

Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells
 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Venom Source and Extraction
2.2. Cell Line and Culture
2.3. Animals and Management Scheme
2.4. Culture Supernatant, Sera and Tumor Lysate
2.5. Antitumoral Effect of H. junceus Scorpion Venom in F3II Bearing BALB/c Mice
2.6. In Vitro Cell Viability Assay (MTT Assay)
2.7. Cytokine Measurement
2.8. Statistical Analysis
3. Results
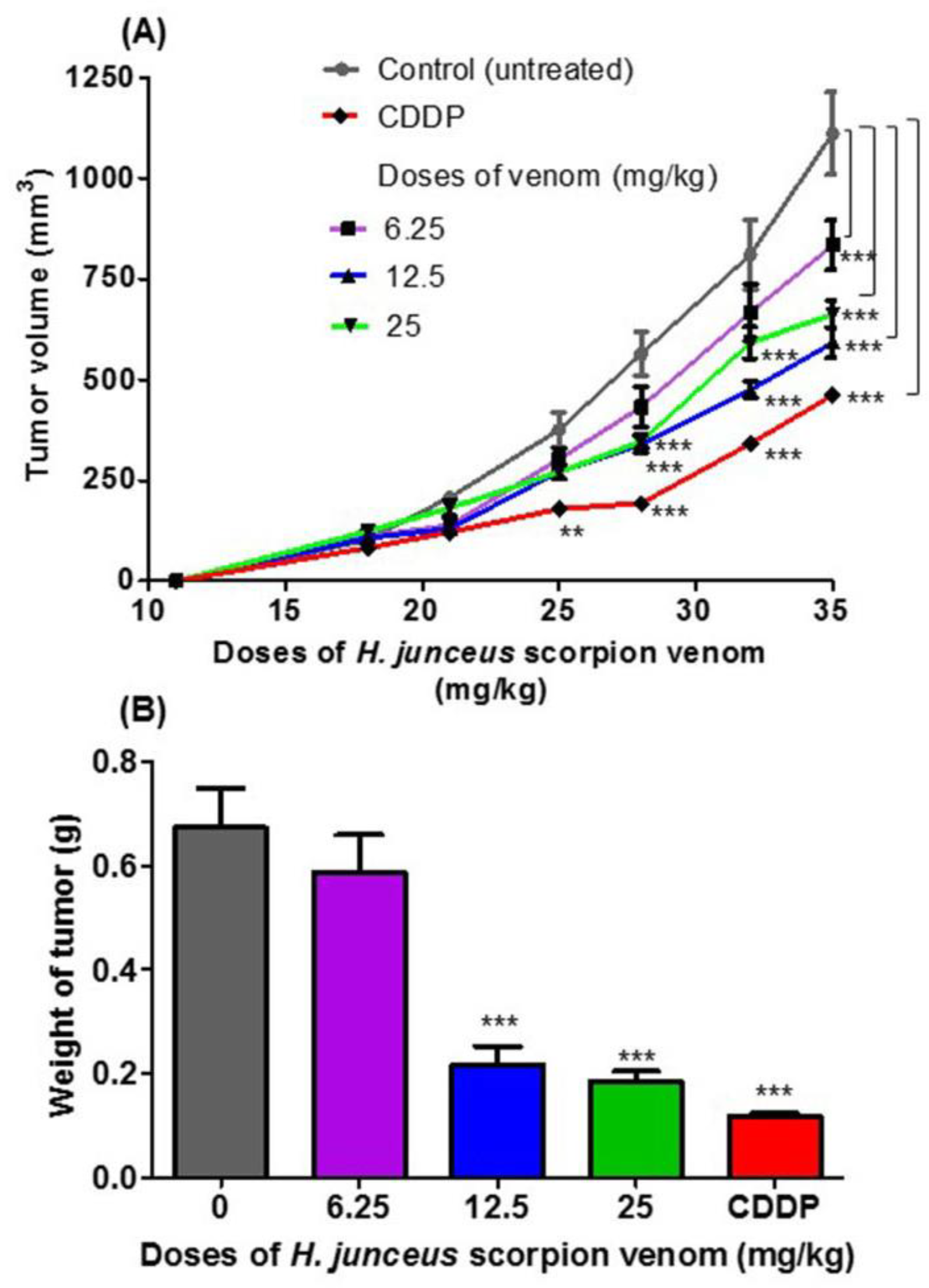
3.1. Antitumoral Effect of H. junceus Scorpion Venom in F3II Bearing-Mice
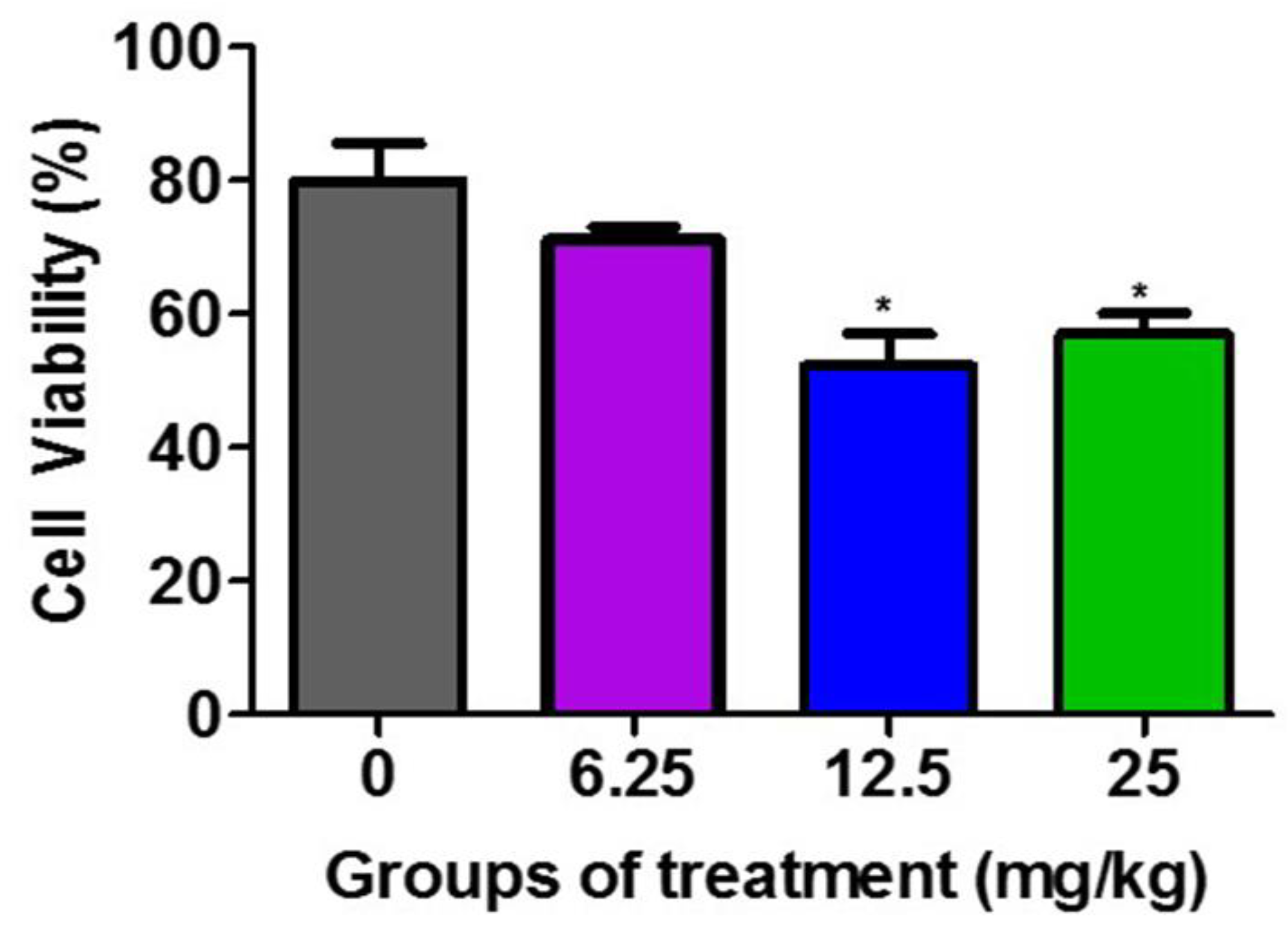
3.2. Effect of Tumor Lysates on the Viability of F3II Tumor Cells
3.3. Cytokine Levels in F3II Supernatant
3.4. Determination of Cytokine Levels in Sera and F3II Tumor Lysates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.; Miller, K.; Sandeep, N.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bazan, M.; Mestre, B.F.; Peix, A. Cardio-Oncology in Cuba: The Present and the Future. JACC Cardio Oncol. 2021, 3, 162–164. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, C.; Saz, A.; Fornaguera, C. Cancer immunotherapies revisited: State of the art of conventional treatments and next-generation nanomedicines. Cancer Gene Ther. 2021, 28, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Najafi, R.; Amini, R.; Solgi, R.; Tanzadehpanah, H.; Esfahani, A.; Saidijam, M. Remarkable apoptotic pathway of Hemiscorpius lepturus scorpion venom on CT26 cell line. Cell Biol. Toxicol. 2019, 35, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.; Pezzuto, J. Natural products as a vital source for the discovery of cancer chemotherapeutic and chemopreventive agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernández, J.; Gutiérrez del Río, I.; Villar, C.; Lombó, F. New Insights toward Colorectal Cancer Chemotherapy Using Natural Bioactive Compounds. Front. Pharmacol. 2017, 8, 109. [Google Scholar] [CrossRef]
- Benarba, B.; Pandiella, A. Colorectal cancer and medicinal plants: Principle findings from recent studies. Biomed. Pharmacother. 2018, 107, 408–423. [Google Scholar] [CrossRef]
- Filin, I.; Solovyeva, V.; Kitaeva, K.; Rutland, C.; Rizvanov, A. Current Trends in Cancer Immunotherapy. Biomedicines 2020, 8, 621. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef]
- Pinto, N.; Carnier, J.; Oyama, L.; Otoch, J.; Alcântara, P.; Tokeshi, F.; Nascimento, C.M. Cancer as a Proinflammatory Environment: Metastasis and Cachexia. Mediators Inflamm. 2015, 2015, 791060. [Google Scholar]
- Esposito, L.; Yamaguti, H.; Souza, C.; Pinto-Da Rocha, R.; Prendini, L. Revision of the neotropical club-tailed scorpions, Physoctonus, Rhopalurus and Troglorhopalurus revalidation of Hete-roctenus, and descriptions of two new genera and three new species (Buthidae: Rhopalurusinae). Bull. Am. Mus. Natural History 2017, 415, 1–136. [Google Scholar] [CrossRef]
- Díaz-García, A.; Morier-Díaz, L.; Frión-Herrera, Y.; Rodríguez-Sánchez, H.; Caballero-Lorenzo, Y.; Mendoza-Llanes, D. In vitro anticancer effect of venom from Cuban scorpion Rhopalurus junceus against a panel of human cancer cell lines. J. Venom. Res. 2013, 4, 5–12. [Google Scholar] [PubMed]
- Casamayor-Laime, Z.; Guevara-García, M.; Gonzalez-Laime, S. Empleo del Vidatox 30 CH en el alivio del dolor y otros síntomas de enfermedades inflamatorias. Rev. Cub Farm. 2020, 53, 1–15. [Google Scholar]
- Díaz-García, A.; Pimentel-González, G.; Basaco-Bernabeu, T.; Rodríguez-Aurrecochea, J.; Rodríguez-Sánchez, H.; Sánchez-Monzón, I. Pharmacokinetics and Biodistribution of Rhopalurus junceus Scorpion Venom in Tumor-Bearing Mice after Intravenous and Oral Administration. Iran. Biomed. J. 2019, 23, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, R.; Tahir, H.; Arshad, M.; Naseem, S.; Ahsan, M. Optimization of the conditions for maximum recovery of venom from scorpions by electrical stimulation. Pakistan J. Zool. 2016, 48, 265–269. [Google Scholar]
- Herrera, Y.; Heras, N.; Cardoso, D. Adaptación a microplacas y validación de la técnica de Lowry. VacciMonitor 1999, 3, 7–11. [Google Scholar]
- García-Gómez, B.; Coronas, F.; Restano-Cassulini, R.; Rodríguez, R.; Possani, L. Biochemical and molecular characterization of the venom from the Cuban scorpion Rhopalurus junceus. Toxicon 2011, 58, 18–27. [Google Scholar] [CrossRef]
- Rodríguez-Ravelo, R.; Batista, C.; Coronas, F.; Zamudio, F.; Hernández-Orihuela, L.; Espinosa-López, G.; Ruiz-Urquiola, A.; Possani, L.D. Comparative proteomic analysis of male and female venoms from the Cuban scorpion Rhopalurus junceus. Toxicon 2015, 107, 327–334. [Google Scholar] [CrossRef]
- Workman, P.; Aboagye, E.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.; Double, J.A.; Everitt, J.; Farningham, D.A.; Glennie, M.J.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1557. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular grow and survival: Application to proliferation and citotoxicity assays. J. Immunol. Meth. 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Allahqoli, L.; Mazidimoradi, A.; Momenimovahed, Z.; Rahmani, A.; Hakimi, S.; Tiznobaik, A.; Gharacheh, M.; Salehiniya, H.; Babaey, F.; Alkatout, I. The Global Incidence, Mortality, and Burden of Breast Cancer in 2019: Correlation with Smoking, Drinking, and Drug Use. Front. Oncol. 2022, 12, 921015. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, D.; Cabezas-Cruz, A.; Mesa, C.; Carmenate, T.; Martínez, D.; Valdés-Zayas, A.; Montero, E.; Pérez, R. Murine Mammary Carcinoma Induces Chronic Systemic Inflammation and Immunosuppression in BALB/c Mice. J. Breast Cancer 2022, 25, 218–232. [Google Scholar] [CrossRef]
- Lagarto, A.; Bueno, V.; Pérez, M.; Rodríguez, C.; Guevara, I.; Valdés, O.; Bellma, A.; Gabilondo, T.; Padrón, A.S. Safety evaluation of the venom from scorpion Rhopalurus junceus: Assessment of oral short term, subchronic toxicity and teratogenic effect. Toxicon 2020, 176, 59–66. [Google Scholar] [CrossRef]
- Díaz-García, A.; Ruiz-Fuentes, J.L.; Frión-Herrera, Y.; Yglesias-Rivera, A.; Riquenez Garlobo, Y.; Rodríguez Sánchez, H.; Rodríguez Aurrecochea, J.C.; López Fuentes, L.X. Rhopalurus junceus scorpion venom induces antitumor effect in vitro and in vivo against a murine mammary adenocarcinoma model. Iran. J. Basic. Med. Sci. 2019, 22, 759–765. [Google Scholar]
- Al Asmari, A.K.; Khan, A.Q. Investigation of in vivo potential of scorpion venom against skin tumorigenesis in mice via targeting markers associated with cancer development. Drug Des. Devel Ther. 2016, 10, 3387–3397. [Google Scholar] [CrossRef]
- Salem, M.L.; Shoukry, N.M.; Teleb, W.K.; Abdel-Daim, M.M.; Abdel-Rahman, M.A. In vitro and in vivo antitumor effects of the Egyptian scorpion Androctonus amoreuxi venom in an Ehrlich ascites tumor model. Springer Plus 2016, 5, 570. [Google Scholar] [CrossRef]
- Habanjar, O.; Bingula, R.; Decombat, C.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. Crosstalk of Inflammatory Cytokines within the Breast Tumor Microenvironment. Int. J. Mol. Sci. 2023, 24, 4002. [Google Scholar] [CrossRef]
- Gajewski, T.; Schreiber, H.; Fu, Y. Innate and adaptive immune cells in the tumour microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and Cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Mikaelian, A.; Traboulay, E.; Zhang, X.; Yeritsyan, E.; Pedersen, P.; Ko, Y.; Matalka, K.Z. Pleiotropic Anticancer Properties of Scorpion Venom Peptides: Rhopalurus princeps Venom as an Anticancer Agent. Drug Des. Devel Ther. 2020, 14, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Place, D.; Kanneganti, T. Cell death–mediated cytokine release and its therapeutic implications. J. Exp. Med. 2019, 216, 1474–1486. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Huang, Y.; Oshima, K.; Yearsley, M.; Zhang, J.; Arnold, M.; Yu, J.; Wang, L.S. The immunomodulatory potential of natural compounds in tumor-bearing mice and humans. Crit. Rev. Food Sci. Nutr. 2019, 59, 992–1007. [Google Scholar] [CrossRef]
- Rébé, C.; Ghiringhelli, F. Interleukin-1β and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef]
- Gocher, A.M.; Workman, C.J.; Vignali, D.A.A. Interferon-γ: Teammate or opponent in the tumour microenvironment? Nat. Rev. Immunol. 2022, 22, 158–172. [Google Scholar] [CrossRef]
- Autenshlyus, A.; Arkhipov, S.; Kunts, T.; Marinkin, I.; Mikhailova, E.; Karpukhina, X.; Varaksin, N.A. Cytokine profiles of tumor supernatants in invasive ductal cancer and fibroadenoma of the breast and its relationship with VEGF-A expression in the tumors. Int. J. Immunopathol. Pharmacol. 2017, 30, 83–88. [Google Scholar] [CrossRef]
- Al-Asmari, A.; Islam, M.; Al-Zahrani, A. In vitro analysis of the anticancer properties of scorpion venom in colorectal and breast cancer cell lines. Oncol. Lett. 2016, 11, 1256–1262. [Google Scholar] [CrossRef]
- Kampo, S.; Ahmmed, B.; Zhou, T.; Owusu, L.; Anabah, T.; Doudou, N.; Kuugbee, E.D.; Cui, Y.; Lu, Z.; Yan, Q.; et al. Scorpion venom analgesic peptide, BmK AGAP inhibits stemness, and epithelial-mesenchymal transition by down-regulating PTX3 in breast cancer. Front. Oncol. 2019, 9, 21. [Google Scholar] [CrossRef]
- He, L.; Kruttika, B.; Duhacheck-Muggy, S.; Ioannidis, A.; Zhang, L.; Nguyen, N.T.; Moatamed, N.A.; Pajonk, F. Tumor necrosis factor receptor signaling modulates carcinogenesis in a mouse model of breast cancer. Neoplasia 2021, 23, 197–209. [Google Scholar] [CrossRef]
- Díaz-García, A. Estudio del Potencial Terapéutico Selectivo y Mecanismos de Acción del Veneno del Escorpión Cubano Rhopalurus junceus Sobre Células Tumorales de Origen Epitelial [Tesis en opción al Título de Doctor en Ciencias Farmacéuticas]. Ph.D. Thesis, University of Havana, La Habana, Cuba, 2015. [Google Scholar]
- Rašková, M.; Lacina, L.; Kejík, Z.; Venhauerová, A.; Skaličková, M.; Kolář, M.; Jakubek, M.; Rosel, D.; Smetana, K.; Brábek, J. The Role of IL-6 in Cancer Cell Invasiveness and Metastasis—Overview and Therapeutic Opportunities. Cells 2022, 11, 3698. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; You, M.; Xu, Y.; Li, F.; Zhang, D.; Li, X. Inhibition of curcumin on myeloid-derived suppressor cells is requisite for controlling lung cancer. Int. Immunopharmacol. 2016, 39, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yglesias-Rivera, A.; Díaz-García, A. Effect of Rhopalurus junceus scorpion venom on inflammation-related cytokines in healthy BALB/c mice. Ann. Microbiol. Immunol. 2018, 1, 1007. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yglesias-Rivera, A.; Sánchez-Rodríguez, H.; Soto-Febles, C.; Monzote, L. Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells. Life 2023, 13, 2287. https://doi.org/10.3390/life13122287
Yglesias-Rivera A, Sánchez-Rodríguez H, Soto-Febles C, Monzote L. Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells. Life. 2023; 13(12):2287. https://doi.org/10.3390/life13122287
Chicago/Turabian StyleYglesias-Rivera, Arianna, Hermis Sánchez-Rodríguez, Carmen Soto-Febles, and Lianet Monzote. 2023. "Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells" Life 13, no. 12: 2287. https://doi.org/10.3390/life13122287
APA StyleYglesias-Rivera, A., Sánchez-Rodríguez, H., Soto-Febles, C., & Monzote, L. (2023). Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells. Life, 13(12), 2287. https://doi.org/10.3390/life13122287