
Environmental Stability and Its Importance for the Emergence of Darwinian Evolution

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
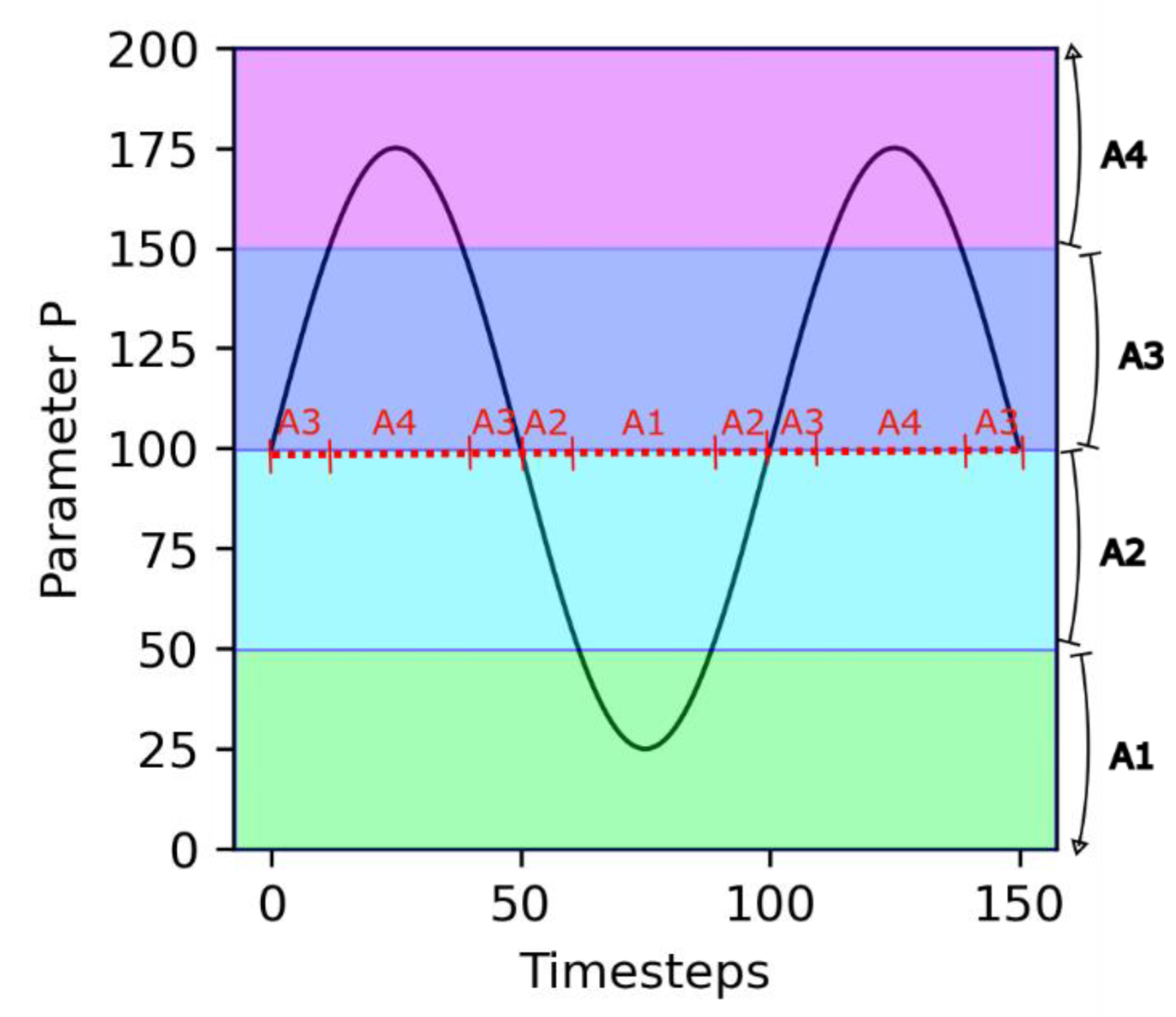
2. Presentation of the Model
3. Results
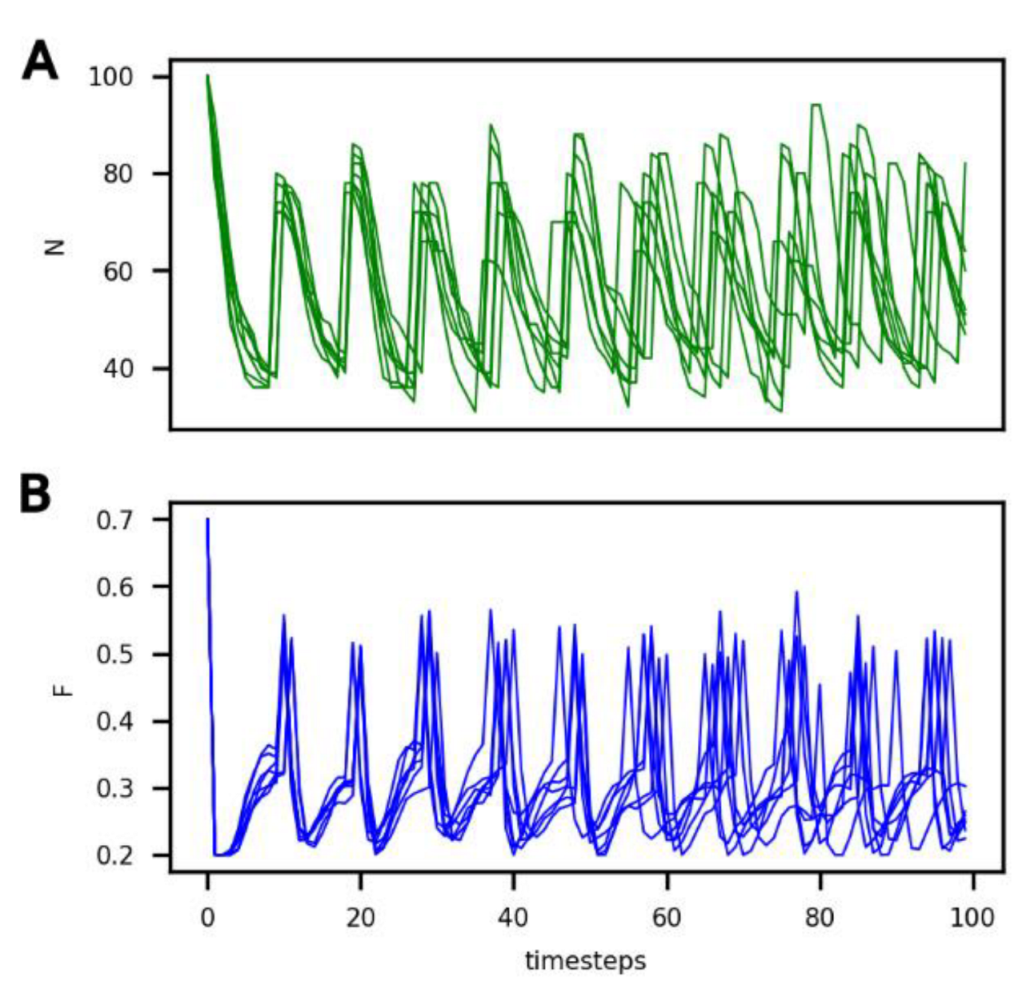
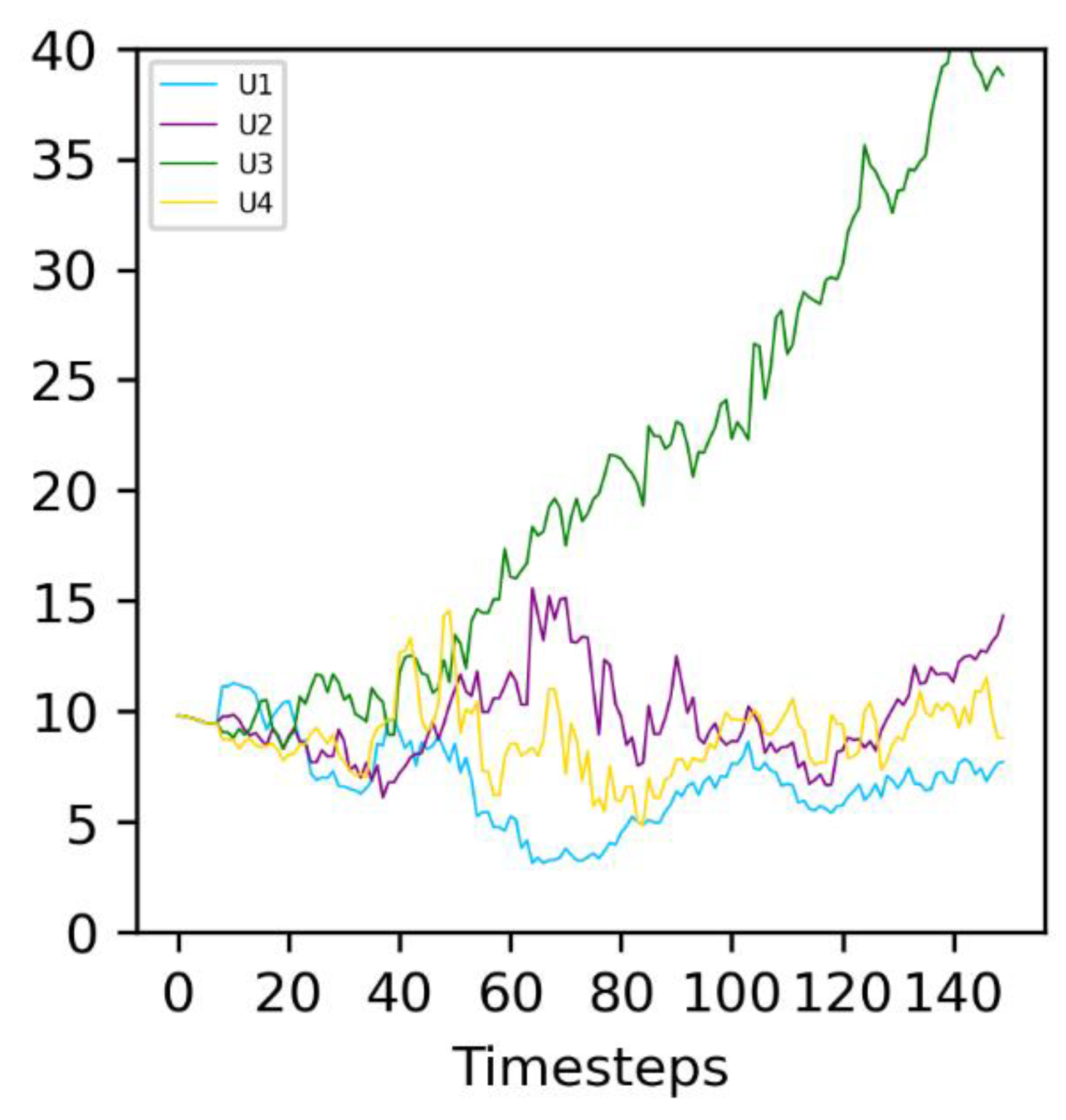
3.1. Reference Population
- (1)
- A common period of oscillations for N and F of approximately 10 timesteps, which can also be considered an average generation time;
- (2)
- A number of protocells in the compartment oscillating between 40 and 80; and
- (3)
- An amount of food in the compartment oscillating between 0.2 and 0.5.
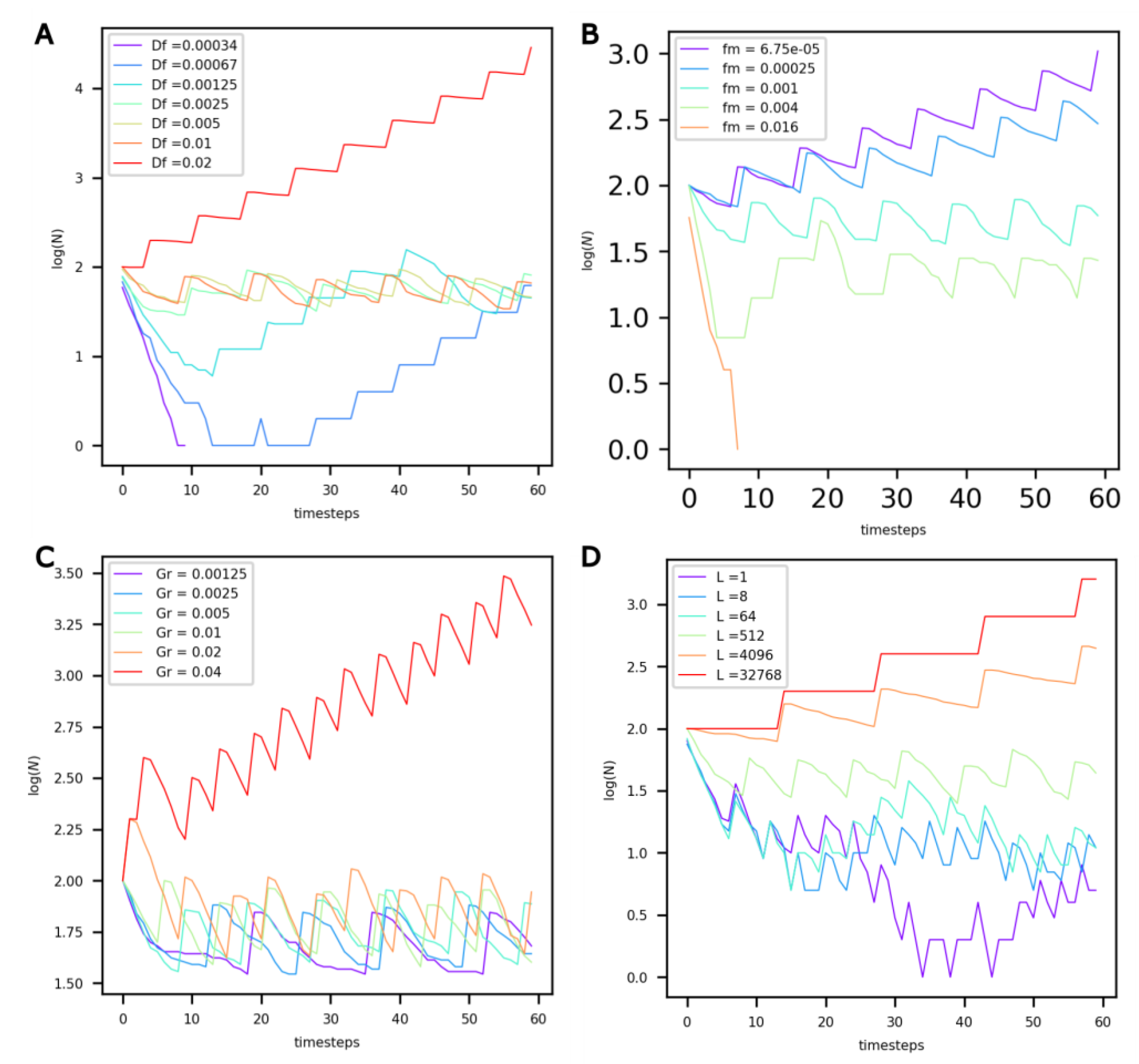
3.2. Influence of Protocell Constants
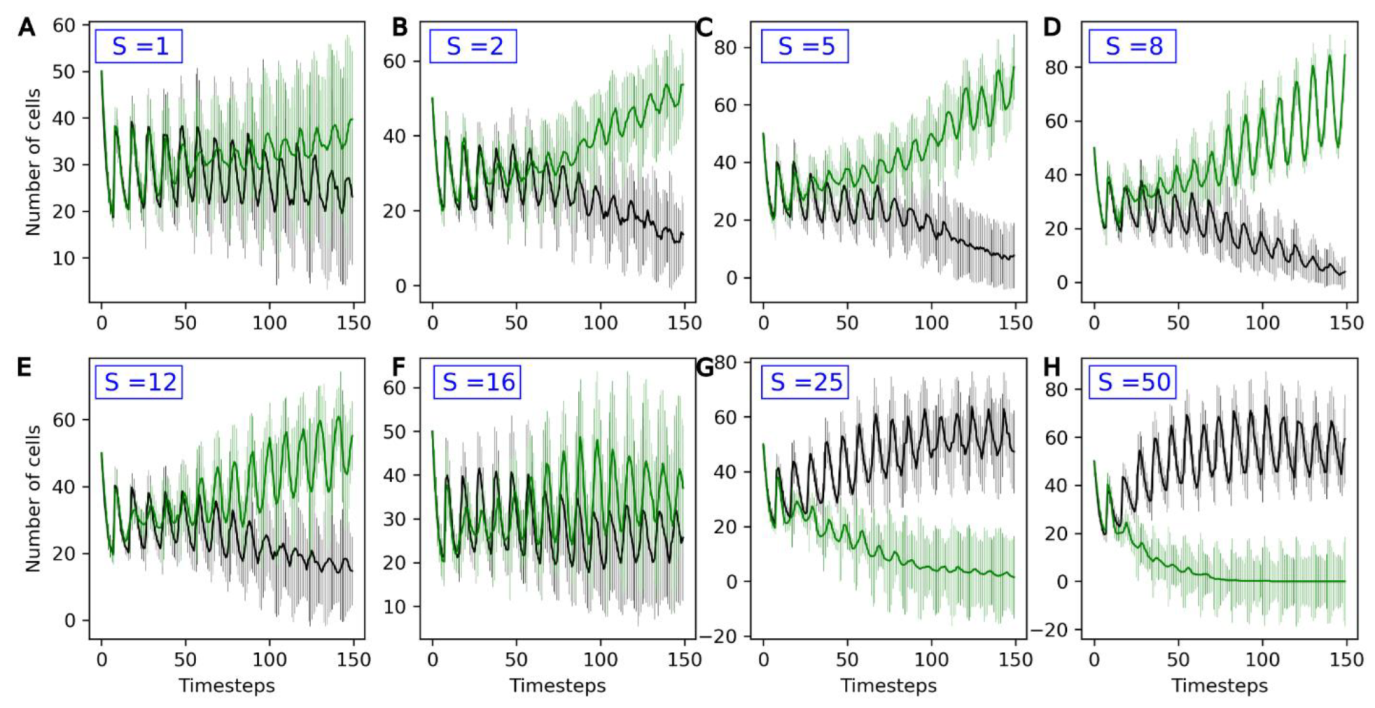
3.3. Competition between Evolving and Non-Evolving Population—Stable Environment
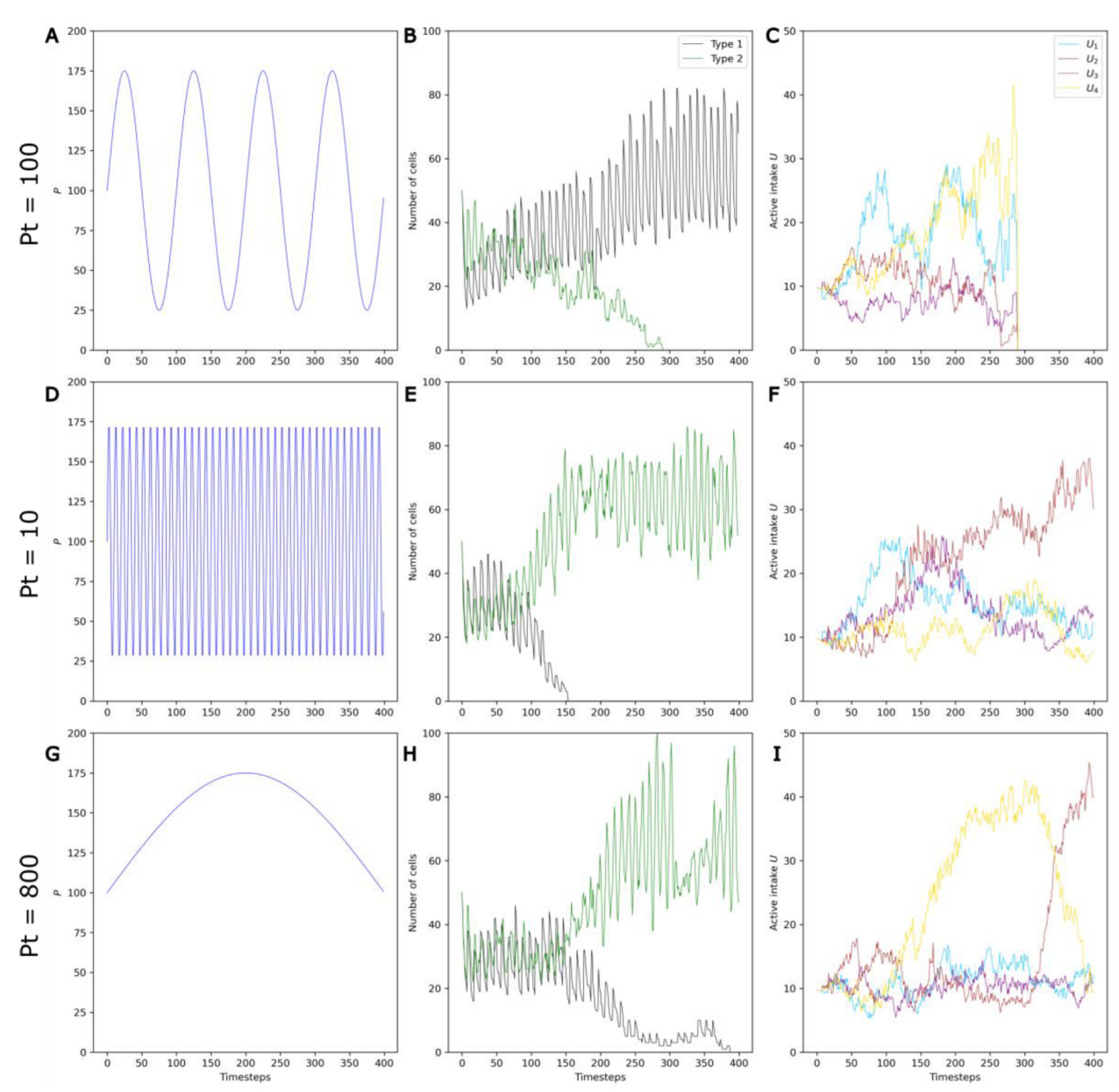
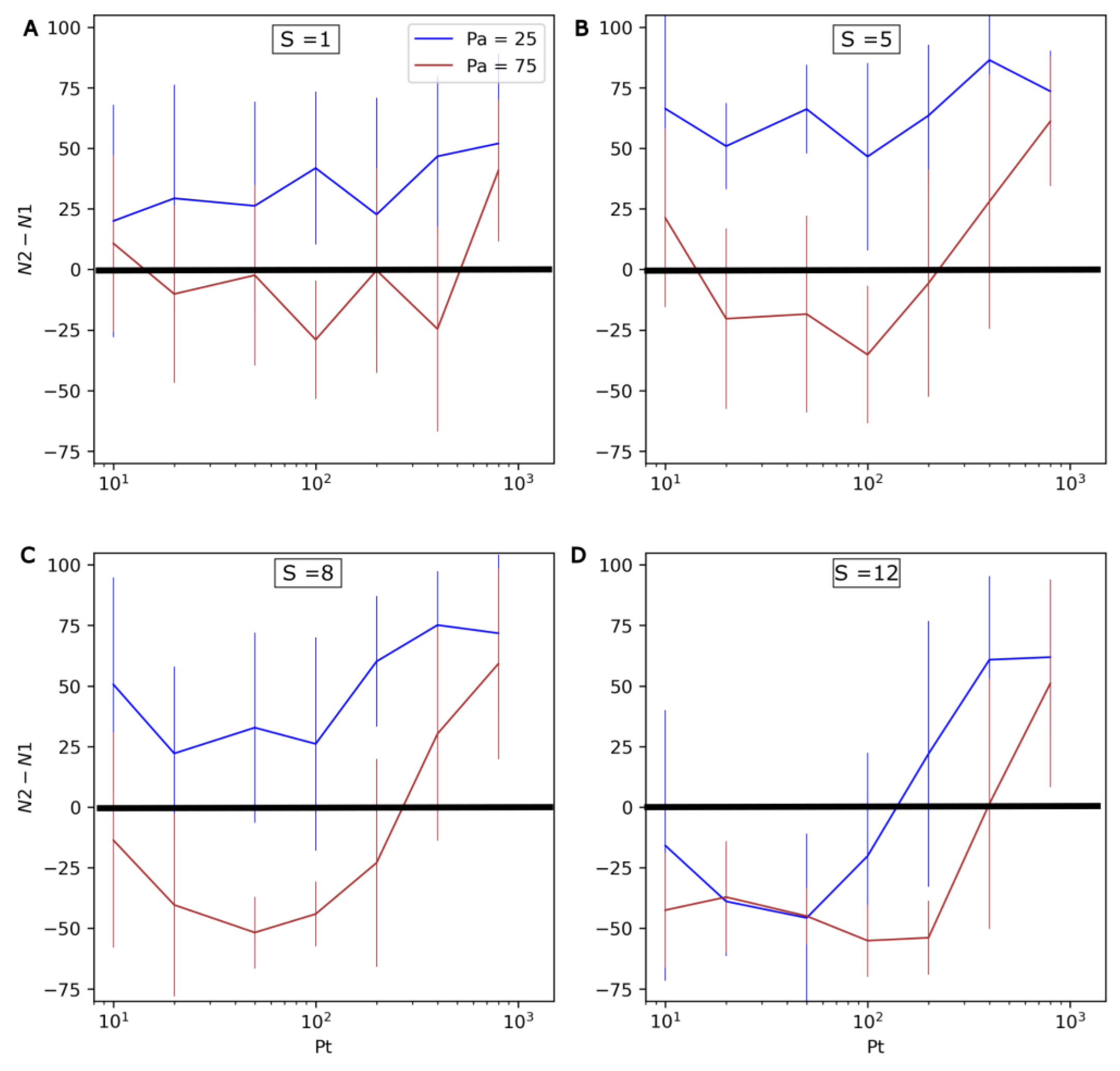
3.4. Competition between Evolving and Non-Evolving Populations—Changing Environment
4. Discussion
4.1. Comparison of the Model with Hydrothermal Environments and Single-Celled Organisms
4.2. Limitations of the Model
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Nomenclature
| N | Number of protocells in the compartment (no unit) |
| N0 | Initial number of cells |
| F | Concentration in food molecules in the compartment (mmol·L−1) |
| F0 | Initial concentration of food in the compartment |
| A1, A2, A3, A4 | Concentrations in molecules A1, A2, A3 and A4 in one protocell (mmol·m−3) |
| A1,C, A2,C, A3,C, A4,C | Concentrations in molecules A1, A2, A3 and A4 in the compartment (mmol·m−3) |
| Fi | Food input rate in the compartment (mmol·m−3·s−1) |
| P | Environmental parameter (no unit) |
| Pt | Period of P oscillations (s) |
| Pa | Amplitude of P oscillations (no unit) |
| Df | Constant for food diffusion between protocells and the environment (mmol·s−1·m−2) |
| V | Volume of a protocell (m3) |
| D | Constant for diffusion of molecules A1, …, A4 between protocells and the environment (m·s−1) |
| U1, U2, U3, U4 | Rates of active intake molecules A1, …, A4 by protocells (mmol·s−1·m−2) |
| fm | Constant for the food required for protocell maintenance (mmol·m−3·s−1) |
| fu | Constant for the food required for active intake of molecules A1, …, A4 (m2) |
| Gr | Constant for protocell growth ((m3·mmol−1)3) |
| L | Constant for protocell death probability (mmol−1) |
| S | Standard deviation of the distribution of change of U1, …, U4 between generations (mmol·s−1·m−2) |
References
- Darwin, C. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life; Natural History Museum: London, UK, 1859; ISBN 978-0-565-09502-4. [Google Scholar]
- Benner, S.A.; Kim, H.-J.; Yang, Z. Setting the Stage: The History, Chemistry, and Geobiology behind RNA. Cold Spring Harb. Perspect. Biol. 2012, 4, a003541. [Google Scholar] [CrossRef] [PubMed]
- Higgs, P.G.; Lehman, N. The RNA World: Molecular Cooperation at the Origins of Life. Nat. Rev. Genet. 2015, 16, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. Prebiotic Chemistry and the Origin of the RNA World. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 99–123. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Russell, M.J. On the Origin of Biochemistry at an Alkaline Hydrothermal Vent. Phil. Trans. R. Soc. B 2007, 362, 1887–1926. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Anollés, D.; Kim, K.M.; Mittenthal, J.E.; Caetano-Anollés, G. Proteome Evolution and the Metabolic Origins of Translation and Cellular Life. J. Mol. Evol. 2011, 72, 14–33. [Google Scholar] [CrossRef]
- Peretó, J. Out of Fuzzy Chemistry: From Prebiotic Chemistry to Metabolic Networks. Chem. Soc. Rev. 2012, 41, 5394. [Google Scholar] [CrossRef]
- Nitschke, W.; Russell, M.J. Beating the Acetyl Coenzyme A-Pathway to the Origin of Life. Phil. Trans. R. Soc. B 2013, 368, 20120258. [Google Scholar] [CrossRef]
- Tran, Q.P.; Adam, Z.R.; Fahrenbach, A.C. Prebiotic Reaction Networks in Water. Life 2020, 10, 352. [Google Scholar] [CrossRef]
- Goldford, J.E.; Smith, H.B.; Longo, L.M.; Wing, B.A.; McGlynn, S.E. Continuity between Ancient Geochemistry and Modern Metabolism Enabled by Non-Autocatalytic Purine Biosynthesis. Syst. Biol. 2022. preprint. [Google Scholar]
- Yadav, M.; Pulletikurti, S.; Yerabolu, J.R.; Krishnamurthy, R. Cyanide as a Primordial Reductant Enables a Protometabolic Reductive Glyoxylate Pathway. Nat. Chem. 2022, 14, 170–178. [Google Scholar] [CrossRef]
- Gánti, T. Chemoton Theory; Mathematical and Computational Chemistry; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003; ISBN 978-0-306-47785-0. [Google Scholar]
- Hanczyc, M.M.; Szostak, J.W. Replicating Vesicles as Models of Primitive Cell Growth and Division. Curr. Opin. Chem. Biol. 2004, 8, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Budin, I.; Szostak, J.W. Physical Effects Underlying the Transition from Primitive to Modern Cell Membranes. Proc. Natl. Acad. Sci. USA 2011, 108, 5249–5254. [Google Scholar] [CrossRef] [PubMed]
- Murtas, G. Early Self-Reproduction, the Emergence of Division Mechanisms in Protocells. Mol. BioSyst. 2013, 9, 195–204. [Google Scholar] [CrossRef]
- Caspi, Y.; Dekker, C. Divided We Stand: Splitting Synthetic Cells for Their Proliferation. Syst. Synth. Biol. 2014, 8, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.A.; Nguyen, D.H.; Wexler, T.B.; Goldman, A.D. The Coevolution of Cellularity and Metabolism Following the Origin of Life. J. Mol. Evol. 2020, 88, 598–617. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Nozawa, R.-S.; Jia, T.Z.; Saio, T.; Mori, E. Biological Phase Separation: Cell Biology Meets Biophysics. Biophys. Rev. 2020, 12, 519–539. [Google Scholar] [CrossRef]
- Kotopoulou, E.; Lopez-Haro, M.; Calvino Gamez, J.J.; García-Ruiz, J.M. Nanoscale Anatomy of Iron-Silica Self-Organized Membranes: Implications for Prebiotic Chemistry. Angew. Chem. Int. Ed. 2021, 60, 1396–1402. [Google Scholar] [CrossRef]
- Gözen, I.; Köksal, E.S.; Põldsalu, I.; Xue, L.; Spustova, K.; Pedrueza-Villalmanzo, E.; Ryskulov, R.; Meng, F.; Jesorka, A. Protocells: Milestones and Recent Advances. Small 2022, 18, 2106624. [Google Scholar] [CrossRef]
- Kauffman, S.A.; Farmer, J.D. Autocatalytic Sets of Proteins. Orig. Life Evol Biosph. 1986, 16, 446–447. [Google Scholar] [CrossRef]
- Hunding, A.; Kepes, F.; Lancet, D.; Minsky, A.; Norris, V.; Raine, D.; Sriram, K.; Root-Bernstein, R. Compositional Complementarity and Prebiotic Ecology in the Origin of Life. Bioessays 2006, 28, 399–412. [Google Scholar] [CrossRef]
- Shapiro, R. A Simpler Origin for Life. Sci. Am. 2007, 296, 46–53. [Google Scholar] [CrossRef]
- Vasas, V.; Fernando, C.; Santos, M.; Kauffman, S.; Szathmáry, E. Evolution before Genes. Biol. Direct 2012, 7, 1–14. [Google Scholar] [CrossRef]
- Jia, T.Z.; Caudan, M.; Mamajanov, I. Origin of Species before Origin of Life: The Role of Speciation in Chemical Evolution. Life 2021, 11, 154. [Google Scholar] [CrossRef]
- Saha, A.; Yi, R.; Fahrenbach, A.C.; Wang, A.; Jia, T.Z. A Physicochemical Consideration of Prebiotic Microenvironments for Self-Assembly and Prebiotic Chemistry. Life 2022, 12, 1595. [Google Scholar] [CrossRef] [PubMed]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The Origin of Life in Alkaline Hydrothermal Vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Cryosphere and Psychrophiles: Insights into a Cold Origin of Life? Life 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Bréhéret, J.G.; Foucher, F.; Hubert, A.; Sorieul, S.; Dass, A.V.; et al. A Hydrothermal-Sedimentary Context for the Origin of Life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; van Zuilen, M.A.; Bach, W. Mineral Self-Organization on a Lifeless Planet. Phys. Life Rev. 2020, 34–35, 62–82. [Google Scholar] [CrossRef]
- Fiore, M.; Chieffo, C.; Lopez, A.; Fayolle, D.; Ruiz, J.; Soulère, L.; Oger, P.; Altamura, E.; Popowycz, F.; Buchet, R. Synthesis of Phospholipids Under Plausible Prebiotic Conditions and Analogies with Phospholipid Biochemistry for Origin of Life Studies. Astrobiology 2022, 22, 598–627. [Google Scholar] [CrossRef]
- Bates, A.E.; Lee, R.W.; Tunnicliffe, V.; Lamare, M.D. Deep-Sea Hydrothermal Vent Animals Seek Cool Fluids in a Highly Variable Thermal Environment. Nat. Commun. 2010, 1, 14. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.M.; Idei, A.; Nakagawa, M.; Ueno, Y.; Fischer, W.W.; McGlynn, S.E. Geochemical and Metagenomic Characterization of Jinata Onsen, a Proterozoic-Analog Hot Spring, Reveals Novel Microbial Diversity Including Iron-Tolerant Phototrophs and Thermophilic Lithotrophs. Microbiology 2018. preprint. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Munoz-Saez, C.; Wilmeth, D.T.; Myers, K.D.; Homann, M.; Arp, G.; Skok, J.R.; Zuilen, M.A. Morphogenesis of Digitate Structures in Hot Spring Silica Sinters of the El Tatio Geothermal Field, Chile. Geobiology 2022, 20, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Froese, T.; Virgo, N.; Ikegami, T. Motility at the Origin of Life: Its Characterization and a Model. Artif. Life 2014, 20, 55–76. [Google Scholar] [CrossRef]
- Fick, A. Ueber Diffusion. Ann. Phys. Chem. 1855, 170, 59–86. [Google Scholar] [CrossRef]
- Eigen, M. Selforganization of Matter and the Evolution of Biological Macromolecules. Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef]
- De Bremond d’Ars, J.; Gibert, D. Low-Temperature Hydrothermal Systems Response to Rainfall Forcing: An Example From Temperature Time Series of Fumaroles at La Soufrière de Guadeloupe Volcano. Front. Earth Sci. 2022, 9, 772176. [Google Scholar] [CrossRef]
- Monod, J. The Growth of Bacterial Cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef]
- Eagon, R.G. Pseudomonas Natriegens, a Marine Bacterium with a Generation Time of less than 10 Minutes. J. Bacteriol. 1962, 83, 736–737. [Google Scholar] [CrossRef]
- Rappé, M.S.; Connon, S.A.; Vergin, K.L.; Giovannoni, S.J. Cultivation of the Ubiquitous SAR11 Marine Bacterioplankton Clade. Nature 2002, 418, 630–633. [Google Scholar] [CrossRef]
- Weissman, J.L.; Hou, S.; Fuhrman, J.A. Estimating Maximal Microbial Growth Rates from Cultures, Metagenomes, and Single Cells via Codon Usage Patterns. Proc. Natl. Acad. Sci. USA 2021, 118, e2016810118. [Google Scholar] [CrossRef] [PubMed]
- Colwell, F.S.; D’Hondt, S. Nature and Extent of the Deep Biosphere. Rev. Mineral. Geochem. 2013, 75, 547–574. [Google Scholar] [CrossRef]
- Zhu, T.F.; Szostak, J.W. Coupled Growth and Division of Model Protocell Membranes. J. Am. Chem. Soc. 2009, 131, 5705–5713. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Wick, R.; Fresta, M.; Mangone, A.; Luisi, P.L. Autopoietic Self-Reproduction of Fatty Acid Vesicles. J. Am. Chem. Soc. 1994, 116, 11649–11654. [Google Scholar] [CrossRef]
- Lynch, M. Evolution of the Mutation Rate. Trends Genet. 2010, 26, 345–352. [Google Scholar] [CrossRef]
- Van Dijk, D.; Dhar, R.; Missarova, A.M.; Espinar, L.; Blevins, W.R.; Lehner, B.; Carey, L.B. Slow-Growing Cells within Isogenic Populations Have Increased RNA Polymerase Error Rates and DNA Damage. Nat. Commun. 2015, 6, 7972. [Google Scholar] [CrossRef]
- Potapov, V.; Fu, X.; Dai, N.; Corrêa, I.R.; Tanner, N.A.; Ong, J.L. Base Modifications Affecting RNA Polymerase and Reverse Transcriptase Fidelity. Nucleic Acids Res. 2018, 46, 5753–5763. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daga, K.R.; Feray Çoşar, M.; Lowenkron, A.; Hao, J.; Rouillard, J. Environmental Stability and Its Importance for the Emergence of Darwinian Evolution. Life 2023, 13, 1960. https://doi.org/10.3390/life13101960
Daga KR, Feray Çoşar M, Lowenkron A, Hao J, Rouillard J. Environmental Stability and Its Importance for the Emergence of Darwinian Evolution. Life. 2023; 13(10):1960. https://doi.org/10.3390/life13101960
Chicago/Turabian StyleDaga, Khushi R., Mensura Feray Çoşar, Abigail Lowenkron, Jihua Hao, and Joti Rouillard. 2023. "Environmental Stability and Its Importance for the Emergence of Darwinian Evolution" Life 13, no. 10: 1960. https://doi.org/10.3390/life13101960
APA StyleDaga, K. R., Feray Çoşar, M., Lowenkron, A., Hao, J., & Rouillard, J. (2023). Environmental Stability and Its Importance for the Emergence of Darwinian Evolution. Life, 13(10), 1960. https://doi.org/10.3390/life13101960