Linked to Pyruvate Dehydrogenase Complex. Life, 13(2), 436. https://doi.org/10.3390/life13020436