
The Influence of Light Wavelength on Human HPA Axis Rhythms: A Systematic Review



Abstract
:1. Introduction
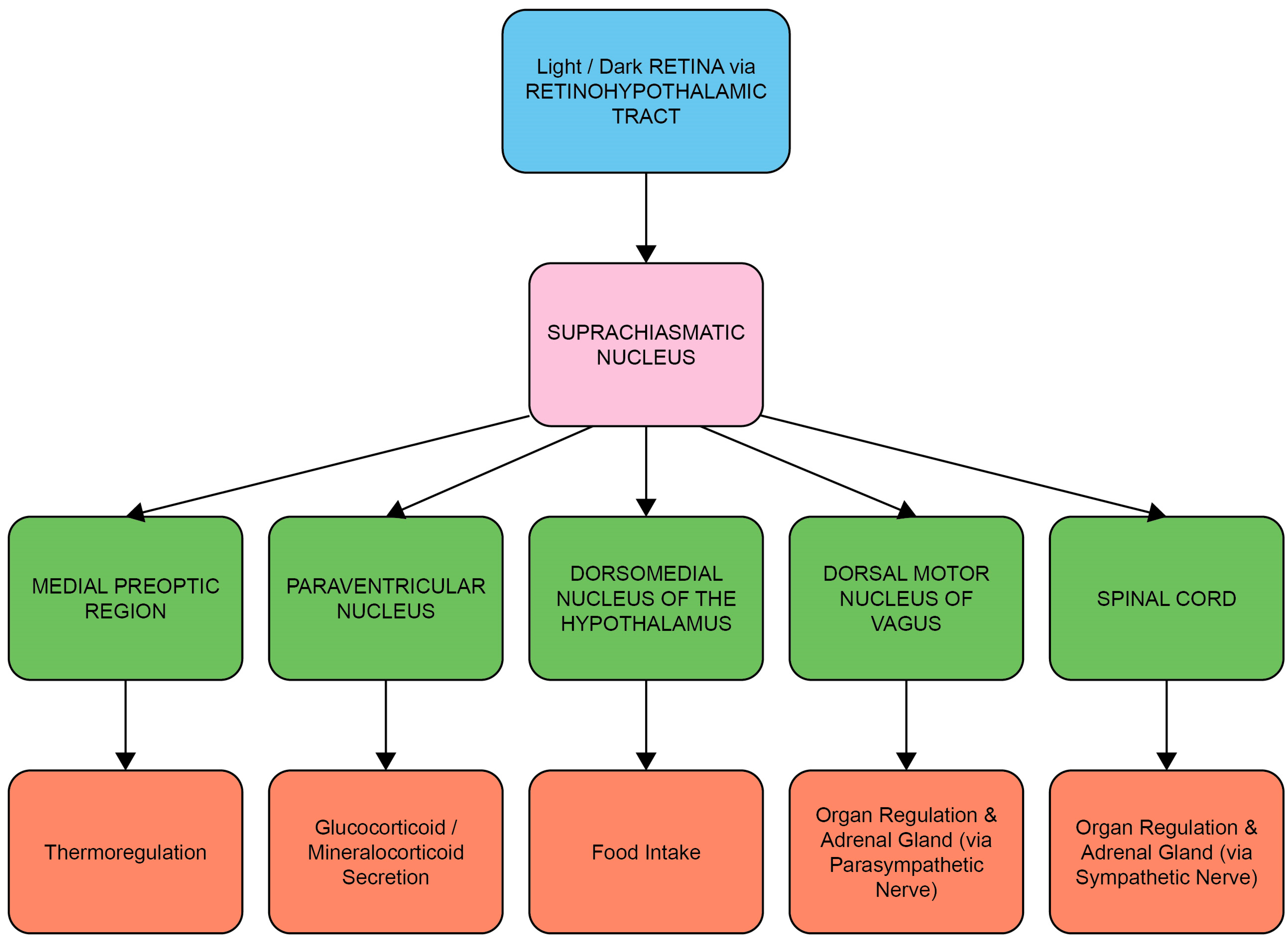
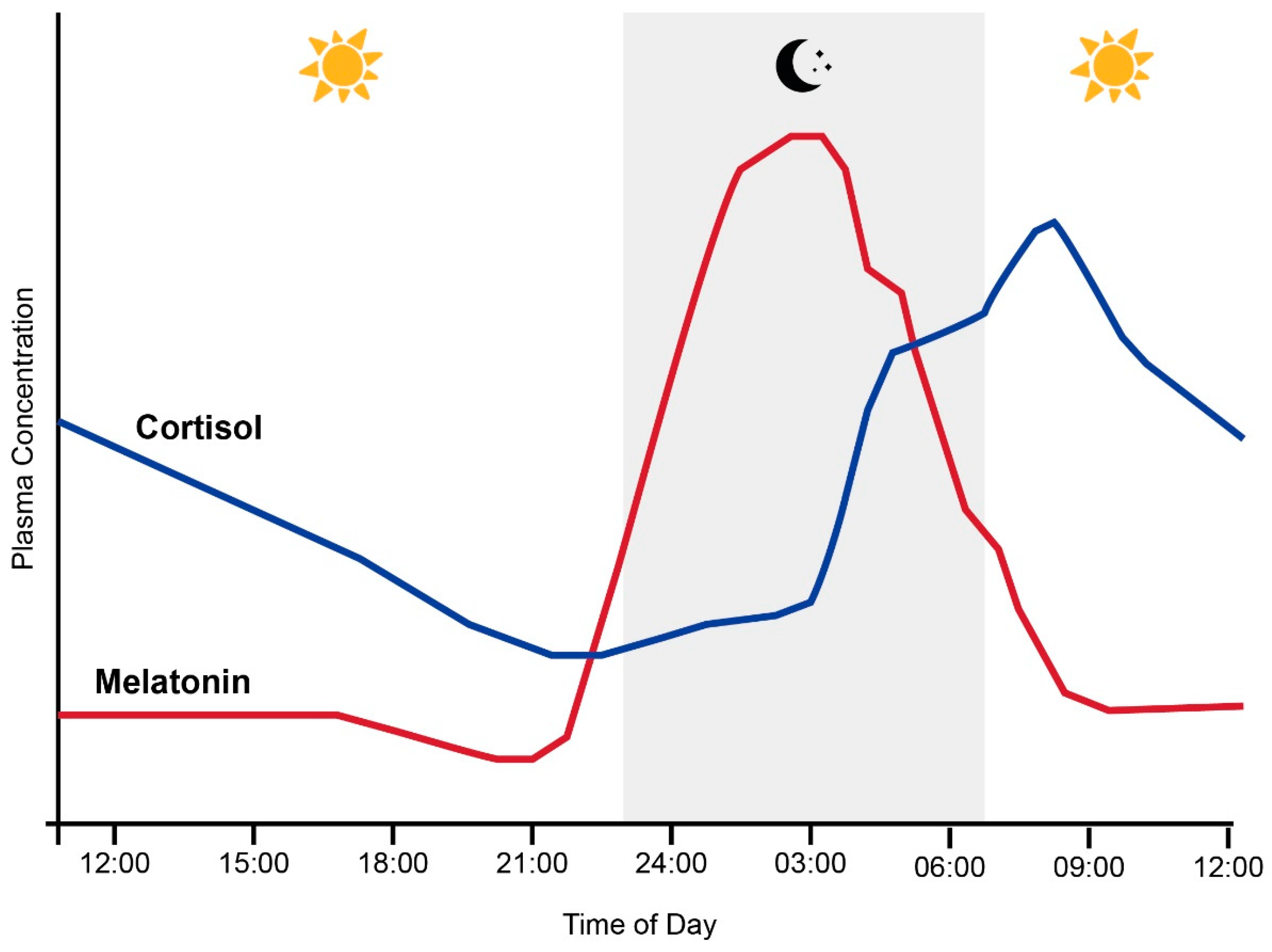
1.1. The Circadian System, Cortisol and Melatonin
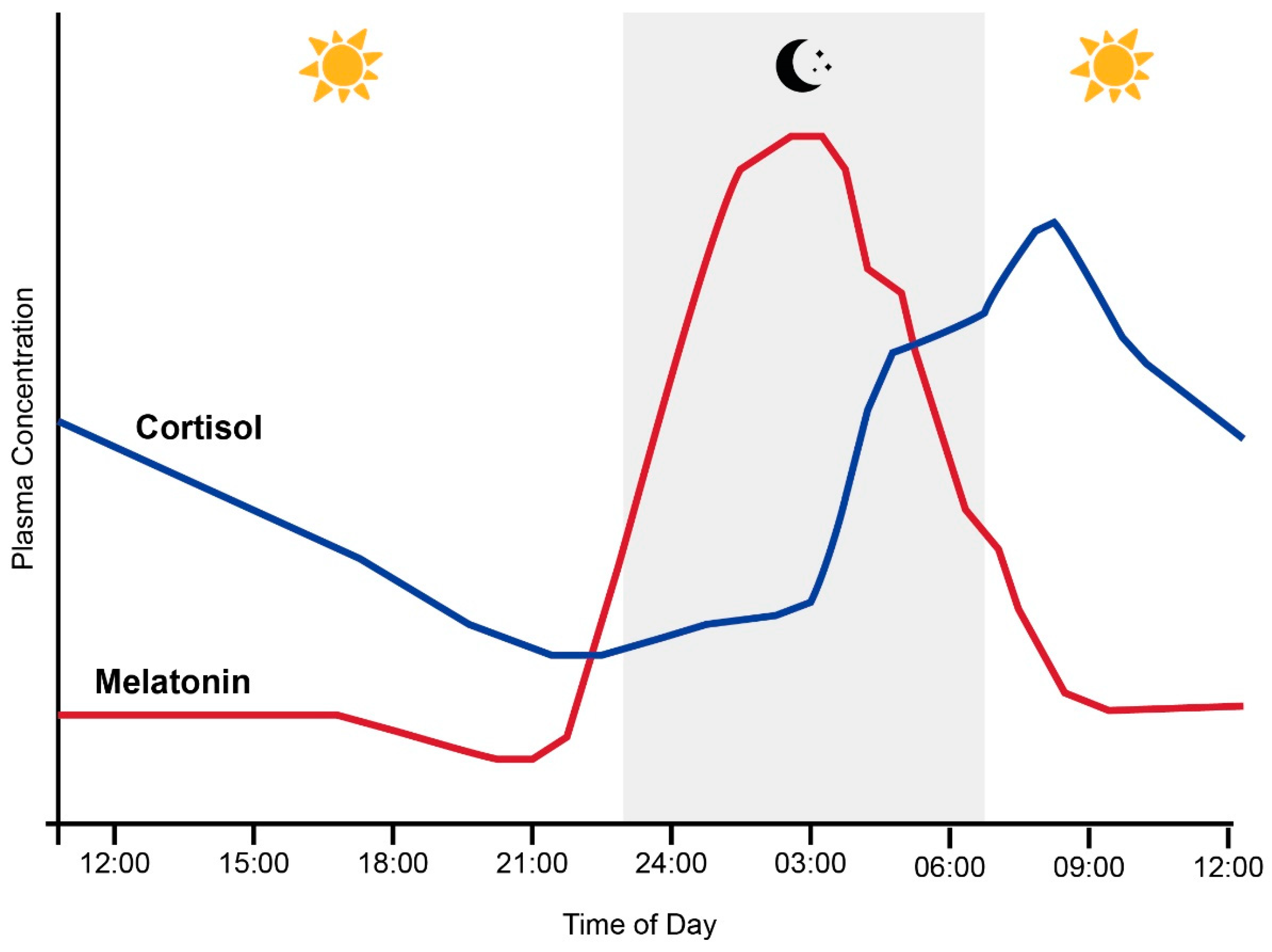
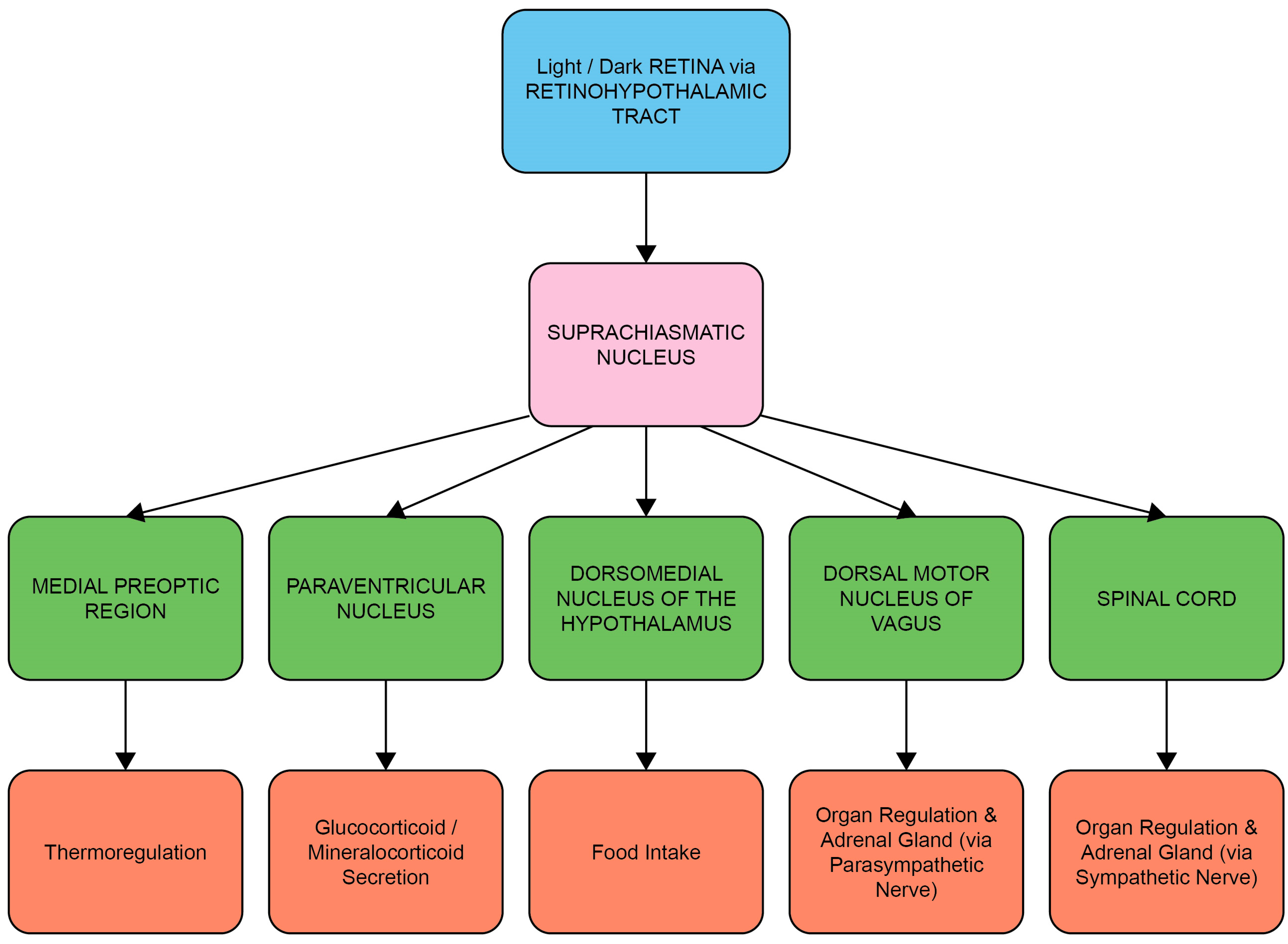
1.2. Entrainment of the Circadian System and HPA Axis by Light
1.3. Systematic Review Aims
2. Methods
2.1. Search Strategy
2.2. Study Selection and Risk of Bias Assessment
2.3. Data Analysis
3. Results
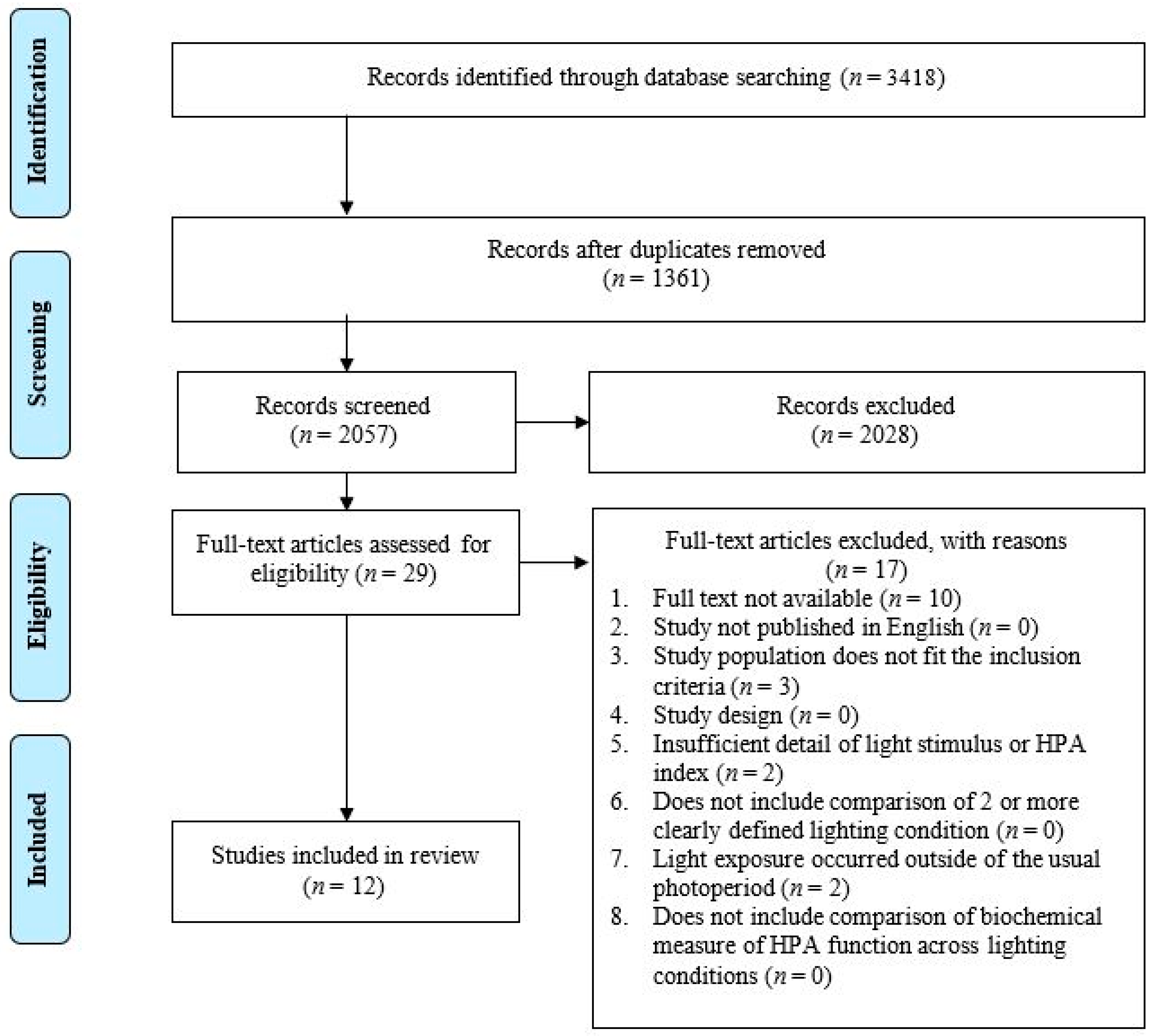
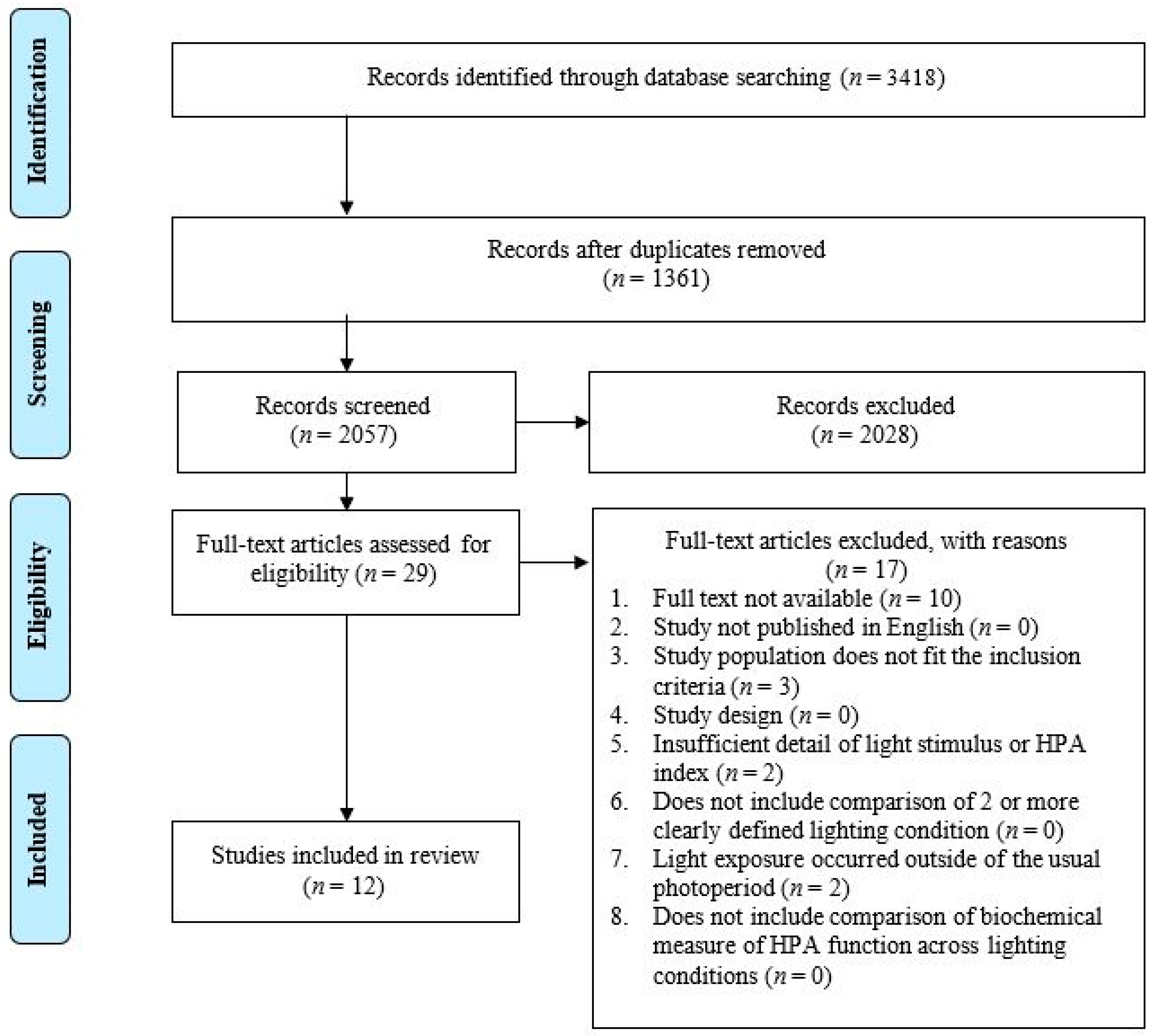
3.1. Study Selection
3.2. Risk of Bias within Included Studies
3.3. Study Characteristics
3.4. Light Exposure Characteristics
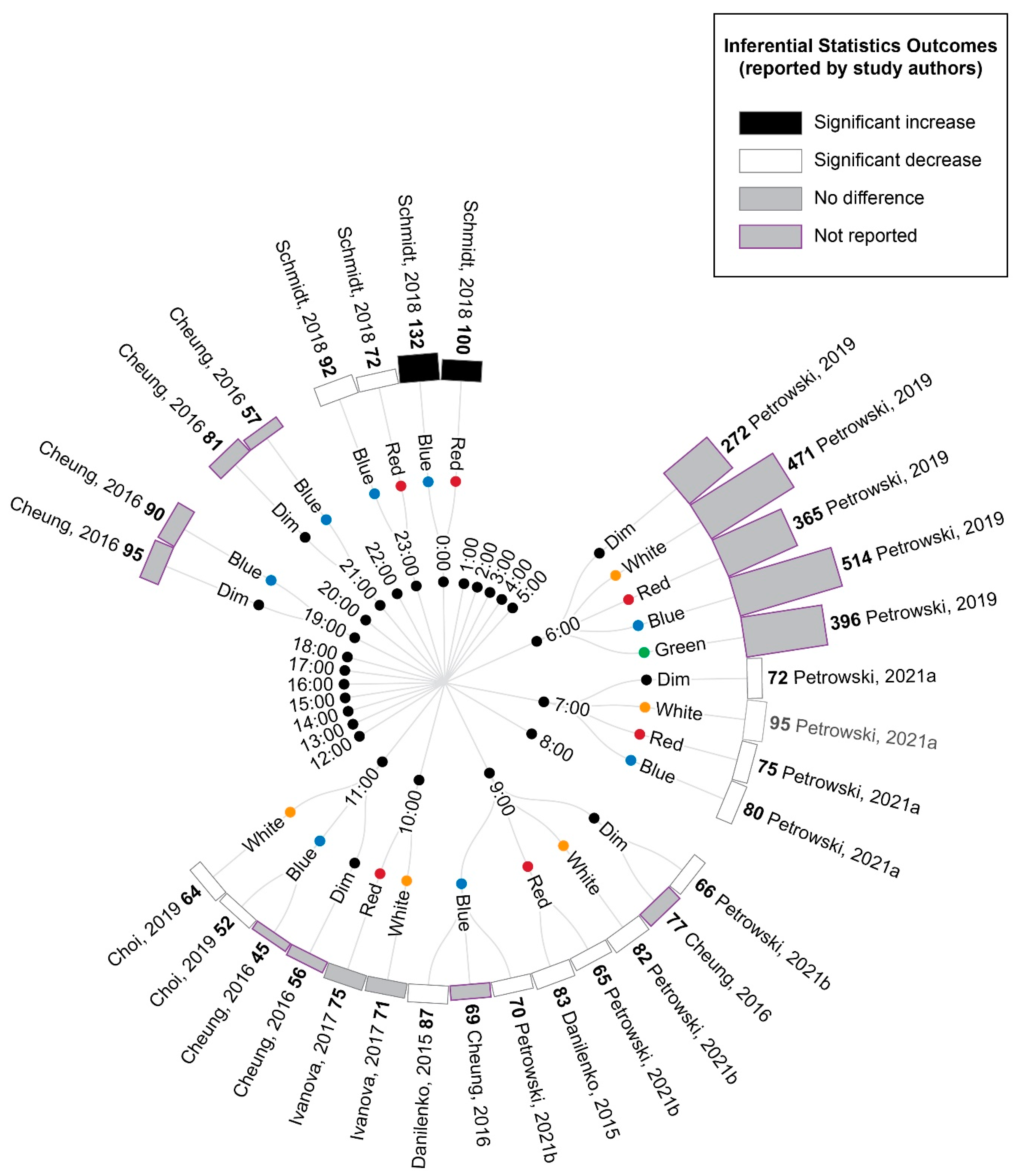
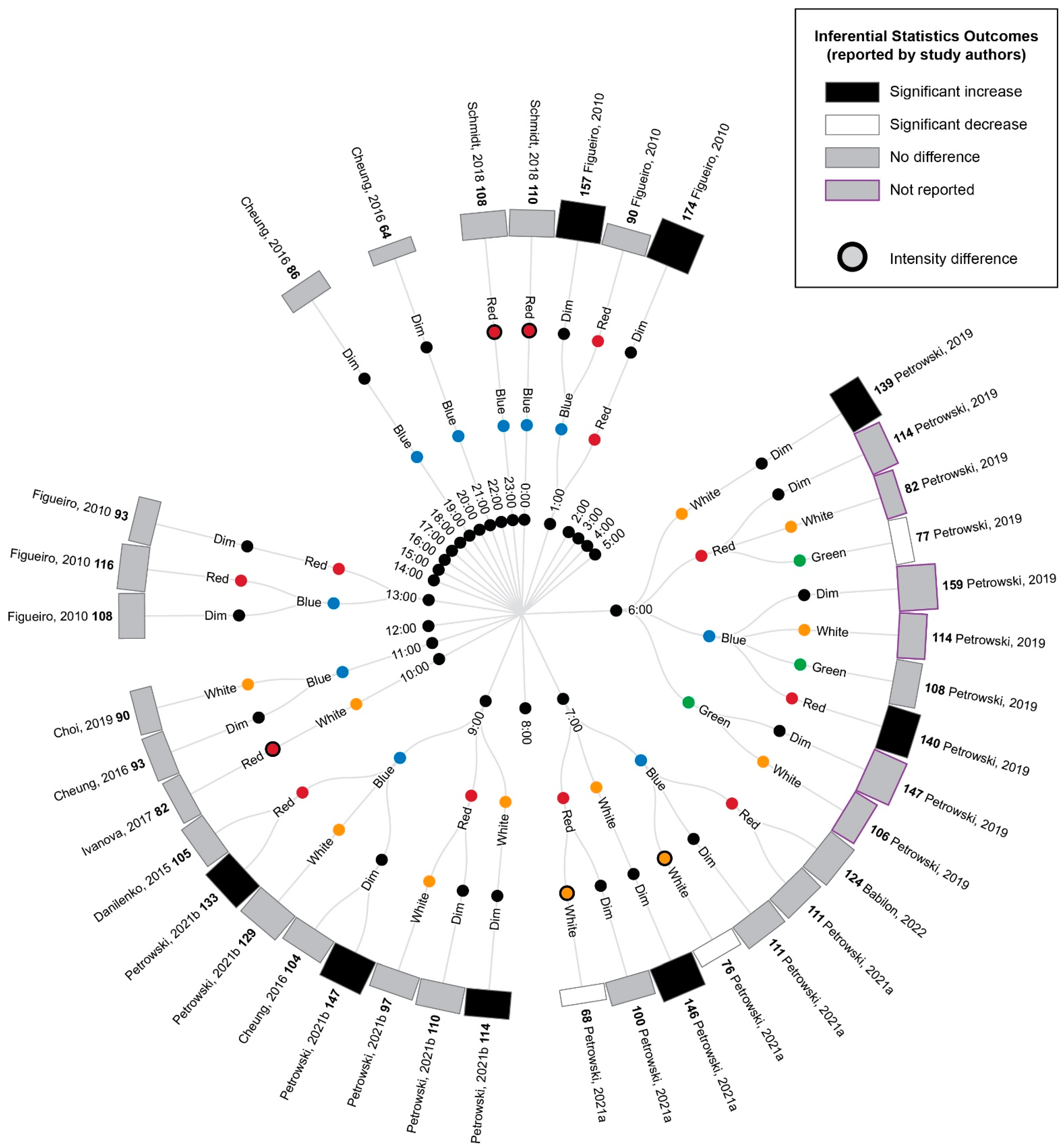
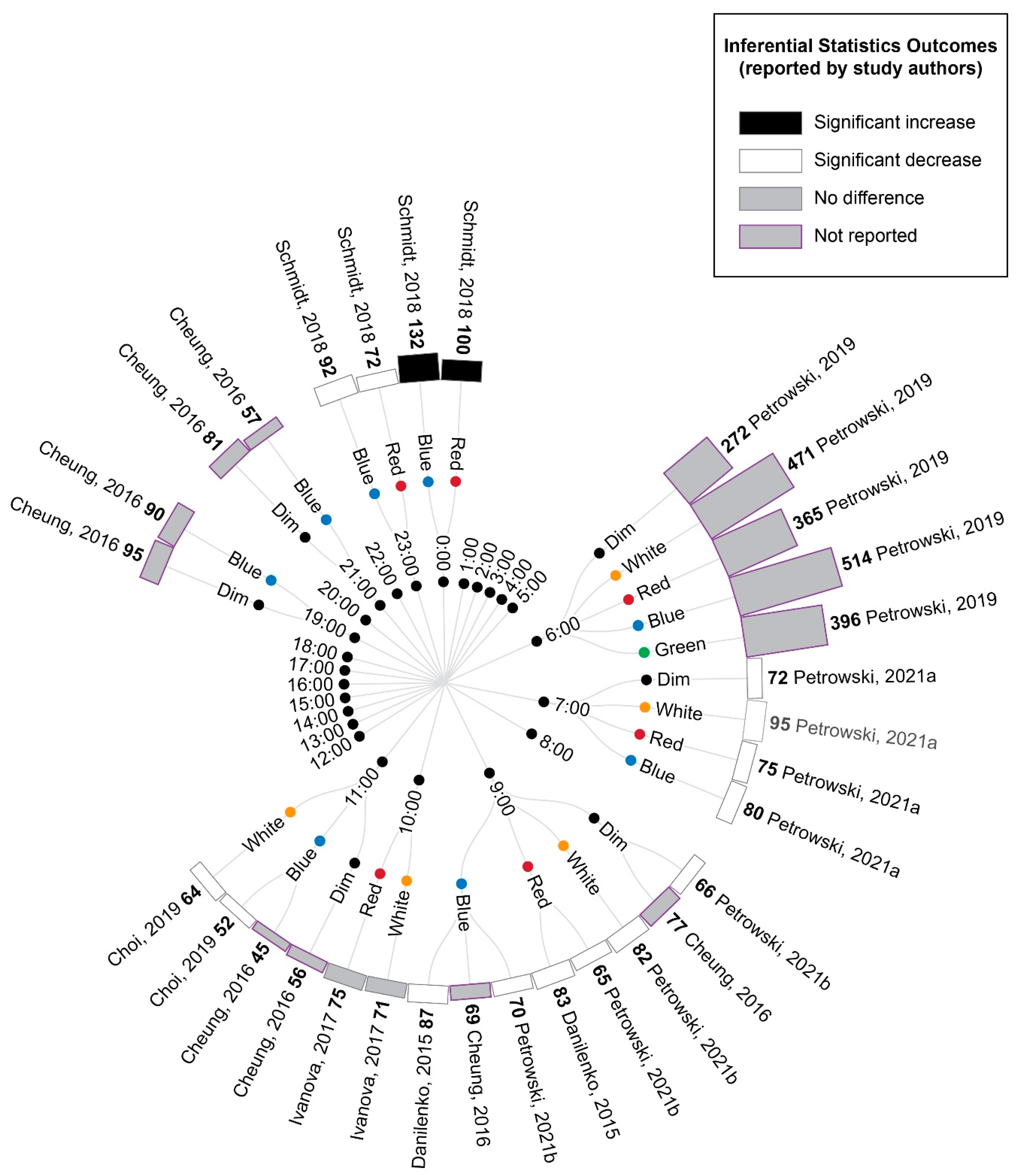
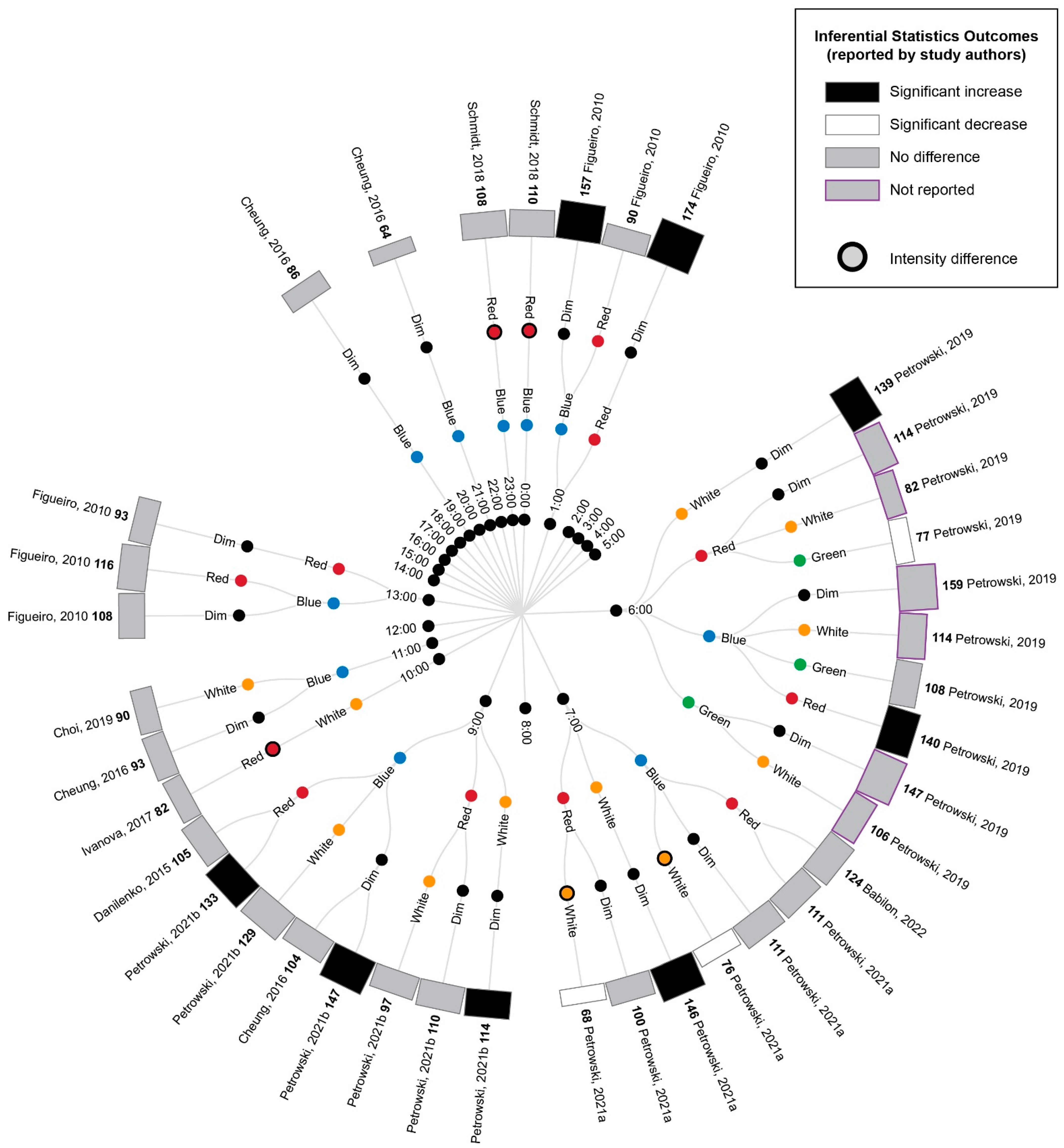
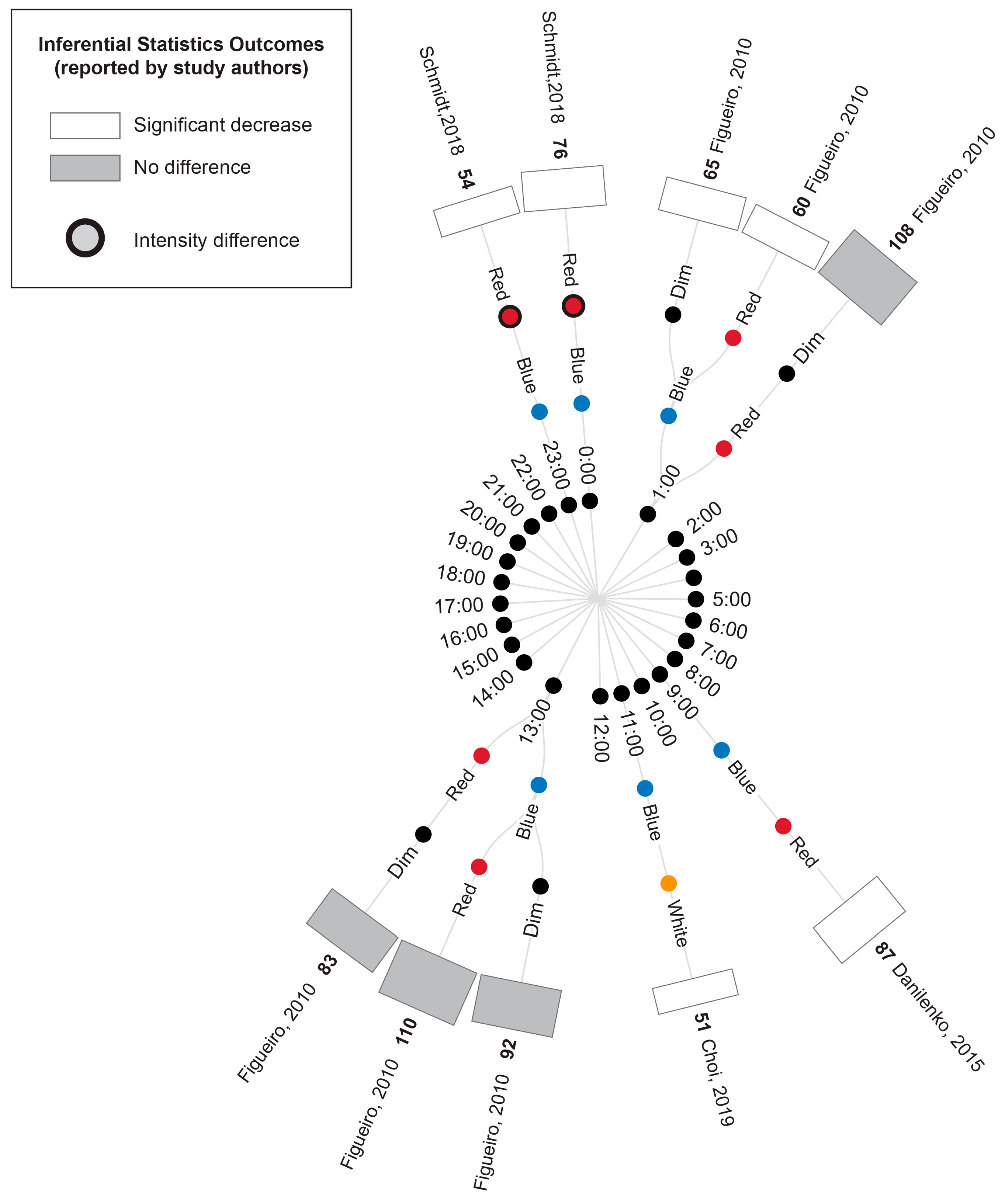
3.5. Study Results
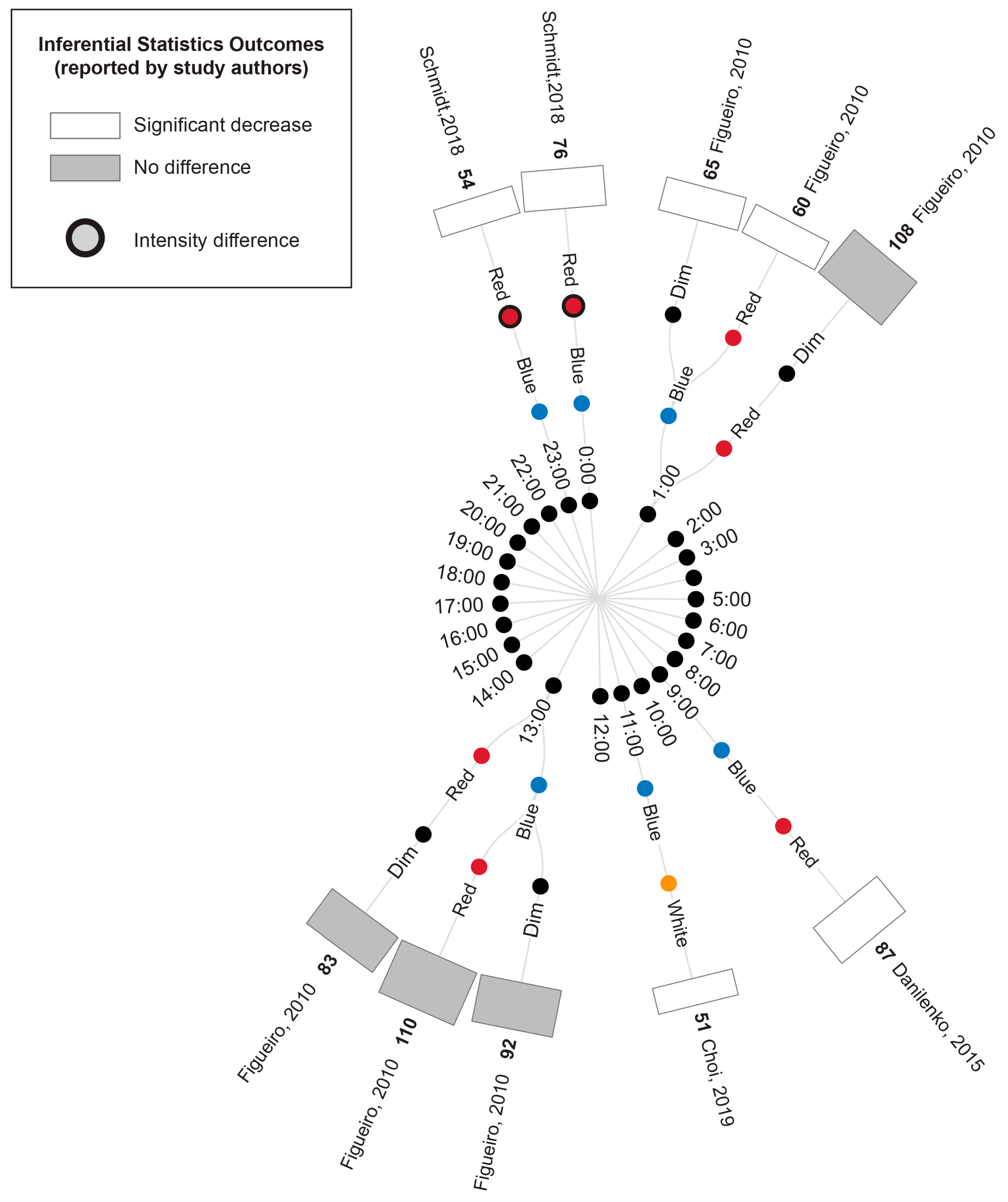
3.5.1. Comparison of Experimental Lighting Conditions with Pre-Exposure Baseline
3.5.2. Comparison of Time-Matched Experimental Lighting Conditions
3.5.3. Concurrent Melatonin Measurements
4. Discussion
4.1. Light Intensity and Wavelength Comparisons
4.2. Limitations and Potential Bias in the Extracted Studies
4.3. Implications and Future Directions for Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brabant, G.; Henley, D.E.; Kaye, J.M.; Lightman, S.L. 2.6.1 Endocrinology, sleep and circadian rhythms. In Oxford Textbook of Endocrinology and Diabetes; Oxford Academic: Oxford, UK, 2011; p. 273. [Google Scholar]
- Androulakis, I.P. Circadian rhythms and the HPA axis: A systems view. WIREs Mech. Dis. 2021, 13, e1518. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Mutoh, T.; Ueyama, T.; Bando, H.; Masubuchi, S.; Nakahara, D.; Tsujimoto, G.; Okamura, H. Light activates the adrenal gland: Timing of gene expression and glucocorticoid release. Cell Metab. 2005, 2, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Buijs, R.M.; Wortel, J.; Van Heerikhuize, J.J.; Feenstra, M.G.; Ter Horst, G.J.; Romijn, H.J.; Kalsbeek, A. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur. J. Neurosci. 1999, 11, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Lyall, L.M.; Wyse, C.A.; Graham, N.; Ferguson, A.; Lyall, D.M.; Cullen, B.; Celis Morales, C.A.; Biello, S.M.; Mackay, D.; Ward, J.; et al. Association of disrupted circadian rhythmicity with mood disorders, subjective wellbeing, and cognitive function: A cross-sectional study of 91 105 participants from the UK Biobank. Lancet Psychiatry 2018, 5, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Ayyar, V.S.; Sukumaran, S. Circadian rhythms: Influence on physiology, pharmacology, and therapeutic interventions. J. Pharmacokinet. Pharmacodyn. 2021, 48, 321–338. [Google Scholar] [CrossRef]
- Auld, F.; Maschauer, E.L.; Morrison, I.; Skene, D.J.; Riha, R.L. Evidence for the efficacy of melatonin in the treatment of primary adult sleep disorders. Sleep Med. Rev. 2017, 34, 10–22. [Google Scholar] [CrossRef]
- Webb, A.R. Considerations for lighting in the built environment: Non-visual effects of light. Energy Build. 2006, 38, 721–727. [Google Scholar] [CrossRef]
- Petrowski, K.; Buehrer, S.; Niedling, M.; Schmalbach, B. The effects of light exposure on the cortisol stress response in human males. Stress 2021, 24, 29–35. [Google Scholar] [CrossRef]
- Babilon, S.; Myland, P.; Klabes, J.; Simon, J.; Khanh, T.Q. Study protocol for measuring the impact of (quasi-)monochromatic light on post-awakening cortisol secretion under controlled laboratory conditions. PLoS ONE 2022, 17, e0267659. [Google Scholar] [CrossRef]
- Bellia, L.; Bisegna, F.; Spada, G. Lighting in indoor environments: Visual and non-visual effects of light sources with different spectral power distributions. Build. Environ. 2011, 46, 1984–1992. [Google Scholar] [CrossRef]
- Do, A.; Li, V.W.; Huang, S.; Michalak, E.E.; Tam, E.M.; Chakrabarty, T.; Yatham, L.N.; Lam, R.W. Blue-Light Therapy for Seasonal and Non-Seasonal Depression: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Can. J. Psychiatry 2022, 67, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, S.M.; Bee, P.E.; Meyer, N.; Dijk, D.J.; Drake, R.J. Light therapies to improve sleep in intrinsic circadian rhythm sleep disorders and neuro-psychiatric illness: A systematic review and meta-analysis. Sleep. Med. Rev. 2019, 46, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Sanz, J.A.; Tapia-Ayuga, C.E. Blue light emission spectra of popular mobile devices: The extent of user protection against melatonin suppression by built-in screen technology and light filtering software systems. Chronobiol. Int. 2020, 37, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Good, P.A.; Taylor, R.H.; Mortimer, M.J. The use of tinted glasses in childhood migraine. Headache 1991, 31, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.H.; Digre, K.B.; Patel, B.C.; Anderson, R.L.; Warner, J.E.; Katz, B.J. The evaluation of light sensitivity in benign essential blepharospasm. Am. J. Ophthalmol. 2006, 142, 82–87. [Google Scholar] [CrossRef]
- Williams, G.J.; Kitchener, G.; Press, L.J.; Scheiman, M.M.; Steele, G.T. The use of tinted lenses and colored overlays for the treatment of dyslexia and other related reading and learning disorders. Optometry 2004, 75, 720–722. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Pfeffer, M.; Korf, H.W.; Wicht, H. Synchronizing effects of melatonin on diurnal and circadian rhythms. Gen. Comp. Endocrinol. 2018, 258, 215–221. [Google Scholar] [CrossRef]
- Rao, R.; Androulakis, I.P. The physiological significance of the circadian dynamics of the HPA axis: Interplay between circadian rhythms, allostasis and stress resilience. Horm. Behav. 2019, 110, 77–89. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Srinivasan, V.; Spence, D.W.; Cardinali, D.P. Role of the Melatonin System in the Control of Sleep. CNS Drugs 2007, 21, 995–1018. [Google Scholar] [CrossRef] [PubMed]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; de Kloet, E.R.; Dijk, D.J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids. Endocr. Rev. 2017, 38, 3–45. [Google Scholar] [CrossRef] [PubMed]
- Clow, A.; Hucklebridge, F.; Thorn, L. The Cortisol Awakening Response in Context. In International Review of Neurobiology; Clow, A., Thorn, L., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 93, pp. 153–175. [Google Scholar]
- Cuesta, M.; Cermakian, N.; Boivin, D.B. Glucocorticoids entrain molecular clock components in human peripheral cells. Faseb. J. 2015, 29, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Montaruli, A.; Castelli, L.; Mulè, A.; Scurati, R.; Esposito, F.; Galasso, L.; Roveda, E. Biological Rhythm and Chronotype: New Perspectives in Health. Biomolecules 2021, 11, 487. [Google Scholar] [CrossRef]
- Hickie, I.B.; Naismith, S.L.; Robillard, R.; Scott, E.M.; Hermens, D.F. Manipulating the sleep-wake cycle and circadian rhythms to improve clinical management of major depression. BMC Med. 2013, 11, 79. [Google Scholar] [CrossRef]
- Tähkämö, L.; Partonen, T.; Pesonen, A.-K. Systematic review of light exposure impact on human circadian rhythm. Chronobiol. Int. 2019, 36, 151–170. [Google Scholar] [CrossRef]
- Vondrašová, D.; Hájek, I.; Illnerová, H. Exposure to long summer days affects the human melatonin and cortisol rhythms. Brain Res. 1997, 759, 166–170. [Google Scholar] [CrossRef]
- Hofman, M.A.; Swaab, D.F. Diurnal and Seasonal Rhythms of Neuronal Activity in the Suprachiasmatic Nucleus of Humans. J. Biol. Rhythm. 1993, 8, 283–295. [Google Scholar] [CrossRef]
- Münch, M.; Bromundt, V. Light and chronobiology: Implications for health and disease. Dialogues Clin. Neurosci. 2012, 14, 448–453. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.J.; Kronauer, R.; Brown, E.; Czeisler, C. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526 Pt 3, 695–702. [Google Scholar] [CrossRef]
- Phillips, A.J.K.; Vidafar, P.; Burns, A.C.; McGlashan, E.M.; Anderson, C.; Rajaratnam, S.M.W.; Lockley, S.W.; Cain, S.W. High sensitivity and interindividual variability in the response of the human circadian system to evening light. Proc. Natl. Acad. Sci. USA 2019, 116, 12019–12024. [Google Scholar] [CrossRef] [PubMed]
- Sliney, D.H. What is light? The visible spectrum and beyond. Eye 2016, 30, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.M.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action spectrum for melatonin regulation in humans: Evidence for a novel circadian photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Jud, C.; Münch, M.; Kobialka, S.; Wirz-Justice, A.; Albrecht, U. Evening exposure to blue light stimulates the expression of the clock gene PER2 in humans. Eur. J. Neurosci. 2006, 23, 1082–1086. [Google Scholar] [CrossRef]
- Gooley, J.J.; Rajaratnam, S.M.; Brainard, G.C.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Spectral responses of the human circadian system depend on the irradiance and duration of exposure to light. Sci. Transl. Med. 2010, 2, 31ra33. [Google Scholar] [CrossRef]
- Rahman, S.A.; Brainard, G.C.; Czeisler, C.A.; Lockley, S.W. Spectral sensitivity of circadian phase resetting, melatonin suppression and acute alerting effects of intermittent light exposure. Biochem. Pharmacol. 2021, 191, 114504. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Cai, H.; Li, X. Non-visual effects of indoor light environment on humans: A review(✰). Physiol. Behav. 2021, 228, 113195. [Google Scholar] [CrossRef]
- Cai, J.; Hao, W.; Zeng, S.; Qu, X.; Guo, Y.; Tang, S.; An, X.; Luo, A. Effects of Red Light on Circadian Rhythm: A Comparison Among Lamps With Similar Correlated Color Temperatures Yet Distinct Spectrums. IEEE Access 2021, 9, 59222–59230. [Google Scholar] [CrossRef]
- Petrowski, K.; Schmalbach, B.; Niedling, M.; Stalder, T. The effects of post-awakening light exposure on the cortisol awakening response in healthy male individuals. Psychoneuroendocrinology 2019, 108, 28–34. [Google Scholar] [CrossRef]
- Petrowski, K.; Bührer, S.; Albus, C.; Schmalbach, B. Increase in cortisol concentration due to standardized bright and blue light exposure on saliva cortisol in the morning following sleep laboratory. Stress 2021, 24, 331–337. [Google Scholar] [CrossRef]
- Choi, K.; Shin, C.; Kim, T.; Chung, H.J.; Suk, H.-J. Awakening effects of blue-enriched morning light exposure on university students’ physiological and subjective responses. Sci. Rep. 2019, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, K.V.; Sergeeva, O.Y. Immediate effect of blue-enhanced light on reproductive hormones in women. Neuro Endocrinol. Lett. 2015, 36, 84–90. [Google Scholar] [PubMed]
- Figueiro, M.G.; Rea, M.S. The Effects of Red and Blue Lights on Circadian Variations in Cortisol, Alpha Amylase, and Melatonin. Int. J. Endocrinol. 2010, 2010, 829351. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Xhrouet, M.; Hamacher, M.; Delloye, E.; LeGoff, C.; Cavalier, E.; Collette, F.; Vandewalle, G. Light exposure via a head-mounted device suppresses melatonin and improves vigilant attention without affecting cortisol and comfort. Psych. J. 2018, 7, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.A.; Subnis, U.; Carlson, L.E.; Garland, S.N.; Santos-Iglesias, P.; Piedalue, K.-A.L.; Deleemans, J.M.; Campbell, T.S. Effects of a light therapy intervention on diurnal salivary cortisol in fatigued cancer survivors: A secondary analysis of a randomized controlled trial. J. Psychosom. Res. 2020, 139, 110266. [Google Scholar] [CrossRef]
- Ancoli-Israel, S.; Rissling, M.; Neikrug, A.; Trofimenko, V.; Natarajan, L.; Parker, B.A.; Lawton, S.; Desan, P.; Liu, L. Light treatment prevents fatigue in women undergoing chemotherapy for breast cancer. Support. Care Cancer 2012, 20, 1211–1219. [Google Scholar] [CrossRef]
- Bower, J.E.; Ganz, P.A.; Dickerson, S.S.; Petersen, L.; Aziz, N.; Fahey, J.L. Diurnal cortisol rhythm and fatigue in breast cancer survivors. Psychoneuroendocrinology 2005, 30, 92–100. [Google Scholar] [CrossRef]
- Agustini, B.; Bocharova, M.; Walker, A.J.; Berk, M.; Young, A.H.; Juruena, M.F. Has the sun set for seasonal affective disorder and HPA axis studies? A systematic review and future prospects. J. Affect. Disord. 2019, 256, 584–593. [Google Scholar] [CrossRef]
- Buckley, T.M.; Schatzberg, A.F. On the interactions of the hypothalamic-pituitary-adrenal (HPA) axis and sleep: Normal HPA axis activity and circadian rhythm, exemplary sleep disorders. J. Clin. Endocrinol. Metab. 2005, 90, 3106–3114. [Google Scholar] [CrossRef]
- Nandam, L.S.; Brazel, M.; Zhou, M.; Jhaveri, D.J. Cortisol and Major Depressive Disorder-Translating Findings From Humans to Animal Models and Back. Front. Psychiatry 2019, 10, 974. [Google Scholar] [CrossRef]
- van Maanen, A.; Meijer, A.M.; van der Heijden, K.B.; Oort, F.J. The effects of light therapy on sleep problems: A systematic review and meta-analysis. Sleep Med. Rev. 2016, 29, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef] [PubMed]
- Bramer, W.M.; Rethlefsen, M.L.; Kleijnen, J.; Franco, O.H. Optimal database combinations for literature searches in systematic reviews: A prospective exploratory study. Syst. Rev. 2017, 6, 245. [Google Scholar] [CrossRef]
- Scheuermaier, K.; Laffan, A.M.; Duffy, J.F. Light Exposure Patterns in Healthy Older and Young Adults. J. Biol. Rhythm. 2010, 25, 113–122. [Google Scholar] [CrossRef]
- Ivanova, I.A.; Danilenko, K.V.; Aftanas, L.I. Investigation of an Immediate Effect of Bright Light on Oxygen Consumption, Heart Rate, Cortisol, and α-Amylase in Seasonal Affective Disorder Subjects and Healthy Controls. Neuropsychobiology 2016, 74, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M. Engauge Digitizer Software. Available online: http://markummitchell.github.io/engauge-digitizer (accessed on 21 December 2022).
- Houston, D.C.R. Flourish. Available online: https://flourish.studio/ (accessed on 21 December 2022).
- Petrowski, K.; Schmalbach, B.; Stalder, T. The effects of light exposure on the cortisol awakening response in healthy individuals. Psychoneuroendocrinology 2019, 107, 61. [Google Scholar] [CrossRef]
- Traut, J.; Prius Mengua, J.; Meijer, E.J.; McKillop, L.E.; Alfonsa, H.; Hoerder-Suabedissen, A.; Song, S.M.; Molnár, Z.; Akerman, C.J.; Vyazovskiy, V.V.; et al. Oral Abstract. J. Sleep Res. 2022, 31. [Google Scholar] [CrossRef]
- Partonen, T.; Meesters, A.N.; Maukonen, M.; Männistö, S.; Gordijn, M.C.; Meesters, Y.; Danilenko, K.V.; Ivanova, I.A.; Aftanas, L.I.; Gbyl, K.; et al. Abstracts. Neuropsychobiology 2016, 74, 226–260. [Google Scholar] [CrossRef]
- Kashyap, K.C.; Nissen, L.; Smith, S.S.; Kyle, G. Abstracts. J. Sleep Res. 2012, 21, 1–371. [Google Scholar] [CrossRef]
- Petrowski, K.; Schmallbach, B.; Stalder, T. The effects of light exposure on the cortisol awakening response. Psychoneuroendocrinology 2019, 100, S13. [Google Scholar] [CrossRef]
- Garefelt, J.; Akerstedt, T.; Westerlund, H.; Hanson, L.M.; Sverke, M.; Kecklund, G. Abstracts. J. Sleep Res. 2014, 23, 1–331. [Google Scholar] [CrossRef]
- Rahman, S.; Marcu, S.; Shapiro, C.; Brown, T.; Casper, R. Attenuating Nocturnal Light Induced Disruption in Endocrine, Genetic and Behavioral Circadian Rhythm Phase Markers by Filtering Short Wavelengths. In Proceedings of the 23rd Annual Meeting of the Associated-Professional-Sleep-Societies, Seattle, WA, USA, 6–11 June 2009; p. A46. [Google Scholar]
- Takemura, Y.; Kido, K.; Kawana, H.; Yamamoto, T.; Sanuki, T.; Mukai, Y. Effects of Green Color Exposure on Stress, Anxiety, and Pain during Peripheral Intravenous Cannulation in Dental Patients Requiring Sedation. Int. J. Environ. Res. Public Health 2021, 18, 5939. [Google Scholar] [CrossRef] [PubMed]
- Ba-Ali, S.; Brøndsted, A.E.; Andersen, H.U.; Sander, B.; Jennum, P.J.; Lund-Andersen, H. Assessment of diurnal melatonin, cortisol, activity, and sleep−wake cycle in patients with and without diabetic retinopathy. Sleep Med. 2019, 54, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Gabel, V.; Maire, M.; Reichert, C.F.; Chellappa, S.L.; Schmidt, C.; Hommes, V.; Viola, A.U.; Cajochen, C. Effects of Artificial Dawn and Morning Blue Light on Daytime Cognitive Performance, Well-being, Cortisol and Melatonin Levels. Chronobiol. Int. 2013, 30, 988–997. [Google Scholar] [CrossRef]
- Komada, Y.; Aoki, K.; Gohshi, S.; Ichioka, H.; Shibata, S. Effects of television luminance and wavelength at habitual bedtime on melatonin and cortisol secretion in humans. Sleep Biol. Rhythm. 2015, 13, 316–322. [Google Scholar] [CrossRef]
- Schmid, S.R.; Höhn, C.; Bothe, K.; Plamberger, C.P.; Angerer, M.; Pletzer, B.; Hoedlmoser, K. How Smart Is It to Go to Bed with the Phone? The Impact of Short-Wavelength Light and Affective States on Sleep and Circadian Rhythms. Clocks Sleep 2021, 3, 558–580. [Google Scholar] [CrossRef]
- Makateb, A.; Rashidinia, A.; Khosravifard, K.; Dabaghi, P. Investigating the effects of a blue-blocking software on the daily rhythm of sleep, melatonin, cortisol, positive and negative emotions. Chronobiol. Int. 2023, 40, 896–902. [Google Scholar] [CrossRef]
- Cheung, I.N.; Zee, P.C.; Shalman, D.; Malkani, R.G.; Kang, J.; Reid, K.J. Morning and Evening Blue-Enriched Light Exposure Alters Metabolic Function in Normal Weight Adults. PLoS ONE 2016, 11, e0155601. [Google Scholar] [CrossRef]
- Sahin, L.; Wood, B.M.; Plitnick, B.; Figueiro, M.G. Daytime light exposure: Effects on biomarkers, measures of alertness, and performance. Behav. Brain Res. 2014, 274, 176–185. [Google Scholar] [CrossRef]
- Wahl, S.; Engelhardt, M.; Schaupp, P.; Lappe, C.; Ivanov, I.V. The inner clock—Blue light sets the human rhythm. J. Biophotonics 2019, 12, e201900102. [Google Scholar] [CrossRef]
- Levitan, R.D. What is the optimal implementation of bright light therapy for seasonal affective disorder (SAD)? J. Psychiatry Neurosci. 2005, 30, 72. [Google Scholar] [PubMed]
- Milosavljevic, N.; Cehajic-Kapetanovic, J.; Procyk, C.A.; Lucas, R.J. Chemogenetic Activation of Melanopsin Retinal Ganglion Cells Induces Signatures of Arousal and/or Anxiety in Mice. Curr. Biol. 2016, 26, 2358–2363. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Cain, S.W.; Chang, A.-M.; Phillips, A.J.K.; Münch, M.Y.; Gronfier, C.; Wyatt, J.K.; Dijk, D.-J.; Wright, K.P.; Czeisler, C.A. Sex difference in the near-24-hour intrinsic period of the human circadian timing system. Proc. Natl. Acad. Sci. USA 2011, 108, 15602–15608. [Google Scholar] [CrossRef]
- Horrocks, P.M.; Jones, A.F.; Ratcliffe, W.A.; Holder, G.; White, A.; Holder, R.; Ratcliffe, J.G.; London, D.R. Patterns of ACTH and cortisol pulsatility over twenty-four hours in normal males and females. Clin. Endocrinol. 1990, 32, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gunn, P.J.; Middleton, B.; Davies, S.K.; Revell, V.L.; Skene, D.J. Sex differences in the circadian profiles of melatonin and cortisol in plasma and urine matrices under constant routine conditions. Chronobiol. Int. 2016, 33, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Torii, H.; Mori, K.; Okano, T.; Kondo, S.; Yang, H.Y.; Yotsukura, E.; Hanyuda, A.; Ogawa, M.; Negishi, K.; Kurihara, T.; et al. Short-Term Exposure to Violet Light Emitted from Eyeglass Frames in Myopic Children: A Randomized Pilot Clinical Trial. J. Clin. Med. 2022, 11, 6000. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Dhakal, R.; Verkicharla, P.K. Short-Term Exposure to Blue Light Shows an Inhibitory Effect on Axial Elongation in Human Eyes Independent of Defocus. Investig. Ophthalmol. Vis. Sci. 2021, 62, 22. [Google Scholar] [CrossRef]
- Cherian, K.; Schatzberg, A.F.; Keller, J. HPA axis in psychotic major depression and schizophrenia spectrum disorders: Cortisol, clinical symptomatology, and cognition. Schizophr. Res. 2019, 213, 72–79. [Google Scholar] [CrossRef]
- Hu, H.; Kang, C.; Hou, X.; Zhang, Q.; Meng, Q.; Jiang, J.; Hao, W. Blue Light Deprivation Produces Depression-Like Responses in Mongolian Gerbils. Front. Psychiatry 2020, 11, 233. [Google Scholar] [CrossRef]
- Keller, J.; Flores, B.; Gomez, R.G.; Solvason, H.B.; Kenna, H.; Williams, G.H.; Schatzberg, A.F. Cortisol circadian rhythm alterations in psychotic major depression. Biol. Psychiatry 2006, 60, 275–281. [Google Scholar] [CrossRef]
- Germain, A.; Kupfer, D.J. Circadian rhythm disturbances in depression. Hum. Psychopharmacol. 2008, 23, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Adam, E.K.; Vrshek-Schallhorn, S.; Kendall, A.D.; Mineka, S.; Zinbarg, R.E.; Craske, M.G. Prospective associations between the cortisol awakening response and first onsets of anxiety disorders over a six-year follow-up--2013 Curt Richter Award Winner. Psychoneuroendocrinology 2014, 44, 47–59. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Search Terms | Search Results |
|---|---|
| “Light intensity” OR “light exposure” OR “artificial light” OR wavelength OR “violet light” OR “blue light” OR “green light” OR “yellow light” OR “red light” OR “light spectrum”. | 155,092 |
| HPA OR “HPA axis” OR “HPA activity” OR “HPA function” OR “hypothalamic pituitary adrenal” OR CRH OR “corticotropin releasing hormone” OR CRF OR “corticotropin releasing factor” OR ACTH OR “adrenocorticotropic hormone” OR CORT OR corticosterone OR cortisol OR glucocorticoid. | 430,111 |
| 1 AND 2 | 597 |
| Pre-Specified Hierarchy of Exclusion Criteria | |
|---|---|
| 1. | Full text not available (e.g., conference abstract). |
| 2. | Study not published in English. |
| 3. | Study population did not meet the inclusion criteria of healthy adult human, postnatal, without any known pre-existing disease (e.g., cancer, sleep deprivation). |
| 4. | Study design was not observational (longitudinal, cross-sectional, book, qualitative, etc.). |
| 5. | Insufficient light stimulus or HPA index detail. |
| 6. | Does not include comparison of two or more clearly defined lighting conditions (e.g., light wavelength information not provided). |
| 7. | Light exposure occurred only outside of the usual daylight photoperiod, as defined by the study authors (or 5:00–24:00, 24 h time, if not defined [56]). |
| 8. | Does not include comparison of biochemical measures of HPA function across lighting conditions (i.e., corticosterone, cortisol, corticotropin releasing hormone and glucocorticoid). |
| OHAT Risk of Bias Questions | |
|---|---|
| 1. | Was administration dose or exposure level adequately randomized? |
| 2. | Was allocation to study adequately concealed? |
| 3. | Were the research personnel and human subjects blinded to the study group during the study? |
| 4. | Were outcome data complete without attrition or exclusion from analysis? |
| 5. | Can we be confident in the exposure characterization? |
| 6. | Can we be confident in the outcome assessment? |
| 7. | Were all measured outcomes reported? |
| 8. | Were statistical methods appropriate? |
| 9. | Did researchers adhere to the study protocol? |
| 10. | Did the study design or analysis account for important confounding and modifying variables (unintended co-exposures) in experimental studies? |
| Extracted Study Details | |
|---|---|
| 1. | Study Information (author, year of publication) |
| 2. | Study Aims |
| 3. | Study Design |
| 4. | Cortisol Collection Method |
| 5. | Number of Participants |
| 6. | Participants Age |
| 7. | Participants Sex |
| 8. | Light Conditions |
| 9. | Light Correlated Colour Temperature (CCT) |
| 10. | Light Irradiance |
| 11. | Illuminance (lux) |
| 12. | Spectral Power Distribution Characteristics (peak and bandwidth) |
| 13. | Time of Light Exposure (24 h) |
| 14. | Duration of Light Exposure |
| 15. | Time of Sample Collection for Cortisol Measurement |
| 16. | Cortisol Yield per Condition |
| 17. | Time of Sample Collection for Melatonin Measurement (where applicable) |
| 18. | Melatonin Yield per Condition (where applicable) |
| Author | Question | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Babilon et al. [10] |  | |  | | |  | | | | |
| Cai et al. [40] | | | | | | | | | | |
| Cheung et al. [73] | |  | | | | | | | | |
| Choi et al. [43] | | | | | | | | | | |
| Danilenko et al. [44] | | | | | | | | | | |
| Figueiro et al. [45] | | | | | | | | | | |
| Ivanova et al. [57] | | | | | | | | | | |
| Petrowski et al. [41] | | | | | | | | | | |
| Petrowski at al. [9] | | | | | | | | | | |
| Petrowski et al. [42] | | | | | | | | | | |
| Sahin et al. [74] | | | | | | | | | | |
| Schmidt et al. [46] | | | | | | | | | | |
= Definitely high. = Probably high. = Probably low. = Definitely low.| Author | Study Aim | Study Design | Participants | Cortisol Collection Method | Melatonin Collection Method | ||
|---|---|---|---|---|---|---|---|
| N | Sex | Age: M (SD), range | |||||
| Babilon et al. [10] | To investigate the spectral dependency of morning cortisol secretion in humans using a consistent and practical method. | Within Participant | 4 | 4 M | 25.25 ± 3.59 | Saliva | - |
| Cai et al. [40] | To investigate the effect of lamps with similar power and Correlated Colour Temperatures (CCTs) but distinct spectra on melatonin and cortisol secretion. | Within Participant | 17 | 9 M, 8 F | 21–30 | Saliva | Saliva |
| Cheung et al. [73] | To determine the acute effects of morning versus evening, blue-enriched light exposure compared to dim light on hunger, metabolic function and physiological arousal. | Within Participant | 19 | 8 M, 11 F | Morning group = 26.0 ± 4.4, Evening group = 29.9 ± 6.1, 20–39 | Blood | - |
| Choi et al. [43] | To investigate physiological (melatonin, cortisol) and subjective responses to morning light exposure of lights with different CCTs. | Within Participant | 15 | 8 M, 7 F | 23.53 ± 3.37 | Saliva | Saliva |
| Danilenko et al. [44] | To investigate the role of melanopsin-based photoreception in the effects of light on reproductive hormones in females. | Between Participants | 16 | 16 F | 28.0 ± 7.2, 20–44 | Blood | Saliva |
| Figueiro et al. [45] | To investigate the impact of narrowband long-wavelength (red) and short-wavelength (blue) light exposures on the endocrine and autonomic systems (as measured by cortisol, alpha amylase and melatonin responses). | Within Participant | 12 | 8 M, 4 F | 19–53 | Saliva | Saliva |
| Ivanova et al. [57] | To investigate the impact of light on metabolism. | Within Participants | 10 | 10 F | 44.9 ± 12.3, 22–59 | Saliva | - |
| Petrowski et al. [41] | To investigate the influence of the spectral composition of light (brightness and wavelength) on the cortisol awakening response. | Within Participant | 53 (30 in Study 1, 23 in Study 2) | 53 M | Study 1 = 24.62 ± 3.32, Study 2 = 22.83 ± 3.33 | Saliva | - |
| Petrowski at al. [9] | To investigate the effect of light intensity and spectral composition on the cortisol response after the Maastricht Acute Stress Test (MAST) | Between Participants | 112 | 112 M | 24.83 ± 4.10 | Saliva | - |
| Petrowski et al. [42] | To compare the effects of short-wavelength blue light and bright light on cortisol secretion. | Within Participants | 49 (23 in Study 1, 26 in Study 2) | 49 M | Study 1 = 29.17 ± 7.57, Study 2 = 26.65 ± 7.00 | Saliva | - |
| Sahin et al. [74] | To investigate the effects of daytime light exposure on performance, biomarkers and alertness. | Within Participant | 13 | 6 M, 7 F | 23 ± 5.5 | Saliva | - |
| Schmidt et al. [46] | To investigate whether a blue-enriched light therapy device affects melatonin secretion, alertness, physiological stress and discomfort measures in the evening hours. | Within Participant | 17 | 7 M, 8 F | 22.8 ± 1.8 | Saliva | Saliva |
| Author | Condition | Intensity | Spectral Power Distribution | Light Exposure | Cortisol Measurement | Melatonin Measurement | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lux | W/m2 | Bandwidth | Peak (nm) | Half Max (nm) | CCT (K) | Start Time | Duration | ||||
| Babilon et al. [10] | Blue light | 24 ± 3 | 30.29 | Narrow | 476 | 13 | - | 6:00 | 2 h | 0, 20, 40, 60, 80 m | Not applicable |
| Red light | 22 ± 2 | 24.24 | Narrow | 649 | 14 | - | 6:00 | 2 h | 0, 20, 40, 60, 80 m | Not applicable | |
| Cai at al. [40] | Blue-enriched lamp 1 | 250 ± 50, 500 ± 50 | - | Broad | 453 | 5150 | 19:00 | 3.5 h | 0, 3, 4, 12 h | 0, 3, 4, 12 h | |
| Blue-enriched lamp 2 | 500 ± 50 | - | Broad | 467 | 5000 | 19:00 | 3.5 h | 0, 3, 4, 12 h | 0, 3, 4, 12 h | ||
| Blue-enriched lamp 3 | 250 ± 50, 500 ± 50 | - | Broad | 453, 467 | 5050 | 19:00 | 3.5 h | 0, 3, 4, 12 h | 0, 3, 4, 12 h | ||
| Dorm Lamp | - | Broad | 450 | 4000 | 19:00 | 3.5 h | 0, 3, 4, 12 h | 0, 3, 4, 12 h | |||
| Cheung et al. [73] | Dim light | <20 | - | - | - | From waking | 16 h | 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5, 4 h | Not-applicable | ||
| Blue-enriched light | 260 (Overhead), 370 (Blue) | - | Narrow blue and broad overhead lighting | 468 ± 8 (narrowband blue) | - | 07:50 or 17:45 | 3 h | 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5, 4 h | Not applicable | ||
| Choi et al. [43] | Dim light | <10 | - | Dynamic (film) | - | - | 9:00 (baseline) | 1 h | - | - | |
| Warm white light | 516.14 | 1.641 | Broad | 625 | - | 3590 | 10:00 | 1 h | 0, 1 h | 0, 1 h | |
| Blue-enriched white light | 518.38 | 1.793 | Broad | 460 | - | 6575 | 10:00 | 1 h | 0, 1 h | 0, 1 h | |
| Danilenko et al. [44] | Dim light (sunglasses during walk to study location) | <10 | - | - | - | - | - | - | 10–15 m | - | - |
| Red light | 1100 | 7 | Narrow | 651 | - | - | 7:30 | 45 m | 0, 22, 44 m | 0, 22, 44 m | |
| Blue-enriched white light | 1300 | 7 | Broad | 469 | - | - | 7:30 | 45 m | 0, 22, 44 m | 0, 22, 44 m | |
| Figueiro et al. [45] | Dim light | <3 | - | - | - | - | - | 8:00, 12:00, 16:00, 20:00, 00:00, 04:00, 08:00 | 1 h | 0, 1 h | 0, 1 h |
| Red light | 40 | 0.19 | Narrow | 625 | 25 | - | 8:00, 12:00, 16:00, 20:00, 00:00, 04:00, 08:00 | 1 h | 0, 1 h | 0, 1 h | |
| Blue light | 40 | 0.4 | Narrow | 470 | 25 | - | 8:00, 12:00, 16:00, 20:00, 00:00, 04:00, 08:00 | 1 h | 0, 1 h | 0, 1 h | |
| Ivanova et al. [57] | Dim light | <100 | - | - | - | - | - | 8:30 (pre-experiment baseline) | 15 m | - | Not applicable |
| Red light | 250 | - | Narrow | 620 | - | - | 9:00 | 30 m | 0, 30, 45 m | Not applicable | |
| Bright white light | 4300 | - | Broad | 550 and 620 | - | - | 9:00 | 30 m | 0, 30, 45 m | Not applicable | |
| Petrowski et al. [41] | Dim light | <2 | - | - | - | - | - | 5:05 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable |
| Bright white light | 414 | 1.566 | RGB | - | - | - | 5:05 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Red light | 235 | 1.341 | Narrow | 635 | - | - | 5:05 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Blue light | 201 | 1.76 | Narrow | 475 | - | - | 5:05 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Green light | 806 | 1.598 | Narrow | 520 | - | - | 5:05 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Petrowski at al. [9] | Dim light | <2 | 1.432 | RGB | - | - | - | 6:00 | 1 h | −15, −10, 0, 15, 30, 45, 60, 75, 90 m | Not applicable |
| Bright white light | 1240 | 1.566 | RGB | - | - | - | 6:00 | 1 h | −15, −10, 0, 15, 30, 45, 60, 75, 90 m | Not applicable | |
| Red light | 235 | 1.341 | Narrow | 635 | - | - | 6:00 | 1 h | −15, −10, 0, 15, 30, 45, 60, 75, 90 m | Not applicable | |
| Blue light | 201 | 1.76 | Narrow | 470–480 | - | - | 6:00 | 1 h | −15, −10, 0, 15, 30, 45, 60, 75, 90 m | Not applicable | |
| Petrowski et al. [42] | Dim light | <2 | - | - | - | - | - | 6:30 (baseline) | 1 h | - | Not applicable |
| Bright white light | 414 | - | - | - | - | - | 7:30 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Red light | 235 | - | Narrow | 635 | - | - | 7:30 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Blue light | 201 | - | Narrow | 470–480 | - | - | 7:30 | 1 h | 0, 15, 30, 45, 60, 75, 90, 105 m | Not applicable | |
| Sahin et al. [74] | Dim light | <5 | Fluorescent | - | - | 3500 | 6:00 (baseline) then 7:00, 11:00, 15:00 | 1 h (baseline), 2 h (experimental) | −10, 50, 110 m | Not applicable | |
| Red light glasses | 213 | 1.1 | - | 631 | 16 | - | 7:00, 11:00, 15:00 | 2 h | −10, 50, 110 m | Not applicable | |
| White light glasses | 361 | 1.12 | - | - | - | 2568 | 7:00, 11:00, 15:00 | 2 h | −10, 50, 110 m | Not applicable | |
| Schmidt et al. [46] | Dim light | <5 | - | - | - | - | - | 18:00 | 5 h | - | - |
| Head-mounted red light | 150 | 2.75 | Narrowband | 660 | - | - | 22:00 | 2 h | −4, −3, −2, −1, 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5 h | −4, −3, −2, −1, 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5 h | |
| Head-mounted, blue-enriched light | 1500 | 4.66 | Broadband | 460 | - | - | 22:00 | 2 h | −4, −3, −2, −1, 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5 h | −4, −3, −2, −1, 0, 0.5, 1, 1.5, 2, 2.5, 3, 3.5 h | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson-Dixon, I.; Murphy, M.J.; Crewther, S.G.; Riddell, N. The Influence of Light Wavelength on Human HPA Axis Rhythms: A Systematic Review. Life 2023, 13, 1968. https://doi.org/10.3390/life13101968
Robertson-Dixon I, Murphy MJ, Crewther SG, Riddell N. The Influence of Light Wavelength on Human HPA Axis Rhythms: A Systematic Review. Life. 2023; 13(10):1968. https://doi.org/10.3390/life13101968
Chicago/Turabian StyleRobertson-Dixon, Isabella, Melanie J. Murphy, Sheila G. Crewther, and Nina Riddell. 2023. "The Influence of Light Wavelength on Human HPA Axis Rhythms: A Systematic Review" Life 13, no. 10: 1968. https://doi.org/10.3390/life13101968
APA StyleRobertson-Dixon, I., Murphy, M. J., Crewther, S. G., & Riddell, N. (2023). The Influence of Light Wavelength on Human HPA Axis Rhythms: A Systematic Review. Life, 13(10), 1968. https://doi.org/10.3390/life13101968