

Alternative Methods as Tools for Obesity Research: In Vitro and In Silico Approaches

 , , ,
, , ,  , , , , and
, , , , and 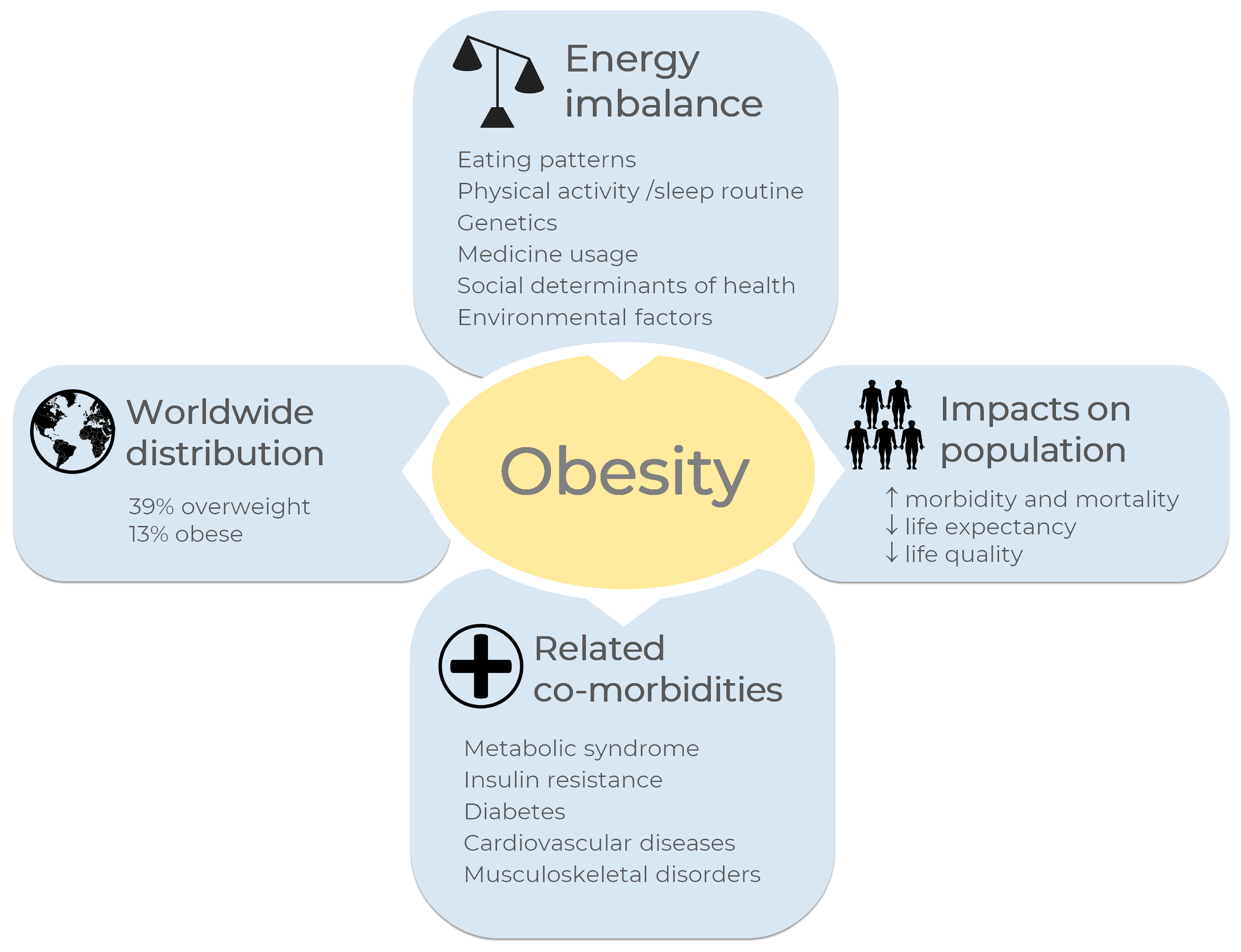{kind=link}
{kind=link}
{kind=link}
{kind=link}
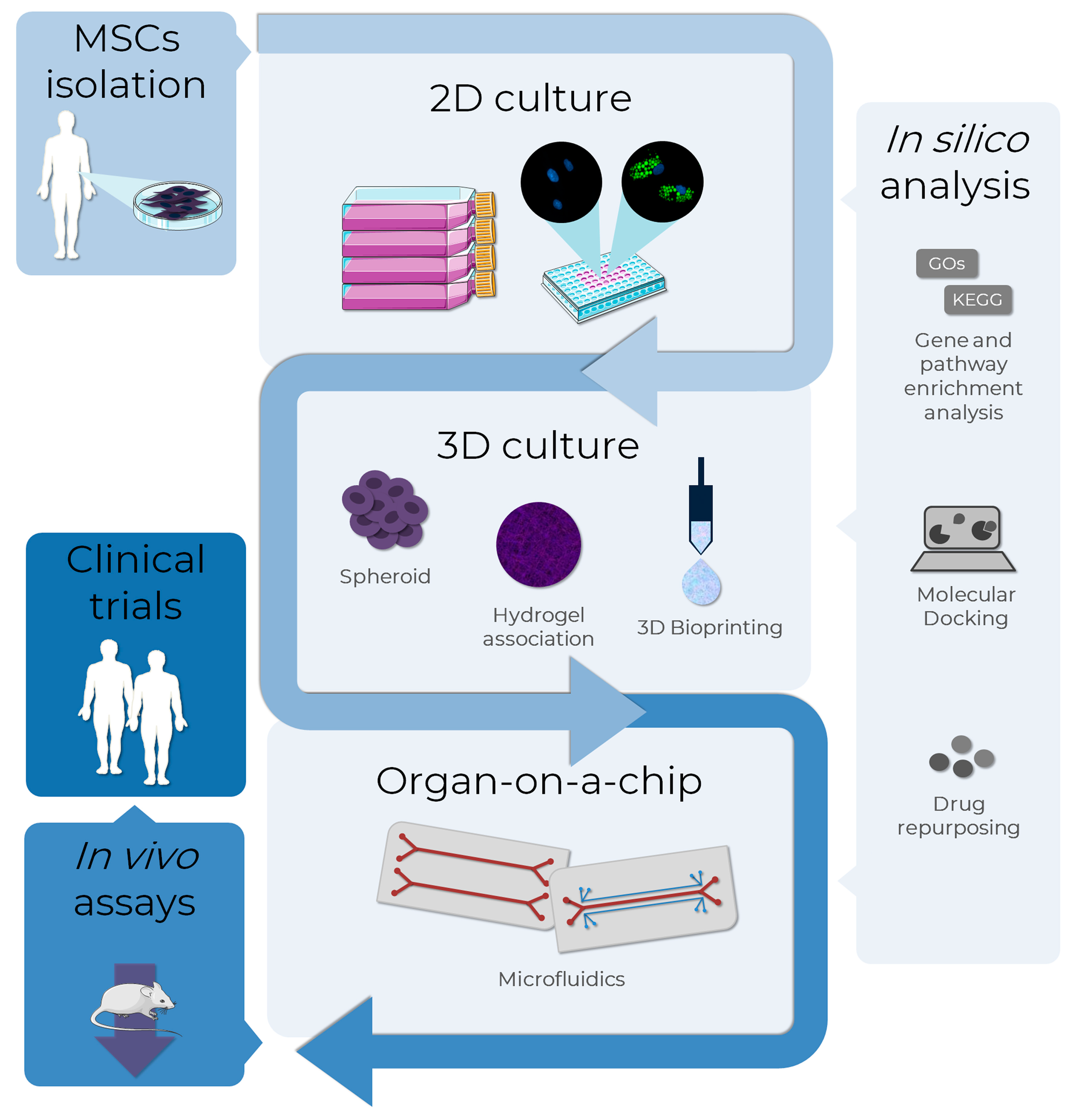{kind=link}
Abstract
1. Introduction
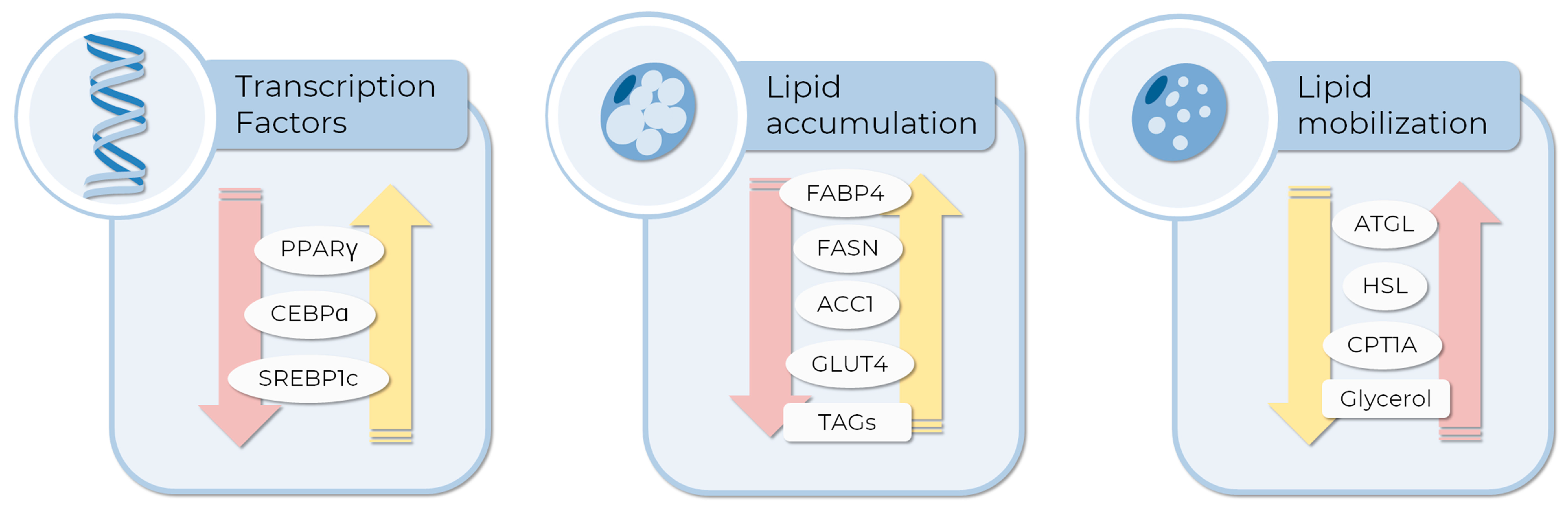
2. Gene Expression in Adipogenesis and Lipolysis
3. Metabolic Features as Endpoints to Evaluate Adipogenesis and Lipolysis in MSCs Models
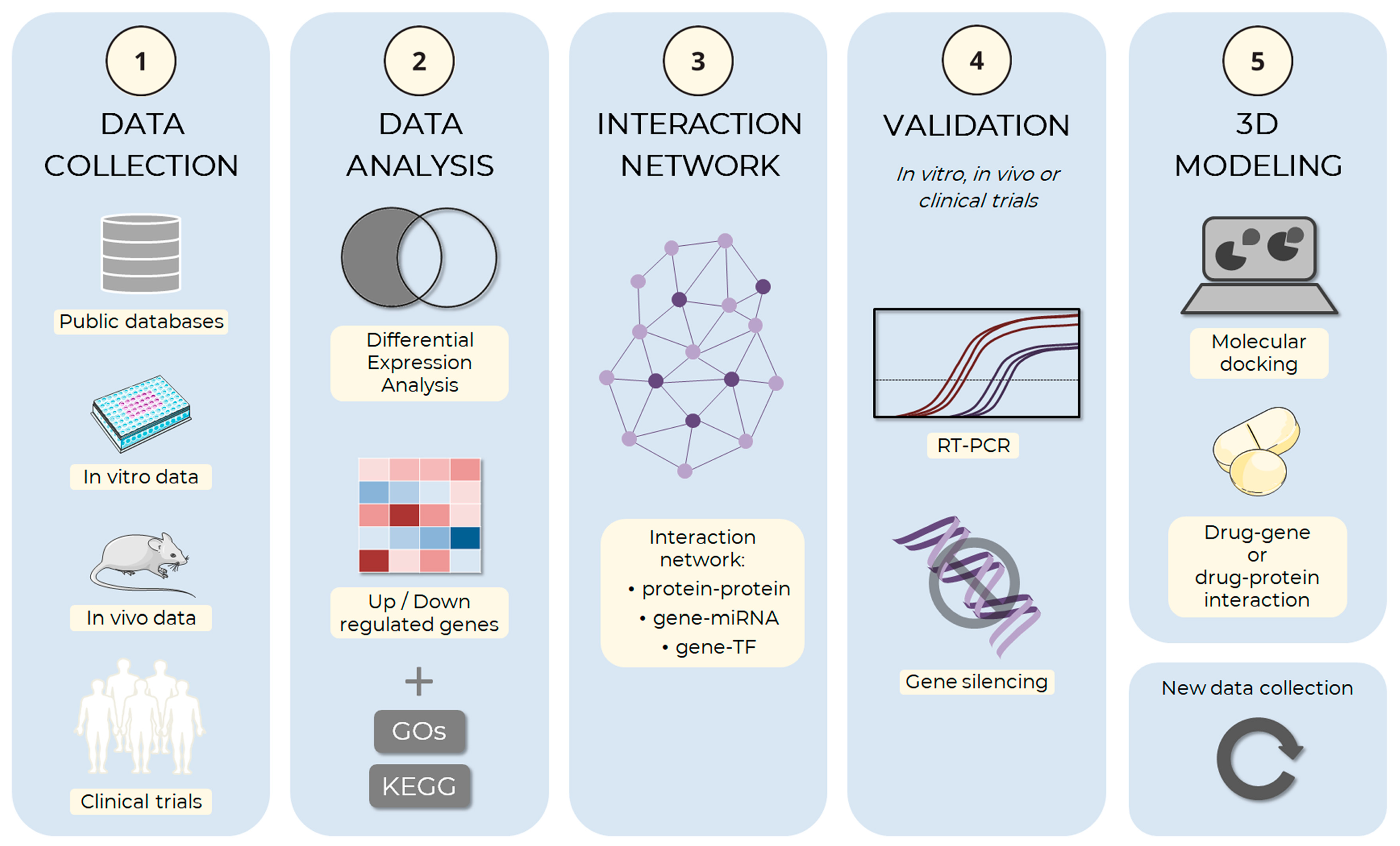
4. Alternative Methods to Animal Testing in Adipogenesis
4.1. In Silico Methods May Provide the Basis for Elucidating the Mechanisms Underlying Adipogenesis
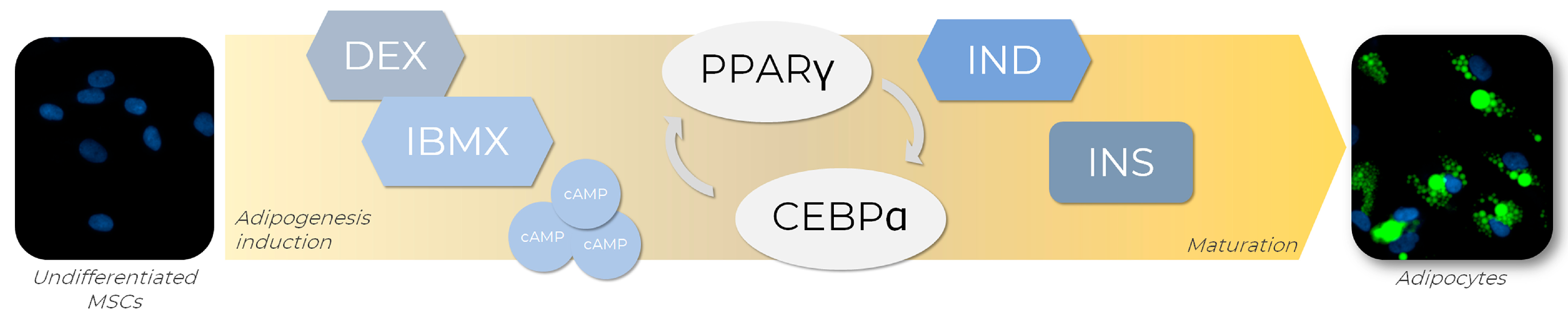
4.2. Inducers for Adipogenesis In Vitro
4.3. D Culture as a Key Model for Adipogenesis Studies
4.3.1. Morphological Evaluation of In Vitro Adipogenesis
4.3.2. D Systems Mimetic the Organ Architecture
4.4. Bioprinting as a New Tool for 3D Systems
4.5. Microfluidics
5. Challenges and Limitations
6. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO, W.H.O. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 11 December 2022).
- Abdelaal, M.; le Roux, C.W.; Docherty, N.G. Morbidity and Mortality Associated with Obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef] [PubMed]
- Safaei, M.; Sundararajan, E.A.; Driss, M.; Boulila, W.; Shapi’i, A. A Systematic Literature Review on Obesity: Understanding the Causes & Consequences of Obesity and Reviewing Various Machine Learning Approaches Used to Predict Obesity. Comput. Biol. Med. 2021, 136, 104754. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Overweight and Obesity: Causes and Consequences. Available online: https://www.cdc.gov/obesity/basics/causes.html?CDC_AA_refVal=https%3A%2F%2Fwww.cdc.gov%2Fobesity%2Fadult%2Fcauses.html (accessed on 11 December 2022).
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-Obesity Drug Discovery: Advances and Challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.; Hambly, C.; Mitchell, S.; Król, E. The Contribution of Animal Models to the Study of Obesity. Lab. Anim. 2008, 42, 413–432. [Google Scholar] [CrossRef]
- Barrett, P.; Mercer, J.G.; Morgan, P.J. Preclinical Models for Obesity Research. Dis. Model. Mech. 2016, 9, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, J.; Mohamed, M.; Bakar, A. A Systematic Review on Different Models of Inducing Obesity in Animals: Advantages and Limitations. J. Adv. Vet. Anim. Res. 2020, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem Cells: Their Source, Potency and Use in Regenerative Therapies with Focus on Adipose-Derived Stem Cells—A Review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.; Rupérez, A.; Gomez-Llorente, C.; Gil, A.; Aguilera, C. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Song, H.-D.; Li, R.-Y.; Zhou, L.-B.; Zhu, Z.-D.; Hu, R.-M.; Han, Z.-G.; Chen, J.-L. The Gene Expression Profiling of Human Visceral Adipose Tissue and Its Secretory Functions. Biochem. Biophys. Res. Commun. 2003, 300, 839–846. [Google Scholar] [CrossRef]
- Schleinitz, D.; Krause, K.; Wohland, T.; Gebhardt, C.; Linder, N.; Stumvoll, M.; Blüher, M.; Bechmann, I.; Kovacs, P.; Gericke, M.; et al. Identification of Distinct Transcriptome Signatures of Human Adipose Tissue from Fifteen Depots. Eur. J. Hum. Genet. 2020, 28, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Raajendiran, A.; Krisp, C.; de Souza, D.P.; Ooi, G.; Burton, P.R.; Taylor, R.A.; Molloy, M.P.; Watt, M.J. Proteome Analysis of Human Adipocytes Identifies Depot-Specific Heterogeneity at Metabolic Control Points. Am. J. Physiol.-Endocrinol. Metab. 2021, 320, E1068–E1084. [Google Scholar] [CrossRef] [PubMed]
- Norreen-Thorsen, M.; Struck, E.C.; Öling, S.; Zwahlen, M.; von Feilitzen, K.; Odeberg, J.; Lindskog, C.; Pontén, F.; Uhlén, M.; Dusart, P.J.; et al. A Human Adipose Tissue Cell-Type Transcriptome Atlas. Cell Rep. 2022, 40, 111046. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.D.; Soh, J.Y.; Innis, S.E.; Dimanche, A.; Ma, L.; Langefeld, C.D.; Comeau, M.E.; Das, S.K.; Schadt, E.E.; Björkegren, J.L.M.; et al. Sex Differences in Human Adipose Tissue Gene Expression and Genetic Regulation Involve Adipogenesis. Genome Res. 2020, 30, 1379–1392. [Google Scholar] [CrossRef]
- Robert, A.W.; Marcon, B.H.; Dallagiovanna, B.; Shigunov, P. Adipogenesis, Osteogenesis, and Chondrogenesis of Human Mesenchymal Stem/Stromal Cells: A Comparative Transcriptome Approach. Front. Cell Dev. Biol. 2020, 8, 561. [Google Scholar] [CrossRef]
- Yi, X.; Wu, P.; Liu, J.; Gong, Y.; Xu, X.; Li, W. Identification of the Potential Key Genes for Adipogenesis from Human Mesenchymal Stem Cells by RNA-Seq. J. Cell. Physiol. 2019, 234, 20217–20227. [Google Scholar] [CrossRef]
- Jeong, J.A.; Ko, K.-M.; Park, H.S.; Lee, J.; Jang, C.; Jeon, C.-J.; Koh, G.Y.; Kim, H. Membrane Proteomic Analysis of Human Mesenchymal Stromal Cells during Adipogenesis. Proteomics 2007, 7, 4181–4191. [Google Scholar] [CrossRef]
- Foley, B.; Doheny, D.L.; Black, M.B.; Pendse, S.N.; Wetmore, B.A.; Clewell, R.A.; Andersen, M.E.; Deisenroth, C. Editor’s Highlight: Screening ToxCast Prioritized Chemicals for PPARG Function in a Human Adipose-Derived Stem Cell Model of Adipogenesis. Toxicol. Sci. 2017, 155, 85–100. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ Is Required for the Differentiation of Adipose Tissue In Vivo and In Vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Evans, R.M.; Barish, G.D.; Wang, Y.-X. PPARs and the Complex Journey to Obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef]
- Vidal-Puig, A.J.; van Considine, R.; Jimenez-Liñan, M.; Werman, A.; Pories, W.J.; Caro, J.F.; Flier, J.S. Peroxisome Proliferator-Activated Receptor Gene Expression in Human Tissues. Effects of Obesity, Weight Loss, and Regulation by Insulin and Glucocorticoids. J. Clin. Investig. 1997, 99, 2416–2422. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.R.; Douagi, I.; Andersson, D.P.; Bäckdahl, J.; Rydén, M.; Arner, P.; Laurencikiene, J. Increased Fat Cell Size: A Major Phenotype of Subcutaneous White Adipose Tissue in Non-Obese Individuals with Type 2 Diabetes. Diabetologia 2016, 59, 560–570. [Google Scholar] [CrossRef]
- Matulewicz, N.; Stefanowicz, M.; Nikołajuk, A.; Karczewska-Kupczewska, M. Markers of Adipogenesis, but Not Inflammation, in Adipose Tissue Are Independently Related to Insulin Sensitivity. J. Clin. Endocrinol. Metab. 2017, 102, 3040–3049. [Google Scholar] [CrossRef] [PubMed]
- Conley, S.M.; Hickson, L.J.; Kellogg, T.A.; McKenzie, T.; Heimbach, J.K.; Taner, T.; Tang, H.; Jordan, K.L.; Saadiq, I.M.; Woollard, J.R.; et al. Human Obesity Induces Dysfunction and Early Senescence in Adipose Tissue-Derived Mesenchymal Stromal/Stem Cells. Front. Cell Dev. Biol. 2020, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Desai, A.; Yang, Z.; Sharma, A.; DeSouza, T.; Genga, R.M.J.; Kucukural, A.; Lifshitz, L.M.; Nielsen, S.; Scheele, C.; et al. Diverse Repertoire of Human Adipocyte Subtypes Develops from Transcriptionally Distinct Mesenchymal Progenitor Cells. Proc. Natl. Acad. Sci. USA 2019, 116, 17970–17979. [Google Scholar] [CrossRef]
- Bolsoni-Lopes, A.; Alonso-Vale, M.I.C. Lipolysis and Lipases in White Adipose Tissue—An Update. Arch. Endocrinol. Metab. 2015, 59, 335–342. [Google Scholar] [CrossRef]
- Wei, E.; Lehner, R.; Vance, D.E. C/EBPα Activates the Transcription of Triacylglycerol Hydrolase in 3T3-L1 Adipocytes. Biochem. J. 2005, 388, 959–966. [Google Scholar] [CrossRef]
- Xu, Q.; Mariman, E.C.M.; Roumans, N.J.T.; Vink, R.G.; Goossens, G.H.; Blaak, E.E.; Jocken, J.W.E. Adipose Tissue Autophagy Related Gene Expression Is Associated with Glucometabolic Status in Human Obesity. Adipocyte 2018, 7, 12–19. [Google Scholar] [CrossRef]
- lo Furno, D.; Graziano, A.C.E.; Avola, R.; Giuffrida, R.; Perciavalle, V.; Bonina, F.; Mannino, G.; Cardile, V. A Citrus Bergamia Extract Decreases Adipogenesis and Increases Lipolysis by Modulating PPAR Levels in Mesenchymal Stem Cells from Human Adipose Tissue. PPAR Res. 2016, 2016, 4563815. [Google Scholar] [CrossRef]
- Bu, S.; Yuan, C.Y.; Xue, Q.; Chen, Y.; Cao, F. Bilobalide Suppresses Adipogenesis in 3T3-L1 Adipocytes via the AMPK Signaling Pathway. Molecules 2019, 24, 3503. [Google Scholar] [CrossRef]
- Rafalski, V.A.; Mancini, E.; Brunet, A. Energy Metabolism and Energy-Sensing Pathways in Mammalian Embryonic and Adult Stem Cell Fate. J. Cell Sci. 2012, 125, 5597–5608. [Google Scholar] [CrossRef] [PubMed]
- Shyh-Chang, N.; Daley, G.Q.; Cantley, L.C. Stem Cell Metabolism in Tissue Development and Aging. Development 2013, 140, 2535–2547. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L. Adipose Tissue Oxygenation. Adipocyte 2014, 3, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Turcotte, R.; et al. Direct Measurement of Local Oxygen Concentration in the Bone Marrow of Live Animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef]
- Chen, C.-T.; van Shih, Y.-R.; Kuo, T.K.; Lee, O.K.; Wei, Y.-H. Coordinated Changes of Mitochondrial Biogenesis and Antioxidant Enzymes During Osteogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells 2008, 26, 960–968. [Google Scholar] [CrossRef]
- Tormos, K.V.; Anso, E.; Hamanaka, R.B.; Eisenbart, J.; Joseph, J.; Kalyanaraman, B.; Chandel, N.S. Mitochondrial Complex III ROS Regulate Adipocyte Differentiation. Cell Metab. 2011, 14, 537–544. [Google Scholar] [CrossRef]
- Zhang, Y.; Marsboom, G.; Toth, P.T.; Rehman, J. Mitochondrial Respiration Regulates Adipogenic Differentiation of Human Mesenchymal Stem Cells. PLoS ONE 2013, 8, e77077. [Google Scholar] [CrossRef]
- Spangenberg, L.; Shigunov, P.; Abud, A.P.R.; Cofré, A.R.; Stimamiglio, M.A.; Kuligovski, C.; Zych, J.; van Schittini, A.; Costa, A.D.T.; Rebelatto, C.K.; et al. Polysome Profiling Shows Extensive Posttranscriptional Regulation during Human Adipocyte Stem Cell Differentiation into Adipocytes. Stem Cell Res. 2013, 11, 902–912. [Google Scholar] [CrossRef]
- Drehmer, D.L.; de Aguiar, A.M.; Brandt, A.P.; Petiz, L.; Cadena, S.M.S.C.; Rebelatto, C.K.; Brofman, P.R.S.; Filipak Neto, F.; Dallagiovanna, B.; Abud, A.P.R. Metabolic Switches during the First Steps of Adipogenic Stem Cells Differentiation. Stem Cell Res. 2016, 17, 413–421. [Google Scholar] [CrossRef]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef]
- Farmer, S.R. Transcriptional Control of Adipocyte Formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.S.; Siersbæk, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome Proliferator-Activated Receptor γ and C/EBPα Synergistically Activate Key Metabolic Adipocyte Genes by Assisted Loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Morigny, P.; Boucher, J.; Arner, P.; Langin, D. Lipid and Glucose Metabolism in White Adipocytes: Pathways, Dysfunction and Therapeutics. Nat. Rev. Endocrinol. 2021, 17, 276–295. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Hatzivassiliou, G.; Sachdeva, U.M.; van Bui, T.; Cross, J.R.; Thompson, C.B. ATP-Citrate Lyase Links Cellular Metabolism to Histone Acetylation. Science 2009, 324, 1076–1080. [Google Scholar] [CrossRef]
- Houten, S.M.; Wanders, R.J.A. A General Introduction to the Biochemistry of Mitochondrial Fatty Acid Β-oxidation. J. Inherit. Metab. Dis. 2010, 33, 469–477. [Google Scholar] [CrossRef]
- Wallace, M.; Metallo, C.M. Tracing Insights into de Novo Lipogenesis in Liver and Adipose Tissues. Semin. Cell Dev. Biol. 2020, 108, 65–71. [Google Scholar] [CrossRef]
- Prentice, K.J.; Saksi, J.; Hotamisligil, G.S. Adipokine FABP4 Integrates Energy Stores and Counterregulatory Metabolic Responses. J. Lipid Res. 2019, 60, 734–740. [Google Scholar] [CrossRef]
- Vogel Hertzel, A.; Bernlohr, D.A. The Mammalian Fatty Acid-Binding Protein Multigene Family: Molecular and Genetic Insights into Function. Trends Endocrinol. Metab. 2000, 11, 175–180. [Google Scholar] [CrossRef]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 Attenuates PPARγ and Adipogenesis and Is Inversely Correlated With PPARγ in Adipose Tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef]
- Jenkins-Kruchten, A.E.; Bennaars-Eiden, A.; Ross, J.R.; Shen, W.-J.; Kraemer, F.B.; Bernlohr, D.A. Fatty Acid-Binding Protein-Hormone-Sensitive Lipase Interaction. J. Biol. Chem. 2003, 278, 47636–47643. [Google Scholar] [CrossRef]
- Gibler, P.; Gimble, J.; Hamel, K.; Rogers, E.; Henderson, M.; Wu, X.; Olesky, S.; Frazier, T. Human Adipose-Derived Stromal/Stem Cell Culture and Analysis Methods for Adipose Tissue Modeling In Vitro: A Systematic Review. Cells 2021, 10, 1378. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Park, J.; Yun, W.; Kang, P.J.; Son, D.; Jang, J.; Kim, I.Y.; You, S. Inhibitory Effect of Celastrol on Adipogenic Differentiation of Human Adipose-Derived Stem Cells. Biochem. Biophys. Res. Commun. 2018, 507, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Arzola-Paniagua, M.A.; García-Salgado López, E.R.; Calvo-Vargas, C.G.; Guevara-Cruz, M. Efficacy of an Orlistat-Resveratrol Combination for Weight Loss in Subjects with Obesity: A Randomized Controlled Trial. Obesity 2016, 24, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Konings, E.; Timmers, S.; van Boekschoten, M.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Müller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E.E. The Effects of 30 Days Resveratrol Supplementation on Adipose Tissue Morphology and Gene Expression Patterns in Obese Men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yu, S.; Han, H.; Yuan, Q.; Chen, J.; Xu, G. Resveratrol Inhibits Human Visceral Preadipocyte Proliferation and Differentiation in Vitro. Lipids 2019, 54, 679–686. [Google Scholar] [CrossRef]
- Park, A. Distinction of White, Beige and Brown Adipocytes Derived from Mesenchymal Stem Cells. World J. Stem Cells 2014, 6, 33. [Google Scholar] [CrossRef]
- Senamontree, S.; Lakthan, T.; Charoenpanich, P.; Chanchao, C.; Charoenpanich, A. Betulinic Acid Decreases Lipid Accumulation in Adipogenesis-Induced Human Mesenchymal Stem Cells with Upregulation of PGC-1α and UCP-1 and Post-Transcriptional Downregulation of Adiponectin and Leptin Secretion. PeerJ 2021, 9, e12321. [Google Scholar] [CrossRef]
- Huang, H.-J.; Lee, Y.-H.; Hsu, Y.-H.; Liao, C.-T.; Lin, Y.-F.; Chiu, H.-W. Current Strategies in Assessment of Nanotoxicity: Alternatives to In Vivo Animal Testing. Int. J. Mol. Sci. 2021, 22, 4216. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique. Med. J. Aust. 1960, 1, 500. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, C.; Jia, J.; Wang, Z.; Li, L.; Deng, X.; Cai, Z.; Yang, L.; Wang, D.; Ma, S.; et al. Regulatory Network of Metformin on Adipogenesis Determined by Combining High-Throughput Sequencing and GEO Database. Adipocyte 2022, 11, 56–68. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, L.; Li, S.; Zhang, W.; Ma, X.; Cheng, G.; Yang, W.; Zan, L. Identification of Potential Key Genes Associated with Adipogenesis through Integrated Analysis of Five Mouse Transcriptome Datasets. Int. J. Mol. Sci. 2018, 19, 3557. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, X.; Yan, R.; Jiang, H.; Wang, T.; Fan, L.; Wu, J.; Cao, J.; Li, W. Gene Expression Profiling of Human Bone Marrow-Derived Mesenchymal Stem Cells during Adipogenesis. Folia Histochem. Cytobiol. 2015, 54, 14–24. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, P.; Song, M.; Wang, Y.; Deng, S.; Hong, W.; Zhang, X.; Yu, B. Identification of Key Genes of Human Bone Marrow Stromal Cells Adipogenesis at an Early Stage. PeerJ 2020, 8, e9484. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, Y.-H.; Liu, L.; Yu, L.-L.; Li, J.-P.; Rao, J.; Hu, F.; Zhu, L.-J.; Bao, H.-H.; Cheng, X.-S. Bioinformatics Analysis of Candidate Genes and Potential Therapeutic Drugs Targeting Adipose Tissue in Obesity. Adipocyte 2022, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A Powerful Approach for Structure-Based Drug Discovery. Curr. Comput. Aided-Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.; Joshi, K.; Sasikala, K.; Alvala, M. Molecular Docking in Modern Drug Discovery: Principles and Recent Applications. In Drug Discovery and Development—New Advances; IntechOpen: London, UK, 2020. [Google Scholar]
- Alalaiwe, A.; Fang, J.-Y.; Lee, H.-J.; Chiu, C.-H.; Hsu, C.-Y. The Demethoxy Derivatives of Curcumin Exhibit Greater Differentiation Suppression in 3T3-L1 Adipocytes Than Curcumin: A Mechanistic Study of Adipogenesis and Molecular Docking. Biomolecules 2021, 11, 1025. [Google Scholar] [CrossRef]
- Bedi, O.; Aggarwal, S.; Trehanpati, N.; Ramakrishna, G.; Grewal, A.S.; Krishan, P. In Vitro Targeted Screening and Molecular Docking of Stilbene, Quinones, and Flavonoid on 3T3-L1 Pre-Adipocytes for Anti-Adipogenic Actions. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 2093–2106. [Google Scholar] [CrossRef]
- Mujwar, S.; Kumar, V. Computational Drug Repurposing Approach to Identify Potential Fatty Acid-Binding Protein-4 Inhibitors to Develop Novel Antiobesity Therapy. Assay Drug Dev. Technol. 2020, 18, 318–327. [Google Scholar] [CrossRef]
- Jourdan, J.-P.; Bureau, R.; Rochais, C.; Dallemagne, P. Drug Repositioning: A Brief Overview. J. Pharm. Pharmacol. 2020, 72, 1145–1151. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural Summaries of PDB Entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.A.; Nguyen, V.T.; Levi, B.; James, A.W. Current Methods of Adipogenic Differentiation of Mesenchymal Stem Cells. Stem Cells Dev. 2011, 20, 1793–1804. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Chen, X.-Y.; Zhang, Z.-J.; Kang, Y.; Liao, W.-M.; Yu, W.-H.; Xiang, A.P. Expression Patterns of Transcription Factor PPARγ and C/EBP Family Members during in Vitro Adipogenesis of Human Bone Marrow Mesenchymal Stem Cells. Cell Biol. Int. 2015, 39, 457–465. [Google Scholar] [CrossRef]
- Cignarelli, A.; Genchi, V.; Perrini, S.; Natalicchio, A.; Laviola, L.; Giorgino, F. Insulin and Insulin Receptors in Adipose Tissue Development. Int. J. Mol. Sci. 2019, 20, 759. [Google Scholar] [CrossRef]
- Bahmad, H.F.; Daouk, R.; Azar, J.; Sapudom, J.; Teo, J.C.M.; Abou-Kheir, W.; Al-Sayegh, M. Modeling Adipogenesis: Current and Future Perspective. Cells 2020, 9, 2326. [Google Scholar] [CrossRef]
- Poulos, S.P.; van Dodson, M.; Hausman, G.J. Cell Line Models for Differentiation: Preadipocytes and Adipocytes. Exp. Biol. Med. 2010, 235, 1185–1193. [Google Scholar] [CrossRef]
- Dai, R.; Wang, Z.; Samanipour, R.; Koo, K.; Kim, K. Adipose-Derived Stem Cells for Tissue Engineering and Regenerative Medicine Applications. Stem Cells Int. 2016, 2016, 6737345. [Google Scholar] [CrossRef]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for Effective Differentiation of 3T3-L1 Cells to Adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef]
- Zhao, X.; Hu, H.; Wang, C.; Bai, L.; Wang, Y.; Wang, W.; Wang, J. A Comparison of Methods for Effective Differentiation of the Frozen-Thawed 3T3-L1 Cells. Anal. Biochem. 2019, 568, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, B.; Xu, W.; Jin, X.; Wang, K.; Wang, H. The Inhibitory Effects of Juglanin on Adipogenesis in 3T3-L1 Adipocytes. Drug Des. Dev. Ther. 2020, 14, 5349–5357. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Kim, T.Y.; Kang, H.; Oh, J.; Park, J.W.; Kim, S.-C.; Kim, M.; Apostolidis, E.; Kim, Y.-C.; Kwon, Y.-I. Anti-Obesity and Anti-Adipogenic Effects of Chitosan Oligosaccharide (GO2KA1) in SD Rats and in 3T3-L1 Preadipocytes Models. Molecules 2021, 26, 331. [Google Scholar] [CrossRef] [PubMed]
- Ariemma, F.; D’Esposito, V.; Liguoro, D.; Oriente, F.; Cabaro, S.; Liotti, A.; Cimmino, I.; Longo, M.; Beguinot, F.; Formisano, P.; et al. Low-Dose Bisphenol-A Impairs Adipogenesis and Generates Dysfunctional 3T3-L1 Adipocytes. PLoS ONE 2016, 11, e0150762. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Shen, Q.; Wu, J.; Li, J.-X. Oleanolic Acid Derivative HA-20 Inhibits Adipogenesis in a Manner Involving PPARγ-FABP4/AP2 Pathway. J. Mol. Endocrinol. 2021, 66, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Benchamana, A.; Mori, H.; MacDougald, O.A.; Soodvilai, S. Regulation of Adipocyte Differentiation and Metabolism by Lansoprazole. Life Sci. 2019, 239, 116897. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Lian, Y.; Tang, J.; Chen, F.; Gao, H.; Zhou, Z.; Hou, N.; Yi, W. Identification of the Anti-Fungal Drug Fenticonazole Nitrate as a Novel PPARγ-Modulating Ligand with Good Therapeutic Index: Structure-Based Screening and Biological Validation. Pharmacol. Res. 2021, 173, 105860. [Google Scholar] [CrossRef] [PubMed]
- Klingelhutz, A.J.; Gourronc, F.A.; Chaly, A.; Wadkins, D.A.; Burand, A.J.; Markan, K.R.; Idiga, S.O.; Wu, M.; Potthoff, M.J.; Ankrum, J.A. Scaffold-Free Generation of Uniform Adipose Spheroids for Metabolism Research and Drug Discovery. Sci. Rep. 2018, 8, 523. [Google Scholar] [CrossRef]
- Abud, A.P.R.; Kuligovski, C.; Corrêa, N.C.R.; de Moraes, E.C.P.; Caruso, R.R.B.; Schuck, D.C.; Brohem, C.A.; Dallagiovanna, B.; de Aguiar, A.M. The Inhibition of Adipogenesis via an in Vitro Assay Can Reduce Animal Use by More Precisely Estimating the Starting Dose for the Acute Toxic Class Method. Toxicol. Lett. 2019, 311, 80–90. [Google Scholar] [CrossRef]
- Abud, A.P.R.; Paschoal, A.C.C.; Kuligovski, C.; Caruso, R.R.B.; Dallagiovanna, B.; de Aguiar, A.M. Using Inhibition of the Adipogenesis of Adipose-Derived Stem Cells in Vitro for Toxicity Prediction. MethodsX 2021, 8, 101515. [Google Scholar] [CrossRef]
- Ribeiro, A.L.; Bassai, L.W.; Robert, A.W.; Machado, T.N.; Bezerra, A.G.; da Silva Horinouchi, C.D.; de Aguiar, A.M. Bismuth-Based Nanoparticles Impair Adipogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Toxicol. In Vitro 2021, 77, 105248. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.C.; Cohenour, E.R.; Harnett, K.G.; Schuh, S.M. BPA, BPAF and TMBPF Alter Adipogenesis and Fat Accumulation in Human Mesenchymal Stem Cells, with Implications for Obesity. Int. J. Mol. Sci. 2021, 22, 5363. [Google Scholar] [CrossRef] [PubMed]
- Palomares, B.; Ruiz-Pino, F.; Navarrete, C.; Velasco, I.; Sánchez-Garrido, M.A.; Jimenez-Jimenez, C.; Pavicic, C.; Vazquez, M.J.; Appendino, G.; Bellido, M.L.; et al. VCE-004.8, A Multitarget Cannabinoquinone, Attenuates Adipogenesis and Prevents Diet-Induced Obesity. Sci. Rep. 2018, 8, 16092. [Google Scholar] [CrossRef] [PubMed]
- Sutjarit, N.; Thongon, N.; Weerachayaphorn, J.; Piyachaturawat, P.; Suksamrarn, A.; Suksen, K.; Papachristou, D.J.; Blair, H.C. Inhibition of Adipogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by a Phytoestrogen Diarylheptanoid from Curcuma Comosa. J. Agric. Food Chem. 2020, 68, 9993–10002. [Google Scholar] [CrossRef]
- Chang, R.C.; Thangavelu, C.S.; Joloya, E.M.; Kuo, A.; Li, Z.; Blumberg, B. Cannabidiol Promotes Adipogenesis of Human and Mouse Mesenchymal Stem Cells via PPARγ by Inducing Lipogenesis but Not Lipolysis. Biochem. Pharmacol. 2022, 197, 114910. [Google Scholar] [CrossRef] [PubMed]
- González-Casanova, J.E.; Cruz, S.L.P.; Vivas, M.C.; Rojas-Gómez, D.M. Influencia de Disruptores Endocrinos Medioambientales Sobre La Adipogénesis Resumen Influence of Endocrine Disrupting Chemicals on Adipogénesis. Arch. Venez. de Farmacol. y Ter. 2018, 37, 28–36. [Google Scholar]
- Mandl, M.; Viertler, H.P.; Hatzmann, F.M.; Brucker, C.; Großmann, S.; Waldegger, P.; Rauchenwald, T.; Mattesich, M.; Zwierzina, M.; Pierer, G.; et al. An Organoid Model Derived from Human Adipose Stem/Progenitor Cells to Study Adipose Tissue Physiology. Adipocyte 2022, 11, 164–174. [Google Scholar] [CrossRef]
- Audano, M.; Pedretti, S.; Caruso, D.; Crestani, M.; de Fabiani, E.; Mitro, N. Regulatory Mechanisms of the Early Phase of White Adipocyte Differentiation: An Overview. Cell. Mol. Life Sci. 2022, 79, 139. [Google Scholar] [CrossRef]
- Aprile, M.; Cataldi, S.; Perfetto, C.; Ambrosio, M.R.; Italiani, P.; Tatè, R.; Blüher, M.; Ciccodicola, A.; Costa, V. In Vitro-Generated Hypertrophic-Like Adipocytes Displaying PPARG Isoforms Unbalance Recapitulate Adipocyte Dysfunctions In Vivo. Cells 2020, 9, 1284. [Google Scholar] [CrossRef]
- Ramirez-Zacarias, J.L.; Castro-Muñozledo, F.; Kuri-Harcuch, W. Quantitation of Adipose Conversion and Triglycerides by Staining Intracytoplasmic Lipids with Oil Red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef]
- Mehlem, A.; Hagberg, C.E.; Muhl, L.; Eriksson, U.; Falkevall, A. Imaging of Neutral Lipids by Oil Red O for Analyzing the Metabolic Status in Health and Disease. Nat. Protoc. 2013, 8, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.D.; Greenspan, P. Application of Nile Red, a Fluorescent Hydrophobic Probe, for the Detection of Neutral Lipid Deposits in Tissue Sections: Comparison with Oil Red O. J. Histochem. Cytochem. 1985, 33, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile Red: A Selective Fluorescent Stain for Intracellular Lipid Droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Bonnefond, H.; Saint-Jean, B.; Rouxel, C.; Sciandra, A.; Bernard, O.; Cadoret, J.-P.; Bougaran, G. The Use of Fluorescent Nile Red and BODIPY for Lipid Measurement in Microalgae. Biotechnol. Biofuels 2015, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Katz, L.S.; Geras-Raaka, E.; Gershengorn, M.C. Heritability of Fat Accumulation in White Adipocytes. Am. J. Physiol.-Endocrinol. Metab. 2014, 307, E335–E344. [Google Scholar] [CrossRef]
- Govender, T.; Ramanna, L.; Rawat, I.; Bux, F. BODIPY Staining, an Alternative to the Nile Red Fluorescence Method for the Evaluation of Intracellular Lipids in Microalgae. Bioresour. Technol. 2012, 114, 507–511. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Strasser, B.C.; Scott, J.A.; Ross, G.M. Optimization of Staining Conditions for Microalgae with Three Lipophilic Dyes to Reduce Precipitation and Fluorescence Variability. Cytom. Part A 2012, 81A, 618–626. [Google Scholar] [CrossRef]
- Sieckmann, K.; Winnerling, N.; Huebecker, M.; Leyendecker, P.; Juliana Silva Ribeiro, D.; Gnad, T.; Pfeifer, A.; Wachten, D.; Hansen, J.N. AdipoQ—A Simple, Open-Source Software to Quantify Adipocyte Morphology and Function in Tissues and in Vitro. Mol. Biol. Cell 2022, 33, br22. [Google Scholar] [CrossRef]
- Shen, J.X.; Couchet, M.; Dufau, J.; de Castro Barbosa, T.; Ulbrich, M.H.; Helmstädter, M.; Kemas, A.M.; Zandi Shafagh, R.; Marques, M.; Hansen, J.B.; et al. 3D Adipose Tissue Culture Links the Organotypic Microenvironment to Improved Adipogenesis. Adv. Sci. 2021, 8, 2100106. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Turner, P.A.; Gurumurthy, B.; Bailey, J.L.; Elks, C.M.; Janorkar, A.V. Adipogenic Differentiation of Human Adipose-Derived Stem Cells Grown as Spheroids. Process Biochem. 2017, 59, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.D.; Pandey, R.; Tsancheva, V.S.; Candeo, A.; Botchway, S.W.; Allan, A.J.; Teboul, L.; Madi, K.; Babra, T.S.; Zolkiewski, L.A.K.; et al. The Development of a High Throughput Drug-Responsive Model of White Adipose Tissue Comprising Adipogenic 3T3-L1 Cells in a 3D Matrix. Biofabrication 2020, 12, 015018. [Google Scholar] [CrossRef]
- Pieters, V.M.; Rjaibi, S.T.; Singh, K.; Li, N.T.; Khan, S.T.; Nunes, S.S.; Dal Cin, A.; Gilbert, P.M.; McGuigan, A.P. A Three-Dimensional Human Adipocyte Model of Fatty Acid-Induced Obesity. Biofabrication 2022, 14, 045009. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.J.; Lee, J.H.; Park, S.B.; Na, Y.-J.; Jung, W.H.; Lee, H.; Kim, K.Y. Development of in Vitro Three-Dimensional Drug Screening System for Obesity-Related Metabolic Syndrome. J. Pharmacol. Sci. 2022, 148, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.; Frank, M.; Staehlke, S.; Peters, K. A Comparative Study on the Adipogenic Differentiation of Mesenchymal Stem/Stromal Cells in 2D and 3D Culture. Cells 2022, 11, 1313. [Google Scholar] [CrossRef]
- Hull, S.M.; Brunel, L.G.; Heilshorn, S.C. 3D Bioprinting of Cell-Laden Hydrogels for Improved Biological Functionality. Adv. Mater. 2022, 34, 2103691. [Google Scholar] [CrossRef]
- Schütz, K.; Placht, A.-M.; Paul, B.; Brüggemeier, S.; Gelinsky, M.; Lode, A. Three-Dimensional Plotting of a Cell-Laden Alginate/Methylcellulose Blend: Towards Biofabrication of Tissue Engineering Constructs with Clinically Relevant Dimensions. J. Tissue Eng. Regen. Med. 2017, 11, 1574–1587. [Google Scholar] [CrossRef]
- Duarte Campos, D.F.; Blaeser, A.; Korsten, A.; Neuss, S.; Jäkel, J.; Vogt, M.; Fischer, H. The Stiffness and Structure of Three-Dimensional Printed Hydrogels Direct the Differentiation of Mesenchymal Stromal Cells Toward Adipogenic and Osteogenic Lineages. Tissue Eng. Part A 2015, 21, 740–756. [Google Scholar] [CrossRef]
- Sakai, S.; Ohi, H.; Hotta, T.; Kamei, H.; Taya, M. Differentiation Potential of Human Adipose Stem Cells Bioprinted with Hyaluronic Acid/Gelatin-Based Bioink through Microextrusion and Visible Light-Initiated Crosslinking. Biopolymers 2018, 109, e23080. [Google Scholar] [CrossRef]
- Zhou, G.; Tian, A.; Yi, X.; Fan, L.; Shao, W.; Wu, H.; Sun, N. Study on a 3D-Bioprinted Tissue Model of Self-Assembled Nanopeptide Hydrogels Combined With Adipose-Derived Mesenchymal Stem Cells. Front. Bioeng. Biotechnol. 2021, 9, 663120. [Google Scholar] [CrossRef]
- Colle, J.; Blondeel, P.; de Bruyne, A.; Bochar, S.; Tytgat, L.; Vercruysse, C.; van Vlierberghe, S.; Dubruel, P.; Declercq, H. Bioprinting Predifferentiated Adipose-Derived Mesenchymal Stem Cell Spheroids with Methacrylated Gelatin Ink for Adipose Tissue Engineering. J. Mater. Sci. Mater. Med. 2020, 31, 36. [Google Scholar] [CrossRef] [PubMed]
- Benmeridja, L.; de Moor, L.; de Maere, E.; Vanlauwe, F.; Ryx, M.; Tytgat, L.; Vercruysse, C.; Dubruel, P.; van Vlierberghe, S.; Blondeel, P.; et al. High-throughput Fabrication of Vascularized Adipose Microtissues for 3D Bioprinting. J. Tissue Eng. Regen. Med. 2020, 14, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Horder, H.; Guaza Lasheras, M.; Grummel, N.; Nadernezhad, A.; Herbig, J.; Ergün, S.; Teßmar, J.; Groll, J.; Fabry, B.; Bauer-Kreisel, P.; et al. Bioprinting and Differentiation of Adipose-Derived Stromal Cell Spheroids for a 3D Breast Cancer-Adipose Tissue Model. Cells 2021, 10, 803. [Google Scholar] [CrossRef] [PubMed]
- Pati, F.; Ha, D.-H.; Jang, J.; Han, H.H.; Rhie, J.-W.; Cho, D.-W. Biomimetic 3D Tissue Printing for Soft Tissue Regeneration. Biomaterials 2015, 62, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Gruene, M.; Pflaum, M.; Deiwick, A.; Koch, L.; Schlie, S.; Unger, C.; Wilhelmi, M.; Haverich, A.; Chichkov, B.N. Adipogenic Differentiation of Laser-Printed 3D Tissue Grafts Consisting of Human Adipose-Derived Stem Cells. Biofabrication 2011, 3, 015005. [Google Scholar] [CrossRef]
- Qi, D.; Wu, S.; Kuss, M.A.; Shi, W.; Chung, S.; Deegan, P.T.; Kamenskiy, A.; He, Y.; Duan, B. Mechanically Robust Cryogels with Injectability and Bioprinting Supportability for Adipose Tissue Engineering. Acta Biomater. 2018, 74, 131–142. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.W.; Chen, H.Y.; Wang, Z.X.; Li, X.D.; Xu, R.X.; Xu, T. 3D Bioprinted Innervation ADMSC Self-Clustering Culture Model Constructs for in vitro Fat Metabolism Research: A Preliminary Study of ADMSC and Neural Progenitor Cell Co-Culture Model Construct Fabrication and Characterization. Mater. Today Chem. 2022, 26, 101092. [Google Scholar] [CrossRef]
- Ingber, D.E. Human Organs-on-Chips for Disease Modelling, Drug Development and Personalized Medicine. Nat. Rev. Genet. 2022, 23, 467–491. [Google Scholar] [CrossRef]
- Hargrove-Grimes, P.; Low, L.A.; Tagle, D.A. Microphysiological Systems: Stakeholder Challenges to Adoption in Drug Development. Cells Tissues Organs 2022, 211, 269–281. [Google Scholar] [CrossRef]
- Pun, S.; Haney, L.C.; Barrile, R. Modelling Human Physiology On-Chip: Historical Perspectives and Future Directions. Micromachines 2021, 12, 1250. [Google Scholar] [CrossRef]
- Frazier, T.P.; Hamel, K.; Wu, X.; Rogers, E.; Lassiter, H.; Robinson, J.; Mohiuddin, O.; Henderson, M.; Gimble, J.M. Adipose-Derived Cells: Building Blocks of Three-Dimensional Microphysiological Systems. Biomater. Transl. 2021, 4, 301–306. [Google Scholar] [CrossRef]
- McCarthy, M.; Brown, T.; Alarcon, A.; Williams, C.; Wu, X.; Abbott, R.D.; Gimble, J.; Frazier, T. Fat-On-A-Chip Models for Research and Discovery in Obesity and Its Metabolic Comorbidities. Tissue Eng. Part B Rev. 2020, 26, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Kongsuphol, P.; Gupta, S.; Liu, Y.; Bhuvanendran Nair Gourikutty, S.; Biswas, S.K.; Ramadan, Q. In Vitro Micro-Physiological Model of the Inflamed Human Adipose Tissue for Immune-Metabolic Analysis in Type II Diabetes. Sci. Rep. 2019, 9, 4887. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, Q.; Gourikutty, S.B.N.; Zhang, Q. OOCHIP: Compartmentalized Microfluidic Perfusion System with Porous Barriers for Enhanced Cell–Cell Crosstalk in Organ-on-a-Chip. Micromachines 2020, 11, 565. [Google Scholar] [CrossRef]
- Liu, Y.; Kongsuphol, P.; Chiam, S.Y.; Zhang, Q.X.; Gourikutty, S.B.N.; Saha, S.; Biswas, S.K.; Ramadan, Q. Adipose-on-a-Chip: A Dynamic Microphysiological in vitro Model of the Human Adipose for Immune-Metabolic Analysis in Type II Diabetes. Lab Chip 2019, 19, 241–253. [Google Scholar] [CrossRef]
- O’Donnell, B.; Al-Ghadban, S.; Ives, C.; L’Ecuyer, M.; Monjure, T.; Romero-Lopez, M.; Li, Z.; Goodman, S.; Lin, H.; Tuan, R.; et al. Adipose Tissue-Derived Stem Cells Retain Their Adipocyte Differentiation Potential in Three-Dimensional Hydrogels and Bioreactors. Biomolecules 2020, 10, 1070. [Google Scholar] [CrossRef]
- Wu, X.; Schneider, N.; Platen, A.; Mitra, I.; Blazek, M.; Zengerle, R.; Schüle, R.; Meier, M. In Situ Characterization of the MTORC1 during Adipogenesis of Human Adult Stem Cells on Chip. Proc. Natl. Acad. Sci. USA 2016, 113, E4143–E4150. [Google Scholar] [CrossRef]
- Yang, F.; Carmona, A.; Stojkova, K.; Garcia Huitron, E.I.; Goddi, A.; Bhushan, A.; Cohen, R.N.; Brey, E.M. A 3D Human Adipose Tissue Model within a Microfluidic Device. Lab Chip 2021, 21, 435–446. [Google Scholar] [CrossRef]
- Yang, F.; Cohen, R.N.; Brey, E.M. Optimization of Co-Culture Conditions for a Human Vascularized Adipose Tissue Model. Bioengineering 2020, 7, 114. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, A.; Qi, W. The Potential to Fight Obesity with Adipogenesis Modulating Compounds. Int. J. Mol. Sci. 2022, 23, 2299. [Google Scholar] [CrossRef]
- Vishvanath, L.; Gupta, R.K. Contribution of Adipogenesis to Healthy Adipose Tissue Expansion in Obesity. J. Clin. Investig. 2019, 129, 4022–4031. [Google Scholar] [CrossRef] [PubMed]
- Paul Gleeson, M.; Hersey, A.; Hannongbua, S. In-Silico ADME Models: A General Assessment of Their Utility in Drug Discovery Applications. Curr. Top. Med. Chem. 2011, 11, 358–381. [Google Scholar] [CrossRef] [PubMed]
- Bassan, A.; Alves, V.M.; Amberg, A.; Anger, L.T.; Beilke, L.; Bender, A.; Bernal, A.; Cronin, M.T.D.; Hsieh, J.-H.; Johnson, C.; et al. In Silico Approaches in Organ Toxicity Hazard Assessment: Current Status and Future Needs for Predicting Heart, Kidney and Lung Toxicities. Comput. Toxicol. 2021, 20, 100188. [Google Scholar] [CrossRef] [PubMed]
- Navien, T.N.; Thevendran, R.; Hamdani, H.Y.; Tang, T.-H.; Citartan, M. In Silico Molecular Docking in DNA Aptamer Development. Biochimie 2021, 180, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.M.D.; Acosta, A. Precision Medicine and Obesity. Gastroenterol. Clin. N. Am. 2021, 50, 127–139. [Google Scholar] [CrossRef]
- Barro, L.; Nebie, O.; Chen, M.-S.; Wu, Y.-W.; Koh, M.B.; Knutson, F.; Watanabe, N.; Takahara, M.; Burnouf, T. Nanofiltration of Growth Media Supplemented with Human Platelet Lysates for Pathogen-Safe Xeno-Free Expansion of Mesenchymal Stromal Cells. Cytotherapy 2020, 22, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Bartness, T.J.; Ryu, V. Neural Control of White, Beige and Brown Adipocytes. Int. J. Obes. Suppl. 2015, 5, S35–S39. [Google Scholar] [CrossRef]
- Chan, H.C.S.; Shan, H.; Dahoun, T.; Vogel, H.; Yuan, S. Advancing Drug Discovery via Artificial Intelligence. Trends Pharmacol. Sci. 2019, 40, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhang, M.; Tong, M.; Yang, L.; Pang, L.; Chen, L.; Xu, G.; Chi, X.; Hong, Q.; Ni, Y.; et al. MiR-148a Is Associated with Obesity and Modulates Adipocyte Differentiation of Mesenchymal Stem Cells through Wnt Signaling. Sci. Rep. 2015, 5, 9930. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Gilani, A.; Yi, Q.; Tang, L. MicroRNAs as Mediators of Adipose Thermogenesis and Potential Therapeutic Targets for Obesity. Biology 2022, 11, 1657. [Google Scholar] [CrossRef]
- Herrero-Aguayo, V.; Jiménez-Vacas, J.M.; Sáez-Martínez, P.; Gómez-Gómez, E.; López-Cánovas, J.L.; Garrido-Sánchez, L.; Herrera-Martínez, A.D.; García-Bermejo, L.; Macías-González, M.; López-Miranda, J.; et al. Influence of Obesity in the MiRNome: MiR-4454, a Key Regulator of Insulin Response Via Splicing Modulation in Prostate. J. Clin. Endocrinol. Metab. 2021, 106, e469–e484. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Peluso, G.; Galderisi, U.; di Bernardo, G. Long Non-Coding RNAs in Regulation of Adipogenesis and Adipose Tissue Function. eLife 2020, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pamplona, J.H.; Zoehler, B.; Shigunov, P.; Barisón, M.J.; Severo, V.R.; Erich, N.M.; Ribeiro, A.L.; Horinouchi, C.D.d.S.; Suzukawa, A.A.; Robert, A.W.; et al. Alternative Methods as Tools for Obesity Research: In Vitro and In Silico Approaches. Life 2023, 13, 108. https://doi.org/10.3390/life13010108
Pamplona JH, Zoehler B, Shigunov P, Barisón MJ, Severo VR, Erich NM, Ribeiro AL, Horinouchi CDdS, Suzukawa AA, Robert AW, et al. Alternative Methods as Tools for Obesity Research: In Vitro and In Silico Approaches. Life. 2023; 13(1):108. https://doi.org/10.3390/life13010108
Chicago/Turabian StylePamplona, Juliana Helena, Bernardo Zoehler, Patrícia Shigunov, María Julia Barisón, Vanessa Rossini Severo, Natalie Mayara Erich, Annanda Lyra Ribeiro, Cintia Delai da Silva Horinouchi, Andréia Akemi Suzukawa, Anny Waloski Robert, and et al. 2023. "Alternative Methods as Tools for Obesity Research: In Vitro and In Silico Approaches" Life 13, no. 1: 108. https://doi.org/10.3390/life13010108
APA StylePamplona, J. H., Zoehler, B., Shigunov, P., Barisón, M. J., Severo, V. R., Erich, N. M., Ribeiro, A. L., Horinouchi, C. D. d. S., Suzukawa, A. A., Robert, A. W., Abud, A. P. R., & Aguiar, A. M. d. (2023). Alternative Methods as Tools for Obesity Research: In Vitro and In Silico Approaches. Life, 13(1), 108. https://doi.org/10.3390/life13010108