Abstract
The recent pandemic of COVID-19 caused by the SARS-CoV-2 virus has brought upon the world an unprecedented challenge. During its acute dissemination, a rush for vaccines started, making the scientific community come together and contribute to the development of efficient therapeutic agents and vaccines. Natural products have been used as sources of individual molecules and extracts capable of inhibiting/neutralizing several microorganisms, including viruses. Natural extracts have shown effective results against the coronavirus family, when first tested in the outbreak of SARS-CoV-1, back in 2002. In this review, the relationship between natural extracts and SARS-CoV is discussed, while also providing insight into misinformation regarding the use of plants as possible therapeutic agents. Studies with plant extracts on coronaviruses are presented, as well as the main inhibition assays and trends for the future regarding the yet unknown long-lasting effects post-infection with SARS-CoV-2.
2. Morphology and Infection Route of Coronavirus
Coronaviruses belong to the Coronaviridae family, consisting of four genera, namely, alpha, beta, gama and delta coronavirus; alpha and beta genera being the ones that infect. Until 2019, SARS-CoV and MERS-CoV were the most highly contagious coronaviruses, while the other four were responsible for the common colds (HCoV-OC43, HCoV-229E, HCoV-NL63 and HCoV-HKU1) [1,8].
Morphologically, coronaviruses display three protein structures on their surface: the spike glycoprotein, the viral membrane glycoprotein, and the envelope protein (Figure 1). The spike protein mediates the entrance into the cell through the host’s angiotensin-converting enzyme 2 (ACE2) (Figure 2) surface receptor; the membrane glycoprotein is unique compared to other glycoproteins in its features making it responsible for its intracellular budding. The envelope protein is involved in the virus assembly, budding and pathogenesis [9,10]. The spikes present on the virus’ surface make it resemble a crown, ergo, the name corona (crown in Spanish). Its genome ranges between 26 and 32 kb, with 6–11 open reading frames that encode 9680 amino acid polyproteins.
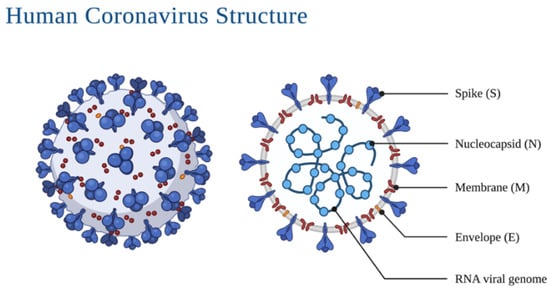
Figure 1.
Detail of the SARS-CoV-2 virus and different aspects of its morphology.
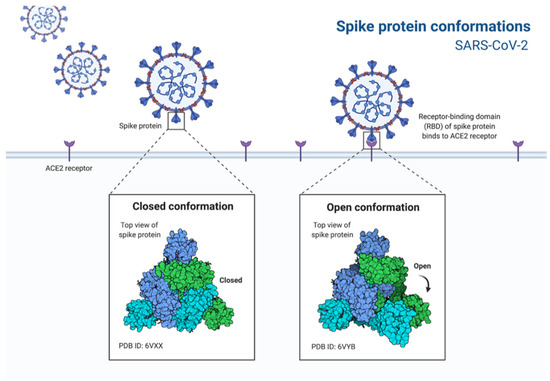
Figure 2.
Depiction of the conformation change in the spike protein of SARS-CoV-2 to bind to the cell ACE2 receptors.
Specific genes in the open reading frame (ORF1) downstream regions, which encode proteins for viral replication, nucleocapsid and spikes formation, are present in all coronaviruses. The RNA is uncoated to allow translation of the two polyproteins, transcription of the subgenomic RNAs and replication of the viral genome. Formed envelope glycoproteins are inserted in the Golgi membranes; genomic RNA and nucleocapsid proteins combine to form nucleocapsids, and the viral particles bud into the ER–Golgi intermediate compartment. Virion-containing vesicles subsequently fuse with the plasma membrane to release the virus [11].
Downregulation of ACE2 and the binding of the spike protein of this receptor contribute to lung injury during SARS, resulting in the excessive production of angiotensin II, stimulating the type 1a angiotensin II receptor (AGRT1A), increasing the pulmonary vascular permeability and explaining the increased lung pathology, with decreased expression of ACE2 [11].
Coronavirus entry into host cells is dependent on the binding of the spike glycoprotein to the cellular receptor. SARS-CoV and SARS-CoV2 use the host’s ACE2 protein, which usually breaks down angiotensinogen and regulates blood pressure and inflammation, as a doorway to enter the cell by binding to it and sneaking into the host cell, many by fusion and endosomal pathway entry. Due to a high number of ACE2 receptors in the lungs, the coronavirus mainly and initially infects the lung tissue, but can spread to several organs due to most tissues being lined with ACE2. After the entrance, viral RNA is released into the host’s cytoplasm and undergoes translation, generating both genomic and subgenomic RNA. The virion assembly occurs in the endoplasmic reticulum and the Golgi complex and is then released through vesicles [9,12].
3. Antiviral Therapies and Vaccine Strategies
When the COVID-19 pandemic reached global proportions, considerable private and public funds were poured into the development of vaccines and antiviral drugs. While vaccines were still at least a year away from being produced on a large scale, which only occurred at the end of 2020 many other drugs were proposed (Table 1). As the pandemic rose in severity during the year of 2021 with new variants, so did the quest to find suitable existing drugs and new ones to fight back in severe disease cases.
In September 2021, the European Medicines Agency (EMA) has only one authorized drug against COVID-19, remdesivir, sold under the brand name of Veklury®, while three marketing authorization applications had been submitted, namely, Olumiant® (baricitinib), Kineret® (anakinra) and RoActemra® (tocilizumab), all of which are already authorized for other illnesses within the EU. The EMA also stated that four medicines are under rolling review, namely, Bamlanivimab and Etesevimab, Regdanvimab, REGN-COV2® (casirivimab/imdevimab) and Sotrovimab (December 2021). The Food and Drug Administration of the United States of America (FDA) states on their webpage the following “Drug and Biological Therapeutic Products” with emergency approval: Actemra® (tocilizumab), Sotrovimab®, Bamlanivimab and Etesevimab®, REGEN-COV® (Casirivimab and Imdevimab), Olumiant® (barcitinib), COVID-19 convalescent plasma and remdesivir. Other drugs have been proposed to treat COVID-19 but have not been effective in showing any improvements in patients, namely, hydroxychloroquine, lopanivir/ritonavir and convalescent plasma [13,14,15].

Table 1.
Approved, in analysis and permission-sought drugs in the EU and US against COVID-19 (September 2021).
Table 1.
Approved, in analysis and permission-sought drugs in the EU and US against COVID-19 (September 2021).
| Rug Name | Main Function | Permission Sought | Reference |
|---|---|---|---|
| Veklury® (remdesivir) | After conversion to remdesivir monophosphate, this compound stalls viral RNA-polymerase | EU and US | [16] |
| Olumiant® (baricitinib) | This tyrosine kinase regulates the immune response of the body, avoiding a cytokine storm | EU and US | [17] |
| Kineret® (anakinra) | This recombinant interleukin (IL)-1 receptor antagonist helps control the hyperinflammatory syndrome of COVID-19 | EU | [18] |
| RoActemra® Actemra® (tocilizumab) | Recombinant humanized monoclonal antibody that prevents the binding of IL-6 to its receptor, inhibiting the inflammatory cascade | EU and FDA | [19] |
| Bamlanivimab and etesevimab | Neutralizing monoclonal antibodies that target the spike glycoprotein of the virus rendering it impossible to enter the host cell | EU and US | [20] |
| Regdanvimab | Monoclonal antibodies which also reduce viral entry in host cells | EU | [21] |
| REGN-COV-2® Casirivimab/imdevimab | Monoclonal antibodies that also target the spike glycoprotein | US | [22] |
| Sotrovimab® (VIR-7831) | Monoclonal antibodies that reduce viral entry into host cells and help clear infected cells | EU and US | [23] |
The main therapies against COVID-19 are antiviral drugs and monoclonal antibodies, which have shown some improvements in moderate and severe diseases. Still, some resistance has been found to some drugs, namely, the monoclonal antibodies Bamlanivimab and etesevimab, derived from mutations in the spike glycoprotein that changes its conformation, rendering the antibodies less effective [24].
However, the most effective method of controlling the pandemic has been proven to be through vaccines (Table 2).
Table 2.
Approved and under rolling review of vaccines in the EU and the US.
Among the existing vaccines, there are four different technologies. One of the most effective is the mRNA vaccines, which use a strain of RNA from the coronavirus, which instructs the human cells how to build the spike protein that is then recognized by the immune system, creating antibodies to fight it [25]. This technology is used by the approved Comirnaty and Spikevax vaccines, and the CureVac, which is still under rolling review. Another technology is the vector vaccine, used by the approved Vaxzevria and Janssen, in which a portion of the genetic material of the coronavirus is placed in a different virus (viral vector), usually an adenovirus, which when inside human cells releases the viral load and instructs the cells to make copies of the spike glycoprotein. This technology is also used by the Sputnik V vaccine [26]. The third technology used in the approved and rolling review vaccines is the protein subunit technology which includes only parts of the coronavirus’ RNA, namely, the spike glycoprotein, which when inside the cells makes the host’s immune system recognize these proteins and create antibodies. Novavax and Sanofi use this technology in their vaccines. Finally, Sinopharm uses the classical approach in terms of vaccines, opting to inject the whole SARS-CoV-2 virus in an inactivated form, allowing the immune system to build up defenses against this virus [26].
4. Plant Metabolites: Debunking Misconceptions
The COVID-19 pandemic showed that even in the face of worldwide unrest, millions of infections and deaths, certain denier movements and organizations were still questioning the pandemic, its origins, and, in some cases, proposing homemade remedies and questionable medications to fight the virus or infection symptoms. These groups were fueled by misinformation mainly spread through the Internet, specifically social media [27]. Among the proposed remedies, many of them based on empirical knowledge, using simple logic, it was assumed that if the symptoms of COVID-19 were those of the common flu then the same homemade remedies could help and cure the infection. These remedies and Chinese medicine-based papers have been published [28,29,30,31,32,33,34] alongside others that claim that specific vitamins and other remedies may improve the recovery or boost the immune system against SARS-CoV-2, but not without controversy [35,36,37,38]. One example of these misconceptions was the use of chloroquine and hydroxychloroquine (antimalarial drugs) against covid, which after much research was deemed to not have any positive effect on overall mortality, ventilation needs and hospitalization, despite promising results in vitro. Thus, it remains to be confirmed if it has any effect on the early stages of the disease [39,40], and one report event accounts for a worsening of the clinical status [41]. Another proposed drug that was proposed as a treatment for COVID-19 was ivermectin, an antiparasitic treatment. Several publications have also debunked its use, claiming that the drug did not result in a lower incidence of admission to hospitals and prolonged emergency observation [42]. Furthermore, in another clinical trial, neither ivermectin or metformin and fluvoxamine showed effects on hypoxemia, hospitalization or death of patients with COVID-19 [43].
While there is generalized scientific support that plants and their metabolites may play a role in drugs against COVID, its prophylaxis, or be used as active agents in fomites or textiles, the approach to using natural molecules should be the same as any other molecule. A big misconception present in most societies is that natural molecules are always safer, more effective, and sustainable, although this is not always the case [44].
Using the SCOPUS database and VOSviewer software, a brief relation between plants and SARS-CoV-2 can be established. About 1504 documents show up using the keywords “coronavirus”, “covid”, “sars”, “plant” and “extract” from 2020 to 2022 within the SCOPUS search engine. Documents that contain at least one of the searched keywords were clustered by VOSviewer according to their relevance and relationship. The plot of the relationships between the documents with these keywords is shown in Figure 3.
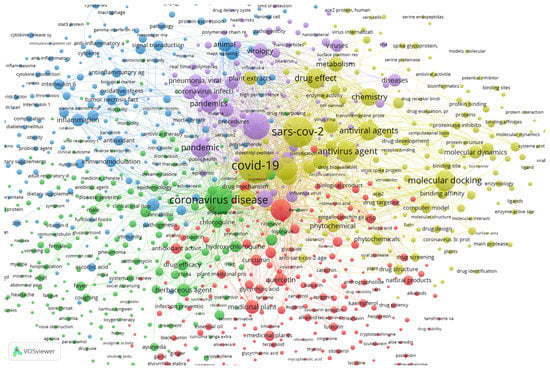
Figure 3.
Plot from VOSviewer showing the bibliographic data from specific keywords between 2020 and 2022, totaling 1504 documents grouped by co-occurrence.
Figure 3 was obtained by analyzing the co-occurrence of bibliographic data obtained from the previously mentioned keywords. The relatedness of items is determined based on the number of documents in which they occur together, with a minimum of 5 occurrences of a keyword to be considered. The visualization weight was set at the total link strength and showed a maximum of 1000. Regarding the relationship between the keywords and plants and metabolites, some specific compounds have connections with other keywords, showing that considerable amounts of studies used those compounds as potential antiviral candidates.
The lines connecting the keywords show a strong connection between them, while isolated circles mean that although the keywords occur together, there are fewer documents with both, implying a lower correlation between them, and thus these compounds have less importance. This can also be observed by the physical distance between the circles. Examples of compounds highly related to the defined keywords and with connecting lines are quercetin, luteolin, kaempferol, carvacrol and gallic acid, shown in Figure 4.
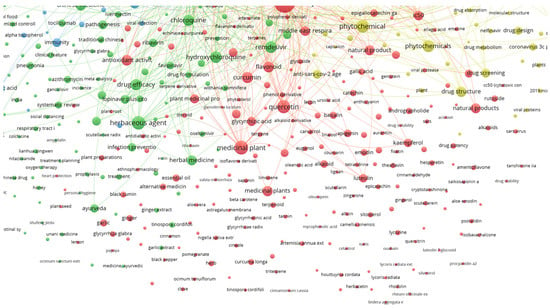
Figure 4.
Detail of relationships between plant secondary metabolites and the searched keywords.
While specific metabolites do occur in Figure 4, there could be many others that are still under research and have not been published yet. The time frame set to build Figure 3 and Figure 4 was set between 2020 and 2022, which is a very short period considering the time the research takes to achieve the results, followed by the writing and publication of the study. Still, the fact that some individual compounds already show bibliographic correlation is impressive and reveals that there is potential. Furthermore, keywords such as medicinal plant, flavonoid, phytochemicals, natural product, flavanone derivative and phytotherapy are all correlated with the keywords, but do not reveal the specific used metabolite or extract, thus demanding further rummage and increasing the potential of natural products in fighting SARS-CoV-2.
5. Antiviral Activity of Natural Sources
Nature has proven to be an inexhaustible source of molecules and compounds for several health products, namely, drugs, remedies and other related formulations [45]. Among the most well-known and studied compounds with biological activity obtained from natural products, the phenolic compounds stand out as one of the classes with higher potential, including as natural antivirals. Table 3 summarizes some of the available information regarding the antiviral activity attributed to them observed in vitro. There are no apparent specific subgroups with higher activity than others; thus, the chemical structure may influence their bioactivity. For example, some extracts rich in phenolic glucosides, obtained from the poplar tree cultivar Beaupré (Populus trichocarpa Torr. & A. Gray ex. Hook.), were effective against the Poliomyelitis virus or the Semliki Forest virus [46], and the presence of free hydroxyl and ether groups seems to influence the anti-rabies activity of phenolic compounds. Natural extracts obtained from rose root (Rhodiola rosea L.) aerial parts revealed the presence of gallic and ellagic acids, two structurally related compounds, being identified as Ebola virus entry inhibitors [1]. In turn, the aqueous extracts from Indian madder (Rubia cordifolia L.) aerial parts showed anti-viral effects against rotavirus. Sun et al. [47] identified xanthopurpurin and vanillic acid as the main ones responsible for the anti-viral effect against the human immunodeficiency virus. Some HIV-related studies have also shown that kaempferol, and mainly kaempferol-7-O-glucoside, extracted from Securigera securidaca (L.) Degen & Dörfl., could be considered as a new potential drug candidate for the treatment of HIV infection [48].
There are also several studies describing the bioactivity of phenolic extracts against the herpes virus (Table 3). Extracts rich in gallic acid (phenolic acid), apigenin (flavone), and naringin (flavanone-7-O-glycoside) appear to be effective against the herpes simplex virus type 1 (HSV-1) [49,50]. Other phenol derivatives such as p-quinone monooximes (obtained from the basic nitrosation of 3-methoxyphenol), and 8-hydroxyquinoline also showed effects in vitro against HSV-1 [51].
Phenolic compounds have also been shown to have antiviral potential against viruses that affect the respiratory system (Table 3). Özçelik et al. [49], verified that an alkaloid called atropine (commercially available) has a remarkable inhibitory effect in vitro against parainfluenza virus type 3 (PI-3), while also being effective against HSV-1. The authors confirmed that genistein (isoflavone), and the phenolic acids gallic, chlorogenic, and quinic, exerted varying degrees of anti-PI-3 effects. Some fractions isolated from the ethanol extract from Origanum vulgare L. also revealed some activity against the respiratory syncytial virus (RSV). However, this activity was weak to moderate when compared with positive controls (commercial drugs).
The compounds identified in these fractions were apigenin and a flavone glycoside identified as acacetin 7-O-[4‴-O-acetyl-β-d-apiofuransyl-(1→3)]-β-d-xylopyranoside [50]. Extracts obtained from the flowers of Bombax ceiba L., particularly rich in the flavonoids quercetin and kaempferol-3-O-(6″-O-E-p-coumaroyl)-β-d-glucopyranoside, and the xanthonoid mangiferin, also revealed anti-RSV activity in vitro [52].
Still within the topic of respiratory viruses, several studies have been carried out to verify the effect of natural extracts rich in phenolic compounds in influenza viruses. Nile et al. [53] isolated quercetin 3-glucoside from fringed pink (Dianthus superbus L.) leaves, performing tests in vitro and in silico that revealed this flavonoid might act as a natural anti-influenza drug. Takeda et al. [54] analyzed the antiviral activity of hibiscus (Hibiscus sabdariffa L.) tea extract against the human Influenza A virus (IAV). The authors evaluated its potential as a novel anti-IAV drug and a safe inactivating agent for a whole inactivated vaccine. The in vitro study revealed that the pH of the hibiscus tea extract is acidic, and its rapid and potent antiviral activity depends mainly on the acidic pH. Given this characteristic, the extract is not suitable for therapeutic or vaccination purposes. However, given the potential of hibiscus tea extract and protocatechuic acid, one of the main components of the extract, with potent acid-dependent antiviral activity, the authors suggest further study of the low-pH-independent antiviral mechanism and attempts to enhance the antiviral activity may establish a novel anti-IAV therapy and vaccination strategy. In another study, You et al. [55] verified that the phenolic compounds present in a fraction of the aqueous extract from the tender leaf of Toona sinensis (A. Juss.) M.Roem., TSL-1, might have antiviral activities against pandemic influenza A (H1N1) through the downregulation of adhesion molecules and chemokine to prevent viral attachment. The authors demonstrated that catechin might be a safe molecule for long-term use to prevent influenza A (H1N1) virus infection. On the other hand, gallic acid might be a sensitive substance to inhibit influenza virus infection.
Table 3.
Phenolic compounds antiviral activity; compound or class of compounds, type of compound or extract, extraction methodology and target virus.
Table 3.
Phenolic compounds antiviral activity; compound or class of compounds, type of compound or extract, extraction methodology and target virus.
| Phenolic Compounds/Class | Type of Compound/Extract | Extraction Methods | Virus | Reference |
|---|---|---|---|---|
| Phenolic glucosides | Ethanol extract from the leaves of the poplar tree cultivar Beaupré (Populus trichocarpa) | Ethanol extraction of the hot water-soluble portion followed by polyamide chromatography employing step-gradient elution with water and dilutions of ethanol | Poliomyelitis virusSemliki forest virus | [46] |
| p-Quinone monooximes derived from 3-methoxyphenol - 8-hydroxyquinoline | Synthetic aromatic nitro compounds | Extract washed with brine, dried over magnesium sulfate and filtered. Filtrate evaporated to dryness under reduced pressure. i-AmNO2 added to a stirred solution of a phenol in DMF in the presence of K2CO3 at 0 °C under argon. Recrystallization of the crude product from an appropriate solvent gave a p-quinone monooxime | Herpes simplex virus type 1 (HSV-1) | [51] |
| Alkyl-esters of gallic acid 3,4,5-trihydroxy derivatives of gallic acid 3,4,5-trimethoxy derivatives of gallic acid Catechin Epicatechin Quercetin Epigalocatechin | Commercial standard and lab synthesis | - | Rabies virus | [47] |
| Apigenin Naringin Atropine Genistein Gallic acid Chlorogenic acid Quinic acid | Commercial standard | Compounds dissolved in dimethyl sulfoxide to prepare a final concentration of 256 μg/mL | Herpes simplex virus type 1 (HSV-1) Parainfluenza virus type 3 (PI-3) | [49] |
| Apigenin Acacetin 7-O-[4′′′-O-acetyl-β-d-apiofuransyl-(1→3)]-β-d-xylopyranoside | Origanum vulgare L. plant extracted with 95% (v/v) ethanol | Air-dried plants percolated with 95% ethanol solution. Ethanol extract concentrated in vacuum to yield a residue, which was suspended in water and partitioned with petroleum ether and ethyl acetate, respectively | Herpes simplex virus type 1 (HSV-1) Respiratory syncytial virus (RSV) | [50] |
| Kaempferol Kaempferol-7-O-glucoside | Hydromethanolic extracts (98%) from Securigera securidaca seeds | Dried seeds extracted methanol (98%) at 40 °C. The methanol extract was eluted with n-hexane: acetone and then with 100% methanol | HIV-1 | [48] |
| Quercetin Kaempferol-3-O-(6″-O-E-p-coumaroyl)-β-d-glucopyranoside Mangiferin | Bombax ceiba L. Flowers extracted with 95% (v/v) ethanol | Extraction by reflux with 95% ethanol | Respiratory syncytial virus (RSV) | [52] |
| Xanthopurpurin (1,3-dihydroxy-9,10-anthracenedione) Vanillic acid (4-hydroxy-3-methoxybenzoic acid) | Aqueous extract from Rubia cordifolia L. aerial parts | Plant aerial parts boiled in distilled water for 1 h, the aqueous solution collected and the residual part re-extracted several times | Rotavirus | [47] |
| Catechin Gallic acid | Commercial standards | - | H1N1 Influenza virus | [55] |
| Gallic acid Ellagic acid | Aqueous and organic-solvent extracts of Rhodiola rosea L. plant | Extracts dried in vacuum at 50 °C and dissolved in DMSO | Ebola virus | [1] |
| Quercetin 3-glucoside | Hydromethanolic (70% methanol) and methanol (100%) extracts from Dianthus superbus L. leaves | Dried leaves extracted using 70% and 100% methanol | H1N1 Influenza | [53] |
| Hibiscus acid Protocatechuic acid | Acidic hibiscus tea extract | Hibiscus tea powder soaked in ultrapure water at 24 °C for 24 h, repeating the process several times | H1N1 Influenza virus | [54] |
Given the evidence of the antiviral effects of phenolic compounds, several studies have emerged regarding the effects of these compounds against SARS-CoV-2. Table 4 reports some of the tests developed specifically for the SARS coronavirus, showing the particularities mentioned above (compound or class of compounds, type of extraction and extraction methods) and the effective extract/compound concentration. While natural compounds abound, the requisite for finding suitable compounds is quite cumbersome.
These compounds must be widely available, easy to obtain in high quantities, have exceptional efficacy (above-synthesized counterparts) and be easy to isolate and synthesize (due to the pharmaceutical industry preferring synthesized compounds due to higher purity). While these traits are difficult to find in compounds, there are some examples of drugs made from plants that cannot be synthesized, or their synthesis is still too expensive to become commercially viable, even though theoretically, it is possible to synthesize all bioorganic molecules. An example of a molecule initially extracted from nature and then synthesized is Taxol, a potent anticancer drug whose structure was finally elucidated in 1994, allowing its use against cancer [56]. Nevertheless, even after complete compound synthesis is achieved, it may not be commercially viable. The time from its discovery and elucidation of the chemical structure until its commercial success becomes an academic effort with the probability of finding analogs of the original drug with equal or better effects. For example, Halichondrin B, a macrolide with antitumor activity obtained from a marine sponge, was discovered in 1986. However, the molecule’s structure and consequent total synthesis were achieved in 1992. The long period in which these tests took place, and the low synthesis yields, led to the discovery of an analogous compound, Eribulin, with much higher yields [57]. From 1981 to 2010, of all new drugs discovered, 28% were modified natural products, 30% mimicked natural products, and 6% were natural products. Furthermore, 64% of the new pharmaceuticals that entered the market were related to natural products, showing the weight that these molecules can have [58].
The entry steps of the SARS-CoV-2 virus into host cells encompass the attachment of the spike (S) glycoprotein to its receptor, angiotensin-converting enzyme 2 (ACE2), and subsequent membrane fusion. Thus, one of the strategies to inhibit the virus’s entry is finding compounds that can bind the S protein, preventing membrane attachment. Yi et al. [59] identified two small molecules from Chinese herbs with this capacity, tetra-O-galloyl-β-D-glucose (TGG) and luteolin. TGG was 50% effective in the inhibition assays with a concentration of 4.5 μM and a selective index of 240.0. On the other hand, luteonin was 50% effective with a concentration of 10.6 μM and a selective index of 14.62. The screening method developed by the authors not only identified these two compounds but also allowed several small molecules to be obtained that can be used to create new classes of anti-SARS-CoV drugs and be potentially useful for screening drugs that inhibit the entry of other viruses into host cells.
In addition to inhibiting the membrane binding process, another way to prevent the virus from growing in host cells is to inhibit essential enzymes such as the SARS-CoV 3CLpro and PLpro viral cysteine proteases. Some natural compounds, including phenolics, indeed have this capacity, as will be discussed below.
In a study carried out by Lin et al. [60], the authors tested the capacity of some phenolic compounds to inhibit the 3C-like protease (3CLpro) of SARS-CoV. It is important to highlight that this protein mediates the proteolytic processing of replicase polypeptides 1a and 1ab into functional proteins and, therefore, it has become an important target for drug development. The authors identified aloe-emodin, an anthraquinone, and hesperetin, a flavanone, obtained from the roots of Isatis indigotica Fortune, as two compounds that inhibit the cleavage activity of the 3CLpro in a dose-dependent way (Table 4). Ho et al. [61] also studied the anthraquinone emodin derived from Rheum officinale Baill. and Polygonum multiflorum Thunb., reporting that this significantly blocked the S protein and ACE2 interaction in a dose-dependent manner, suggesting that emodin may be considered a potential therapeutic agent in the treatment of SARS.
Park et al. [62] performed a biological evaluation on nine phlorotannins isolated from the edible brown algae Ecklonia cava Kjellman. The nine isolated phlorotannins, except phloroglucinol, possessed SARS-CoV 3CLpro inhibitory activities in a dose-dependent and competitive manner. Among them, two eckol groups with a diphenyl ether linked, dieckol, showed the most potent SARS-CoV 3CLpro trans/cis-cleavage inhibitory effects (IC50 = 2.7 and 68.1 μM, respectively). Moreover, dieckol exhibited a high association rate in the SPR (surface plasmon resonance) sensorgram and formed extremely strong hydrogen bonds to the catalytic dyad (Cys145 and His41) of the SARS-CoV 3CLpro. Park et al. [63] studied nine alkylated chalcones isolated from Angelica keiskei for their inhibitory activity against SARS-CoV proteases 3CLpro and PLpro. Of the isolated alkylated chalcones, chalcone 6, containing the perhydroxyl group, exhibited the most potent 3CLpro and PLpro inhibitory activity with IC50 values of 11.4 and 1.2 µM. With this work, the authors proved that the chalcones exhibited competitive inhibition characteristics to the SARS-CoV 3CLpro, whereas noncompetitive inhibition was observed with the SARS-CoV PLpro. Park et al. [64] studied the inhibitory activity of Broussonetia papyrifera-derived polyphenols against 3CLpro and PLpro cysteine proteases. The authors obtained the isolated compounds broussochalcone B, broussochalcone A, 4-hydroxyisolonchocarpin, papyriflavonol A, 3′-(3-methylbut-2-enyl)-3′,4,7-trihydroxyflavane, kazinol A, kazinol B, broussoflavan A, kazinol F and kazinol J. All the studied polyphenols were more potent against PLpro than against 3CLpro. Papyriflavonol A was the most potent inhibitor of PLpro with an IC50 value of 3.7 μM (Table 4).
The lipophilic compounds tanshinones derived from Salvia miltiorrhiza Bunge were also found to be specific and selective inhibitors for the SARS-CoV 3CLpro and PLpro viral cysteine proteases (Park et al., 2012), the activity was significantly affected by subtle changes in structure. The IC50 values of these inhibitors, although higher than those of peptide-derived and small molecule viral cysteine protease inhibitors, were nonetheless in the low micromolar (from 0.8 to 30.0 μM).
Another class of compounds studied for anti-Sars-Cov-2 activity is the saikosaponins (triterpenoid glycosides). Cheng et al. [65] tested the anticoronaviral activity of saikosaponins A, B2, C and D and their mode of action. All these saikosaponins demonstrated antiviral activity in vitro, with the strongest activity being noted for saikosaponin B2. Furthermore, saikosaponin B2 also showed an inhibitory effect on viral attachment and penetration. In fact, Bahbah et al. [66] proposed saikosaponins A, B and D derived from Bupleurum falcatum L. (Umbelliferae) for the treatment of COVID-19. The authors stated in their published letter that a molecular docking study demonstrated that saikosaponin A has a high affinity to bind to a target receptor of the SARS-CoV-2, the ACE II receptor.
There are also studies testing the ability of natural extracts, instead of isolated compounds, to inhibit SARS-CoV-2 proteases. Luo et al. [67] tried to clarify the capacity of inhibition of the compounds from Rheum palmatum L. on the SARS-3CL protease. The authors verified that the hydroethanolic extracts from R. palmatum had a high level of anti-SARS-CoV 3CL protease activity. One of the extracts, RH121, obtained after extraction with ethyl acetate and separated by silica gel column chromatography with a gradient of chloroform/methanol (10:0-0:10, v/v), showed an IC50 value of 13.76 µg/mL and an inhibition rate up to 96%. Therefore, Luo et al. [67] concluded that the extracts from R. palmatum have a high level of inhibitory activity against 3CL protease, suggesting these may represent a potential therapeutic for SARS. Kim et al. [68] evaluated the effects of the ethanol extract of the seeds of Psoralea corylifolia L., which revealed high activity against the SARS-CoV PLpro with an IC50 of 15 µg/mL. Given its potential, the authors fractionated the extract, obtaining six aromatic compounds, namely, bavachinin, neobavaisoflavone, isobavachalcone, 4′-O-methylbavachalcone, psoralidin and corylifol A. All the isolated flavonoids inhibited PLpro in a dose-dependent manner with IC50 (concentration causing 50% of inhibition) ranging between 4.2 and 38.4 µM.
Finally, Schwarz et al. [69] based their research on the fact that the protein coded by the open-reading-frame 3a of SARS coronavirus forms a cation-selective channel that may become expressed in the infected cell, and the activity of the channel is involved in the mechanism of virus release. In this way, the authors tested the flavonols kaempferol, kaempferol glycosides and acylated kaempferol glucoside derivatives from the leaves of the plant Quercus ilex L. and kaempferol triglycoside from Viola odorata L. for their capacity to block the 3a channel and, therefore, inhibit virus release. The most effective one was the glycoside juglanin (carrying an arabinose residue) for inhibition of the 3a-mediated current, with an IC50 value of 2.3 µM. Kaempferol derivatives with rhamnose residue also seem to be quite effective (Table 4).
Table 4.
Anti-SARS coronavirus from phenolic and other compounds, type of extraction, extraction methodology and concentrations with activity.
Table 4.
Anti-SARS coronavirus from phenolic and other compounds, type of extraction, extraction methodology and concentrations with activity.
| Phenolic Compounds/Class | Type of Extract | Extraction Methods | Concentration | Reference |
|---|---|---|---|---|
| Tetra-O-galloyl-beta-D-glucose (TGG) Luteolin | Hydroethanolic extract | Herbs extracted by maceration with 85% ethanol at room temperature for 2 weeks | 4.5 µM 83.4 µM | [59] |
| Aloe emodin Hesperetin | Aqueous extract from Isatis indigotica roots | Plant roots extracted twice with 10 volumes of distilled boiling water for 1 h | 132 μM, 366 μM and 911.592 μM 60 μM, 8.3 μM and 2718 μM | [60] |
| Saikosaponins (A, B2, C and D) | Commercial standards | Saikosaponins dissolved in DMSO and further diluted with RPMI 1640 medium | 25 µmmol | [70] |
| A. emodin | Aqueous extract of the root tuber from Rheum officinale Baill. and Polygonum multiflorum Thunb. | Deionized water | IC50 values ranged from 1 to 10 μg/mL. | [61] |
| Flavonoids from a Chinese multiherb remedy constituted by Herba Houttuyniae, Flos Chrysanthemi Indici, Herba Artemisiae Scopariae, Herba Eupatorii and Fructus Tsaoko | Hydroethanolic extract | Herb mixture extracted with 95% EtOH at room temperature | From the nine flavonoids with inhibitory effects, luteolin was the most potent with a CH50 (50% inhibitory concentration) of 0.19 mM | [71] |
| n.a. | Hydroethanolic extracts of Rheum palmatum L. (ethanol (75%)) | Petroleum ether and chloroform, and ethyl acetate | Among the extracts, RH121 has the highest activity, with an IC50 of 13.76 μg/mL | [67] |
| Tanshinones | Hydroethanolic extract (95%) of the dried roots of Salvia miltiorrhiza Bunge | The crude extract was filtered and evaporated under reduced pressure. The obtained residue was suspended with distilled water | IC50 value of 0.7µM | [65] |
| Phlorotannins | Ethanol extract of brown alga Ecklonia cava Kjellman | Ethanol at room temperature | Of the nine phlorotannins tested, two eckol groups with a diphenyl ether linked dieckol showed the most potent SARS-CoV 3CLpro trans/cis-cleavage inhibitory effects; IC50 = 2.7 and 68.1 μM, respectively) | [62] |
| Flavonoids (bavachinin, neobavaisoflavone, isobavachalcone, 4′-O-methylbavachalcone, psoralidin and corylifol A) | Fractionation of hydroethanolic extract from the seeds of Psoralea corylifolia L. | Ethanol, water and n-hexane | IC50 ranging between 4.2 and 38.4 µM | [68] |
| Juglanin Kaempferol-3-O-α-rhamnopyranosyl(1→2) [α-rhamnopyranosyl(1→6)]-β-glucopyranoside | Kaempferol acylated glucosides were previously isolated from polar extracts from the leaves of the plant Quercus ilex L.; the kaempferol triglycoside was an isolate from Viola odorata L (decoction and infusion) | Cyclohexane, Et2O, MeOH and MeOH–H2O | The most effective one was the glycoside juglanin with an IC50 of 2.3 µM | [69,72] |
| Alkylated chalcones | Hydroethanolic extract from the leaves of Angelica keiskei Ito | 95% ethanol for a week at room temperature | IC50 values of 11.4 (3CLpro) and 1.2 µM (PLpro) | [63] |
| Broussochalcone B, Broussochalcone A, 4-hydroxyisolonchocarpin, Papyriflavonol A, 3′-(3-methylbut-2-enyl)-3′,4,7-trihydroxyflavane, Kazinol A, Kazinol B, Broussoflavan A, Kazinol F, Kazinol J | Ethanolic extracts from the roots of Broussonetia papyrifera (L.) Vent. | Ethanol at room temperature and evaporated using a rotary evaporator at temperatures below 45 °C to obtain the total extract. | Papyriflavonol A was the most potent inhibitor of PLpro with an IC50 of 3.7 μM. | [64] |
| Epicatechin 5-O-beta-D-glucopyranoside-3-benzoate Neohesperidin Kaempferol-3,7-O-dirhamnoside (Kaempferitrin) Quercetin-3-O-neohesperidoside Oleic acid 3,4,5-Trimethoxyphenol Epicatechin Oxypeucedanin hydrate 3-Feruloylquinic acid Eriodictyol Apigenin Luteolin C-Methyl flavone β-amyrin Isovitexin-2″-O-rhamnoside | Methanolic extracts from Fragaria ananassa Duch. | 80% methanol at room temperature. | Strawberry methanolic extract showed the highest antiviral activity against SARS-CoV-2 with an IC50 value to 0.0062 µg/mL | [2] |
| Sinapic acid | Ethanolic extract of broccoli (obtained from a local market) | 95% ethanol | Potent SARS CoV-2 inhibition with a half-maximal inhibitory concentration (IC50) value of 2.69 µg/mL | [73] |
| Derivatives of luteolin, kaempferol, apigenin, isorhamnetin, myricetin, chrysoeriol, biochanin, isookanin and scutellarein | V. vinifera (var. Paulsen 1103) leaf extract | 75% (v/v) methanol/0.05% (v/v) trifluoroacetic acid | Leaf extract was able to inhibit both HSV-1 and SARS-CoV-2 replication in the early stages of infection by directly blocking the proteins enriched on the viral surface, at a very low concentration of 10 µg/mL | [74] |
| TFC of sampled wild S. nigra was 9.57 ± 0.65 mg RE g−1 DW of plant material for berry extracts and 77.59 ± 10.23 mg RE g−1 DW of plant material for flower extracts. TPC of sampled wild S. nigra was 41.31 ± 9.44 mg gallic acid equivalent (GAE) g−1 DW for berry extracts and 451.72 ± 25.31 mg GAE g−1 DW for flower extracts | Ethanolic Extract of Sambucus nigra L. berry and flowers | 80 % ethanol at 60 °C | Concentration-dependent inhibition of ACE2-SARS-CoV2 S-protein RBD binding was demonstrated in vitro for elderberry fruits and flowers extracts (IC50 of 1.66 mg DW ml−1 and 0.532 mg DW ml−1, respectively) | [75] |
| Caffeic acid, caftaric acid, chlorogenic acid, cichoric acid, cynarin, echinacoside | Echinacea purpurea (echinaforce) | 65% ethanol | 50 µg/mL inactivation of SARS-CoV1 and 2 | [76] |
| Glyzyrrhizin | Glycyrrhiza glabra | Acquired compound | 300 mg/L | [77] |
| Baicalein | Scutellaria baicalensis | 70% ethanol (crude extract of plant) and commercial baicalein | IC50 8.52 µg/mL (crude extract) and 0.39 µg/mL (baicalein) | [78] |
| Epigallocatechin, genistein, sulforaphane, chlorogenic acid, resveratrol and quercetin | - | Acquired compounds | IC50—33.9 µg/mL (epigallocatechin) | [79] |
| Whole plant extract | Taraxacum officinale | Aqueous extractions for 1 h | EC50—14.9 mg/mL | [80] |
The latest reviews demonstrate that extracts or preparations with inhibitory activity against coronaviruses include Lycoris radiata (L’Hér.) Herb., Artemisia annua L., Pyrrosia lingua (Thunb.) Farw., Lindera aggregata (Sims) Kosterm. and Isatis indigotica Fortune, and the extracts of Rheum officinale Baill., Polygonum multiflorum Thunb., Houttuynia cordata Thunb., Gentiana scabra Bunge, Dioscorea batatas L., Cassia tora L., Taxillus chinensis (DC.) Danser, Cibotium barometz (L.) J.Sm., Anthemis hyaline DC., Nigella sativa L. and Citrus sinensis (L.) Osbeck [45]. Furthermore, regarding the class of bioactive compounds that act against coronaviruses, alkaloids, phenolic compounds including flavonoids, chalcones, lignins and tannins, glycosides, stand out as the most promising agents [45].
7. Future and Trends
The relationship between viruses and humans has always been about balancing infectiousness and lethality, and while outbreaks of highly contagious and lethal viruses have been few, recent studies have shown that they are expected to increase in coming years. Improvement of hygiene and knowledge of viruses has helped mankind to protect itself from viruses, but some pockets of densely packed population and wild animals has created the perfect concoction for spillover of viruses and other pathogens from animals to humans, and while this could be relatively easy to overcome, other changes are more cumbersome.
Climate change, which represents a huge issue on its own [100], is probably the most important factor that will increase the pressure for viruses such as the Coronaviridae to jump from animals to humans [101]. Deforestation increases pressure on wild ecosystems, making wild animals share habitats with humans. Beyond this, floods increase the prevalence of viral vectors, and droughts change ecosystems, making them abruptly shift, allowing for new connections between animals, resulting in a mixture of viruses that can easily infect humans.
Beyond the grim scenario of climate change helping to increase the prevalence of viruses, the unknown long-standing symptoms, and opportunistic diseases in patients previously infected with COVID-19 is another matter of concern. Autoimmune diseases are manifesting themselves in some patients after recovering from mild to severe COVID-19, which shows the long road ahead for satisfactory knowledge of this illness and its sequelae over time [102,103,104].
While mitigating climate change may help reduce the probability of higher occurrences of a global pandemic, it will not avoid them. In fact, it is hardly possible to completely avoid contained outbreaks of viral pandemics, and thus, mankind can only work to reduce their widespread and be prepared to quickly implement vaccines, treatments and measures to mitigate their effects, which could control or even preview further outbreaks. These efforts must be carried out by all branches of society, and all possibilities should be pondered.
Plants, being a widely disseminated and endless resource, and are in some cases the cure or treatment for many previous diseases and pathogenic vectors, are once again paramount to fend of this coronavirus, and will be effective in the coming pandemics.
The use of plant extracts as adjuvants in antiviral treatments, as accessory treatment for symptomatic relief, as application in protective equipment for health professionals, and being mixed into disinfectant solutions and other applications is still being studied, and results of publications and intellectual property are expected to be rolled out in the coming months and years.
Author Contributions
Writing—review and editing, S.A.H.; writing—original draft, M.C.; investigation, S.A.H., M.C., F.S.R. and T.C.S.P.P.; funding acquisition, I.C.F.R.F.; project administration, L.B. and M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Portuguese Fundação para Ciência e Tecnologia (FCT), which financed the Centro de Investigação de Montanha through (UIDB/00690/2020), as well as the researchers S. Heleno and M. Carocho (CEECIND/00831/2018, CEECIND/03040/2017) as well as L. Barros. The research was also funded by the Programa Operacional Regional Norte 2020, within the “PlantCovid” project, NORTE-01-02B7-FEDER-054870.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data was produced.
Acknowledgments
The authors are grateful to the Foundation for Science and Technology (FCT, Portugal) for financial support through national funds FCT/MCTES to the CIMO (UIDB/00690/2020). Acknowledgments to the European Fund for Regional Development through the Programa Operacional Regional Norte 2020, within the “PlantCovid” project, NORTE-01-02B7-FEDER-054870. S. Heleno and M. Carocho thank FCT for their individual employment program-contract (CEECIND/00831/2018, CEECIND/03040/2017). L. Barros also thanks the national funding by FCT, through the institutional scientific employment program-contract for her contract.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adedeji, A.O.; Severson, W.; Jonsson, C.; Singh, K.; Weiss, S.R.; Sarafianos, S.G. Novel inhibitors of severe acute respiratory syndrome coronavirus entry that act by three distinct mechanisms. J. Virol. 2013, 87, 8017–8028. [Google Scholar] [CrossRef]
- Al-Sanea, M.M.; Abelyan, N.; Abdelgawad, M.A.; Musa, A.; Ghoneim, M.M.; Al-Warhi, T.; Aljaeed, N.; Alotaibi, O.J.; Alnusaire, T.S.; Abdelwahab, S.F.; et al. Strawberry and ginger silver nanoparticles as potential inhibitors for SARS-CoV-2 sssisted by in silico modeling and metabolic profiling. Antibiotics 2021, 10, 824. [Google Scholar] [CrossRef]
- Artika, I.M.; Dewantari, A.K.; Wiyatno, A. Molecular biology of coronaviruses: Current knowledge. Heliyon 2020, 6, e04743. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Drosten, C.; Doerr, H.; Stürmer, M.; Preiser, W. Severe acute respiratory syndrome (SARS)-paradigm of an emerging viral infection. J. Clin. Virol. 2004, 29, 13–22. [Google Scholar] [CrossRef]
- Raj, V.S.; Osterhaus, A.D.; Fouchier, R.A.; Haagmans, B.L. MERS: Emergence of a novel human coronavirus. Curr. Opin. Virol. 2014, 5, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 transmission and pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef]
- World Health Organization—WHO. Middle East Respiratory Syndrome Coronavirus (MERS-CoV). 2020. Available online: https://www.who.int/news/item/27-07-2017-countries-agree-next-steps-to-combat-global-health-threat-by-mers-cov/ (accessed on 26 July 2021).
- Chen, Z.; Boon, S.S.; Wang, M.H.; Chan, R.W.Y.; Chan, P.K.S. Genomic and evolutionary comparison between SARS-CoV-2 and other human coronaviruses. J. Virol. Methods 2021, 289, 114032. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Chapter 3—Morphology, genome organization, replication, and pathogenesis of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). In Coronavirus Disease 2019 (COVID-19); Saxena, S.K., Ed.; Springer: Singapore, 2020. [Google Scholar]
- Schoeman, D.; Fielding, B.C. Coronavirus envelope protein: Current knowledge. Virol. J. 2019, 16, 69. [Google Scholar] [CrossRef]
- De Wit, E.; Doremalen, N.; Falzarano Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Low, J.S.; Jerak, J.; Tortorici, M.A.; McCallum, M.; Pinto, D.; Cassotta, A.; Foglierini, M.; Mele, F.; Abdelnabi, R.; Weynand, B.; et al. ACE-2-biinding exposes the SARS-CoV-2 fusion peptide to broadly neutralizing coronavisus antibodies. Science 2022, 377, 735–742. [Google Scholar] [CrossRef]
- Munir, M.; Tandiabang, P.; Setyawati, T.; Basry, A.; Cyio, A.; Rahman, N. Bioethical perspective of convalescent plasma therapy for COVID-19: A systematic review. Transfus. Clin. Et Biol. 2021, 28, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alba, E.; Nuzzolo-Shihadeh, L.; Aguirre-García, G.M.; Espinosa-Mora, J.; Lecona-Garcia, J.D.; Flores-Pérez, R.O.; Mendoza-Garza, M.; Camacho-Ortiz, A. Baricitinib plus dexamethasone compared to dexamethasone for the treatment of severe COVID-19 pneumonia: A retrospective analysis. J. Microbiol. Immunol. Infect. 2021, 54, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Snow, T.A.C.; Saleem, N.; Ambler, G.; Nastouli, E.; McCoy, L.E.; Singer, M.; Arulkumaran, N. Convalescent plasma for COVID-19: A meta-analysis, trial sequential analysis, and meta-regression. Br. J. Anaesth. 2021, 127, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.P.K.; Dangerfield, T.L.; Taylor, D.W.; Johnson, K.A. Remdesivir is a delayed translocation inhibitor of SARS-CoV-2 replication. Mol. Cell 2021, 81, 1548–1552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.R.; Zhang, Y.N.; Li, X.D.; Zhang, H.Q.; Xiao, S.Q.; Deng, F.; Yuan, Z.M.; Ye, H.Q.; Zhang, B. A cell-based large-scale screening of natural compounds for inhibitors of SARS-CoV-2. Signal Transduct. Target. Ther. 2020, 5, 218. [Google Scholar] [CrossRef]
- Kyriazopoulou, E.; Huet, T.; Cavalli, G.; Gori, A.; Kyprianou, M.; Pickkers, P.; Eugen-Olsen, J.; Clerici, M.; Veas, F.; Chatellier, G.; et al. Effect of anakinra on mortality in patients with COVID-19: A systematic review and patient-level meta-analysis. Lancet Rheumatol. 2021, 10, E690–E697. [Google Scholar] [CrossRef]
- Taramasso, L.; Magnasco, L.; Portunato, F.; Briano, F.; Vena, A.; Giacobbe, D.R.; Dentone, C.; Robba, C.; Ball, L.; Loconte, M.; et al. Clinical presentation of secondary infectious complications in COVID-19 patients with intensive care unit treated with tocilizumab or standard of care. Eur. J. Intern. Med. 2021, 94, 39–44. [Google Scholar] [CrossRef]
- Dougan, M.; Nirula, A.; Azizad, M.; Mocherla, B.; Gottlieb, R.L.; Chen, P.; Hebert, C.; Perry, R.; Boscia, J.; Heller, B.; et al. Bamlanivimab plus etesevimab in mild or moderate Covid-19. N. Engl. J. Med. 2021, 385, 1382–1392. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jang, Y.R.; Hong, J.H.; Jung, J.G.; Park, J.; Streinu-Cercel, A.; Streinu-Cercel, A.; Săndulescu, O.; Lee, S.J.; Kim, S.H.; et al. Safety, virologic efficacy, and pharmacokinetics of CT-P59, a neutralizing monoclonal antibody against SARS-CoV-2 spike receptor-binding protein: Two randomized, placebo-controlled phase 1 studies in healthy subjects and patients with mild SARS-CoV-2 infection. Clin. Ther. 2021, 43, 1706–1727. [Google Scholar]
- Razonable, R.R.; Pawlowski, C.; O'Horo, J.C.; Arndt, L.L.; Arndt, R.; Bierle, D.M.; Borgen, M.D.; Hanson, S.N.; Hedin, M.C.; Lenehan, P.; et al. Casirivimab-Imdevimab treatment is associated with reduced rates of hospitalization among high-risk patients with mild to moderate coronavirus disease-19. EClinicalMecdicine 2021, 40, 101102. [Google Scholar] [CrossRef]
- Baral, P.K.; Yin, J.; James, M.N.G. Treatment and prevention strategies for the COVID 19 pandemic: A review of immunotherapeutics approaches for neutralizing SARS-CoV-2. Int. J. Biol. Macromol. 2021, 186, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hofmann-Winkler, H.; Krüger, N.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Arora, P.; Sidarovich, A.; Moldenhauer, A.-S.; Winkler, M.S.; et al. SARS-CoV-2 variant B.1.617 is resistant to bamlanivimab and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 36, 109415. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Alhatlani, B. An overview of current COVID-19 vaccine platforms. Comput. Struct. Biotechnol. J. 2021, 19, 2508–2517. [Google Scholar] [CrossRef]
- Ceron, W.; De-Lima-Santos, M.-F.; Quiles, M.G. Fake news agendas in the era of COVID-19: Identifying trends through fact-checking content. Online Soc. Netw. Media 2021, 21, 100116. [Google Scholar] [CrossRef]
- Gautam, S.; Gautam, A.; Chhetri, S.; Bhattarai, U. Immunity against COVID-19: Potential role of Ayush Kwath. J. Ayurveda Integr. Med. 2020, 13, 100350. [Google Scholar] [CrossRef]
- Hu, H.; Ji, Z.; Feng, C.; Pang, W.; Chen, Z.; Zhang, J.; Wang, H. PROSPERO’s systematic review protocols of traditional Chinese medicine for COVID-19: An overview. Integr. Med. Res. 2021, 10, 100774. [Google Scholar] [CrossRef]
- Liang, S.-B.; Fang, M.; Liang, C.-H.; Lan, H.-D.; Shen, C.; Yan, L.-J.; Hu, X.-Y.; Han, M.; Robinson, N.; Liu, J.-P. Therapeutic effects and safety of oral Chinese patent medicine for COVID-19: A rapid systematic review and meta-analysis of randomized controlled trials. Complement. Ther. Med. 2021, 60, 102744. [Google Scholar] [CrossRef]
- Orisakwe, O.E.; Orish, C.N.; Nwanaforo, E.O. Coronavirus disease (COVID-19) and Africa: Acclaimed home remedies. Sci. Afr. 2020, 10, e00620. [Google Scholar] [CrossRef]
- Ren, W.; Liang, P.; Ma, Y.; Sun, Q.; Pu, Q.; Dong, L.; Luo, G.; Mazhar, M.; Liu, J.; Wang, R.; et al. Research progress of traditional Chinese medicine against COVID-19. Biomed. Pharmacother. 2021, 137, 111310. [Google Scholar] [CrossRef]
- Yin, X.; Cai, S.-B.; Tao, L.-T.; Chen, L.-M.; Zhang, Z.-D.; Xiao, S.-H.; Fan, A.Y.; Zou, X. Recovery of a patient with severe COVID-19 by acupuncture and Chinese herbal medicine adjuvant to standard care. J. Integr. Med. 2021, 19, 460–466. [Google Scholar] [PubMed]
- Zhu, W.; Xu, M.; Chen, C.Z.; Guo, H.; Shen, M.; Hu, X.; Shinn, P.; Klumpp-Thomas, C.; Michael, S.G.; Zheng, W. Identification of SARS-CoV-2 3CL protease inhibitors by a quantitative high-throughput screening. ACS Pharmacol. Transl. Sci. 2020, 3, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Feyaerts, A.F.; Luyten, W. Vitamin C as prophylaxis and adjunctive medical treatment for COVID-19? Nutrition 2020, 79, 110948. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.T.M.R.; Ferdousi, J.; Shahinozzaman, M. Previously published ethnopharmacological reports reveal the potentiality of plants and plant-derived products used as traditional home remedies by Bangladeshi COVID-19 patients to combat SARS-CoV-2. Saudi J. Biol. Sci. 2021, 28, 6653–6673. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Fan, G.; Xiao, G.; Wang, T.; Xu, D.; Gao, J.; Ge, S.; Li, Q.; Ma, Y.; Zhang, H.; et al. Traditional chinese medicine in COVID-19. Acta Pharm. Sin. B 2021, 11, 3337–3363. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, H.; Feehan, J.; Dhaheri, A.S.A.; Ali, H.I.; Platat, C.; Ismail, L.C.; Apostopoulos, V.; Stojanovska, L. Immune-boosting role of vitamins D, C, E, zinc, selenium and moega-3 fatty acids: Could they help against COVID-19? Maturitas 2021, 143, 1–9. [Google Scholar] [CrossRef]
- Gasmi, A.; Peana, M.; Noor, S.; Lysiuk, R.; Menzel, A.; Benahmed, A.G.; Bjørklund, G. Chloroquine and hydroxychloroquine in the treatment of COVID-19: The never-ending story. Appl. Microbiol. Biotechnol. 2021, 105, 1333–1343. [Google Scholar] [CrossRef]
- Di Stefano, L.; Ogburn, E.L.; Ram, M.; Scharfstein, D.O.; Li, T.; Khanal, P.; Baksh, S.N.; McBee, N.; Gruber, J.; Gildea, M.R.; et al. Hydroxychlorooquine/chloroquine for the treatment of hospitalized patients with COVID-19: An individual participant data meta-analysis. PLoS ONE 2022, 17, e0273526. [Google Scholar] [CrossRef]
- Réa-Neto, Á.; Bernardelli, R.S.; Câmara, B.M.D.; Reese, F.B.; Queiroga, M.V.O.; Oliveira, M.C. An open-label randomized controlled trial evaluating the efficacy of cholorquine/hydroxychloroquine in severe COVID-19 patients. Sci. Rep. 2021, 11, 9023. [Google Scholar] [CrossRef]
- Reis, G.; Silva, E.A.S.M.; Silva, D.C.M.; Thabane, L.; Milagres, A.C.; Ferreira, T.S.; Santos, C.V.Q.; Campos, V.H.S.; Nogueira, A.M.R.; Almeida, A.P.F.G.; et al. Effect of early treatment with ivermectin among patients with Codid-19. N. Engl. J. Med. 2022, 386, 1721–1731. [Google Scholar] [CrossRef]
- Bramante, C.T.; Huling, J.D.; Tignanelli, C.J.; Buse, J.B.; Liebovitz, D.M.; Nicklas, J.M.; Kenneth Cohen, K.; Puskarich, M.A.; Belani, H.K.; Proper, J.L.; et al. Randomized trial of metformin, ivermectin, and fluvoxamine for Covid-19. N. Engl. J. Med. 2022, 387, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Firn, R. Are NP’S different from synthetic chemicals? In Nature’s Chemicals: The Natural Products That Shaped Our World; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Islam, M.T.; Sarkar, C.; El-Kersh, D.M.; Jamaddar, S.; Uddin, S.J.; Shilpi, J.A.; Mubarak, M.S. Natural products and their derivatives against coronavirus: A review of the non-clinical and pre-clinical data. Phytother. Res. 2020, 34, 2471–2492. [Google Scholar] [CrossRef] [PubMed]
- Van Hoof, L.; Totté, J.; Corthout, J.; Pieters, L.; Mertens, F.; Berghe, D.A.V.; Vlietinck, A.J.; Dommisse, R.; Esmans, E. Plant Antiviral Agents, VI. Plant Antiviral Agents, VI. Isolation of Antiviral Phenolic Glucosides from Populus Cultivar Beaupre by Droplet Counter-Current Chromatography. J. Nat. Prod. 1989, 52, 875–878. [Google Scholar]
- Chávez, J.H.; Leal, P.C.; Yunes, R.A.; Nunes, R.J.; Barardi, C.R.; Pinto, A.R.; Simões, C.M.; Zanetti, C.R. Evaluation of antiviral activity of phenolic compounds and derivatives against rabies virus. Vet. Microbiol. 2006, 116, 53–59. [Google Scholar] [CrossRef]
- Behbahani, M.; Sayedipour, S.; Pourazar, A.; Shanehsazzadeh, M. In vitro anti-HIV-1 activities of kaempferol and kaempferol-7-Oglucoside isolated from Securigera securidaca. Res. Pharm. Sci. 2014, 9, 463–469. [Google Scholar] [PubMed]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Guo, Y.-S.; Wang, C.-H.; Li, G.-Q.; Xu, J.-J.; Chung, H.Y.; Ye, W.-C.; Li, Y.-L.; Wang, G.-C. Phenolic compounds from Origanum vulgare and their antioxidant and antiviral activities. Food Chem. 2014, 152, 300–306. [Google Scholar] [CrossRef]
- Ishikawa, T.; Watanabe, T.; Tanigawa, H.; Saito, T.; Kotake, K.-I.; Ohashi, Y.; Ishii, H. Nitrosation of phenolic substrates under mildly basic conditions: Selective preparation of p-quinone monooximes and their antiviral activities. J. Org. Chem. 1996, 61, 2774–2779. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Wu, P.; Zhang, X.-L.; Xia, C.; Li, G.-Q.; Ye, W.-C.; Wang, G.-C.; Li, Y.-L. Phenolic compounds from the flowers of Bombax malabaricum and their antioxidant and antiviral activities. Molecules 2015, 20, 19947–19957. [Google Scholar] [CrossRef]
- Nile, S.H.; Kim, D.H.; Nile, A.; Park, G.S.; Gansukh, E.; Kai, G. Probing the effect of quercetin 3-glucoside from Dianthus superbus L. against influenza virus infection- In vitro and in silico biochemical and toxicological screening. Food Chem. Toxicol. 2020, 135, 110985. [Google Scholar] [CrossRef]
- Takeda, Y.; Okuyama, Y.; Nakano, H.; Yaoita, Y.; Machida, K.; Ogawa, H.; Imai, K. Antiviral Activities of Hibiscus sabdariffa L. tea extract against human influenza a virus rely largely on acidic pH but partially on a low-pH-independent mechanism. Food Environ. Virol. 2019, 12, 9–19. [Google Scholar] [CrossRef] [PubMed]
- You, H.-L.; Huang, C.-C.; Chen, C.-J.; Chang, C.-C.; Liao, P.-L.; Huang, S.-T. Anti-pandemic influenza A (H1N1) virus potential of catechin and gallic acid. J. Chin. Med Assoc. 2018, 81, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C. Organic synthesis: The art and science of replicating the molecules of living nature and creating others like them in the laboratory. Proc. R. Soc. 2014, 479, 20130690. [Google Scholar] [CrossRef] [PubMed]
- Crane, E.A.; Gademann, K. Capturing biological activity in natural products fragments by chemical synthesis. Angew. Chem. Int. Ed. 2016, 55, 3882–3902. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as source of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef]
- Lin, C.-W.; Tsai, F.-J.; Tsai, C.-H.; Lai, C.-C.; Wan, L.; Ho, T.-Y.; Hsieh, C.-C.; Chao, P.-D.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Park, J.-Y.; Kim, J.H.; Kwon, J.M.; Kwon, H.-J.; Jeong, H.J.; Kim, Y.M.; Kim, D.; Lee, W.S.; Ryu, Y.B. Dieckol, a SARS-CoV 3CLpro inhibitor, isolated from the edible brown algae Ecklonia cava. Bioorganic Med. Chem. 2013, 21, 3730–3737. [Google Scholar] [CrossRef]
- Park, J.-Y.; Ko, J.A.; Kim, D.W.; Kim, Y.M.; Kwon, H.J.; Jeong, H.J.; Kim, C.Y.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Chalcones isolated from Angelica keiskei inhibit cysteine proteases of SARS-CoV. J. Enzym. Inhib. Med. Chem. 2016, 31, 23–30. [Google Scholar] [CrossRef]
- Park, J.-Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef]
- Park, J.-Y.; Kim, J.H.; Kim, Y.M.; Jeong, H.J.; Kim, D.W.; Park, K.H.; Kwon, H.; Park, S.; Lee, W.S.; Ryu, Y.B. Tanshinones as selective and slow-binding inhibitors for SARS-CoV cysteine proteases. Bioorganic Med. Chem. 2012, 20, 5928–5935. [Google Scholar] [CrossRef] [PubMed]
- Bahbah, E.I.; Negida, A.; Nabet, M.S. Purposing saikosaponins for the treatment of COVID-19. Med. Hypotheses 2020, 140, 109782. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Su, X.; Gong, S.; Qin, Y.; Liu, W.; Li, J.; Yu, H.; Xu, Q. Anti-SARS coronavirus 3C-like protease effects of Rheum palmatum L. extracts. Biosci. Trends 2009, 3, 124–126. [Google Scholar] [PubMed]
- Kim, D.W.; Seo, K.H.; Curtis-Long, M.J.; Oh, K.Y.; Oh, J.W.; Cho, J.K.; Lee, K.H.; Park, K.H. Phenolic phytochemical displaying SARS-CoV papain-like protease inhibition from the seeds of Psoralea corylifolia. J. Enzym. Med. Chem. 2014, 29, 59–63. [Google Scholar] [CrossRef]
- Schwarz, S.; Sauter, D.; Wang, K.; Zhang, R.; Sun, B.; Karioti, A.; Bilia, A.R.; Efferth, T.; Schwarz, W. Kaempferol derivatives as antiviral drugs against the 3a channel protein of coronavirus. Planta Medica 2014, 80, 177–182. [Google Scholar] [CrossRef]
- Cheng, P.-W.; Ng, L.-T.; Chiang, L.-C.; Lin, C.-C. Antiviral effects of saikosaponins on human coronavirus 229E in vitro. Clin. Exp. Pharmacol. Physiol. 2006, 33, 612–616. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, D. Anticomplementary principles of a Chinese multiherb remedy for the treatment and prevention of SARS. J. Ethnopharmacol. 2008, 117, 351–361. [Google Scholar] [CrossRef]
- Karioti, A.; Bilia, A.R.; Skaltsa, H. Quercus ilex L.: A rich source of polyacylated flavonoid glycosides. Food Chem. 2010, 123, 131–142. [Google Scholar] [CrossRef]
- Orfali, R.; Rateb, M.; Hassan, H.; Alonazi, M.; Gomaa, M.; Mahrous, N.; GabAllah, M.; Kandeil, A.; Perveen, S.; Abdelmohsen, U.; et al. Sinapic Acid Suppresses SARS CoV-2 Replication by Targeting Its Envelope Protein. Antibiotics 2021, 10, 420. [Google Scholar] [CrossRef]
- Zannella, C.; Giugliano, R.; Chianese, A.; Buonocore, C.; Vitale, G.A.; Sanna, G.; Sarno, F.; Manzin, A.; Nebbioso, A.; Termolino, P.; et al. Antiviral Activity of Vitis vinifera Leaf Extract against SARS-CoV-2 and HSV-1. Viruses 2021, 13, 1263. [Google Scholar] [CrossRef] [PubMed]
- Boroduske, A.; Jekabsons, K.; Riekstina, U.; Muceniece, R.; Rostoks, N.; Ilva, N. Wild Sambucus nigra L. from north-east edge of the species range: A valuable germplasm with inhibitory capacity against SARS-CoV2 S-protein RBD and hACE2 binding in vitro. Ind. Crops Prod. 2021, 165, 113438. [Google Scholar] [CrossRef] [PubMed]
- Signer, J.; Jonsdottir, H.R.; Albrich, W.C.; Strasser, M.; Züst, R.; Ryter, S.; Ackerman-Gäumann, R.; Lenz, N.; Siegrist, D.; Suter, A.; et al. In vitro virucidal activity of Echinaforce, an Echinacea purpurea preparation, against coronaviruses, including common cold coronavirus 229E and SARS-CoV-2. Virol. J. 2020, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H. Glycyrrhizin, and active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ye, F.; Sun, Q.; Liang, H.; Li, C.; Li, S.; Lu, R.; Huang, B.; Tna, W.; Lai, L. Scutellaria baicalensis extract and baicalein inhibit replication of SARS-CoV-2 and its 3C-like protease in vitro. J. Enzym. Inhib. Med. Chem. 2021, 36, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, T.; Hishiki, T.; Baig, M.S.; Rajpoot, S.; Saqib, U.; Takasaki, T.; Hara, Y. Epigallocatechin gallate (EGCG) attenuates severe acute respiratory coronavirus disease 2 (SARS-CoV-2) infection by blocking the interaction of SARS-CoV-2 spike protein receptor-binding domain to human angiotensin-converting enzyme 2. PLoS ONE 2022, 17, e0271112. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Gigl, M.; Le, N.P.K.; Dawid, C.; Lamy, E. In vitro effect of Taraxacum officinale lear aqueous extract on the interaction between ACE2 cell surface receptor and SARS-CoV-2 spike protein D614 and four mutants. Pharmaceuticals 2021, 14, 1055. [Google Scholar] [CrossRef]
- Touret, F.; Gilles, M.; Barral, K.; Nougairède, A.; van Helden, J.; Decroly, E.; de Lamballerie, X.; Coutard, B. In vitro screening of a FDA approved chemical library reveals potential inhibitors of SARS-CoV-2 replication. Sci. Rep. 2020, 10, 13093. [Google Scholar]
- Brown, A.S.; Ackerley, D.F.; Calcott, M.J. High-throughput screening for inhibitors of the SARS-CoV-2 protease using a FRET-biosensor. Molecules 2020, 25, 4666. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Gallo, G.; Campos, C.B.; Hardy, L.; Würtele, M. Biochemical screening for SARS-CoV-2 main protease inhibitors. PLoS ONE 2020, 15, e0240079. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zheng, W.; Huang, R. High-throughput screening assays for SARS-CoV-2 drug development: Current status and future directions. Drug Discov. Today 2021, 26, 2439–2444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Qiao, W.; Zhang, J.; Qi, Z. Baricitinib, a drug with potential effect to prevent SARS-CoV-2 from entering target cells and control cytokine storm induced by COVID-19. Int. Immunopharmacol. 2020, 86, 106749. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, S.; Yi, D.; Li, Q.; Ma, L.; Zhang, Y.; Wang, J.; Li, X.; Guo, F.; Lin, R.; et al. A cell-based assay to discover inhibitors of SARS-CoV-2 RNA dependent RNA polymerase. Antivir. Res. 2021, 190, 105078. [Google Scholar] [CrossRef]
- Nie, J.; Li, Q.; Wu, J.; Zhao, C.; Hao, H.; Liu, H.; Zhang, L.; Nie, L.; Qin, H.; Wang, M.; et al. Establishment and validation of a pseudovirus neutralization assay for SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 680–686. [Google Scholar] [CrossRef]
- Govinda, K.C.; Giovanni, B.; Srijan, V.; Mahmudulla, H.; Jayme, H.; Jeremy, Y. A machine learning platform to estimate anti-SARS-CoV-2 activities. Nat. Mach. Intell. 2021, 3, 527–535. [Google Scholar]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Xiu, S.; Dick, A.; Ju, H.; Mirzaie, S.; Abdi, F.; Cocklin, S.; Zhan, P.; Liu, X. Inhibitors of SARS-CoV-2 entry: Current and future opportunities. J. Med. Chem. 2020, 63, 12256–12274. [Google Scholar] [CrossRef]
- Lundin, A.; Dijkman, R.; Bergström, T.; Kann, N.; Adamiak, B.; Hannoun, C.; Kindler, E.; Jónsdóttir, H.R.; Muth, D.; Kint, J.; et al. Targeting membrane-bound viral RNA synthesis reveals potent inhibition of diverse coronaviruses including the middle east respiratory syndrome virus. PLoS Pathol. 2014, 10, e1004166. [Google Scholar] [CrossRef]
- Gurevich, E.V.; Gurevich, V.V. Therapeutic potential of small molecules and engineered proteins. In Arrestins-Pharmacology and Therapeutic Potential; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–12. [Google Scholar]
- Ngo, H.X.; Garneau-Tsodikova, S. What are the drugs of the future? MedChemComm 2018, 9, 757–758. [Google Scholar] [CrossRef] [PubMed]
- Milewska, A.; Ciejka, J.; Kaminski, K.; Karewicz, A.; Bielska, D.; Zeglen, S.; Wojciech, K.; Nowakowska, M.; Potempa, J.; Bosch, B.J.; et al. Novel polymeric inhibitors of HCoV-NL63. Antivir. Res. 2013, 97, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Mitsuki, Y.Y.; Ohnishi, K.; Takagi, H.; Oshima, M.; Yamamoto, T.; Mizukoshi, F.; Terahara, K.; Kobayashi, K.; Yamamoto, N.; Yamaoka, S.; et al. A single amino acid substitution in the S1 and S2 Spike protein domains determines the neutralization escape phenotype of SARS-CoV. Microbes Infect. 2008, 10, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Wahedi, H.M.; Ahmad, S.; Abbasi, S.W. Stilbene-based natural compounds as promising drug candidates against COVID-19. J. Biomol. Struct. Dyn. 2020, 39, 3225–3234. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Rivera, O.A.; Shukla, S.; Shin, M.D.; Chen, A.; Beiss, V.; Moreno-Gonzalez, M.A.; Zheng, Y.; Clark, A.E.; Carlin, A.F.; Pokorski, J.K.; et al. Cowpea mosaic virus nanoparticle vaccine candidate displaying peptide epitopes can neutralize the severe acute respiratory syndrome coronavirus. ACS Infect. Dis. 2021, 7, 3096–3110. [Google Scholar] [CrossRef]
- Patrick, R.; Garad, R.; Snell, T.; Enticott, J.; Meadows, G. Australians report climate change as a bigger concern than COVID-19. J. Clim. Chang. Health 2021, 3, 100032. [Google Scholar] [CrossRef]
- Beyer, R.M.; Manica, A.; Mora, C. Shifts in global bat diversity suggest a possible role in climate change in the emergence of SARS-CoV-1 and SARS-CoV-2. Sci. Total Environ. 2021, 767, 145413. [Google Scholar] [CrossRef]
- Galeotti, C.; Bayry, J. Autoimmune and inflammatory diseases following COVID-19. Nat. Rev. Rheumatol. 2020, 16, 413–414. [Google Scholar] [CrossRef]
- Toscano, G.; Palmerini, F.; Ravaglia, S.; Ruiz, L.; Invernizzi, P.; Cuzzoni, M.G.; Baldanti, F.; Daturi, R.; Postorino, P.; Cavallini, A.; et al. Guillain-Barré syndrome associated with SARS-CoV-2. N. Engl. J. Med. 2020, 382, 2574–2576. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, T.; Chen, J.; Hou, C.; Hua, L.; He, S.; Guo, Y.; Zhang, S.; Wang, Y.; Yuan, J.; et al. Clinical and autoimmune characteristics of severe and critical cases of COVID-19. Clin. Transl. Sci. 2020, 13, 1077–1086. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).