

Glycine Effect on the Expression Profile of Orphan Receptors GPR21, GPR26, GPR39, GPR82 and GPR6 in a Model of Inflammation in 3T3-L1 Cells

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Experimental Design
2.3. RNA Extraction and cDNA Synthesis
2.4. Quantitative Real-Time RT-qPCR
- GPR21, GPR26, GPR39, GPR82, GPR6, TNF-α, adipoQ and 36B4 (Rplp0) as the housekeeping gene were analysed by the Nano LightCycler System (Roche Diagnostics) (Table 1). RT-qPCR was performed using the FastStart Essential DNA Probes Master Mix (Roche Applied Science, Mannheim, Germany). Probes from Universal Probe Library were used (Table 1) (Roche Applied Science, Mannheim, Germany), with 0.3 µL of each primer (Oligo T4, Irapuato, Mexico) and 1000 ng of RNA. The reaction was performed in three steps. Step 1: 10 min preheating at 95 °C; step 2: 45 cycles of 15 s at 90 °C, 30 s at 60 °C and 15 s at 72 °C; step 3: cooling for 300 s at 40 °C. Relative changes in gene expression were determined using the 2−ΔΔCt method [56].
2.5. Principal Component Analysis (PCA) of Existing Orphan Receptors Involved in Inflammatory and Anti-Inflammatory Processes
2.6. Statistical Analysis
3. Results
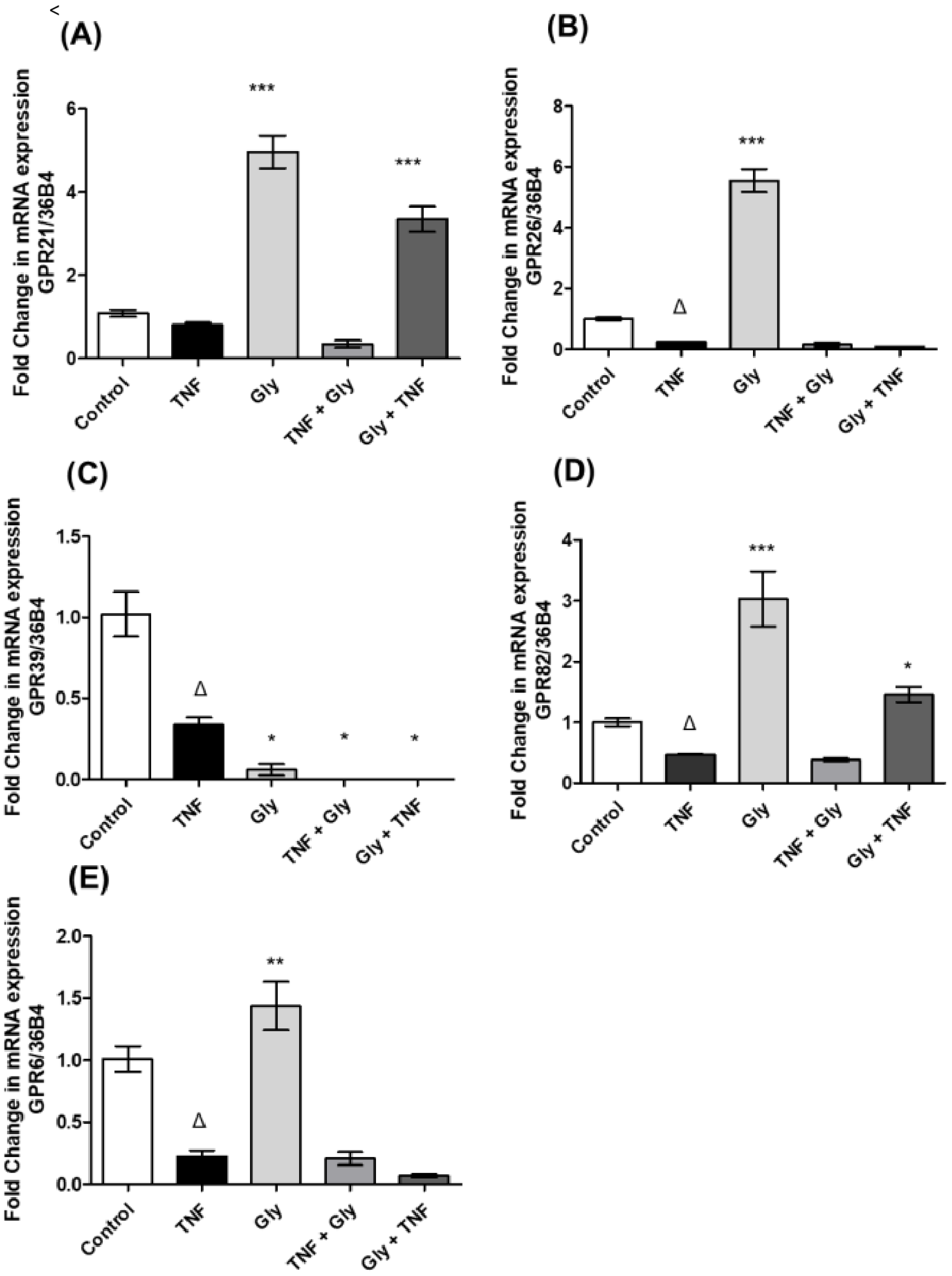
3.1. Impact of Inflammatory and Anti-Inflammatory Environments on GPR21, GPR26, GPR39, GPR82 and GPR6 Gene Expression
3.2. Pre- and Post-Treatment with Glycine on Orphan Receptor Expression
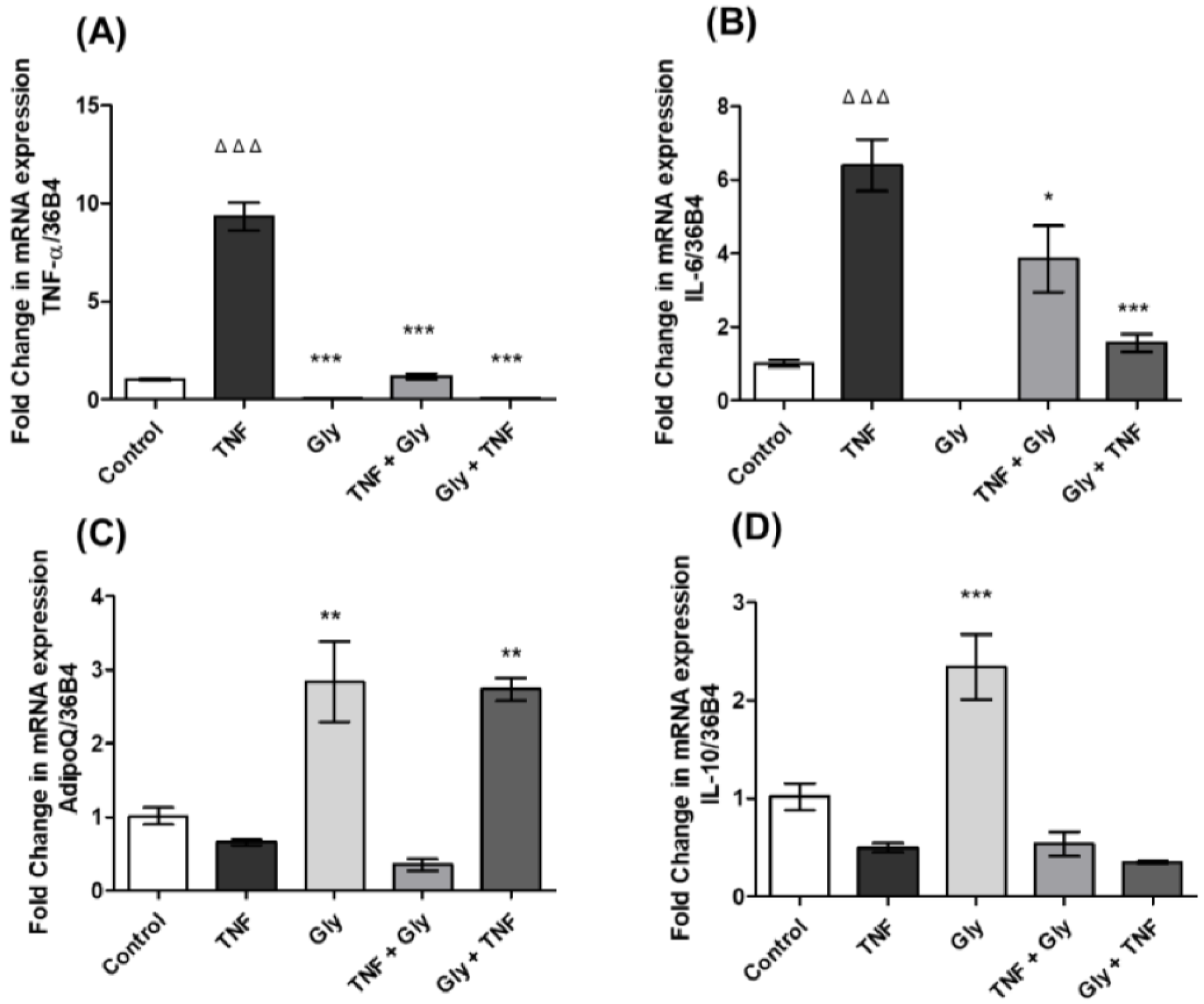
3.3. Modulation of TNF-α and IL-6 Gene Expression by Glycine in Differentiated 3T3-L1 Adipocytes
3.4. AdipoQ and IL-10 Gene Expression Changes by Glycine in 3T3-L1 Adipocytes
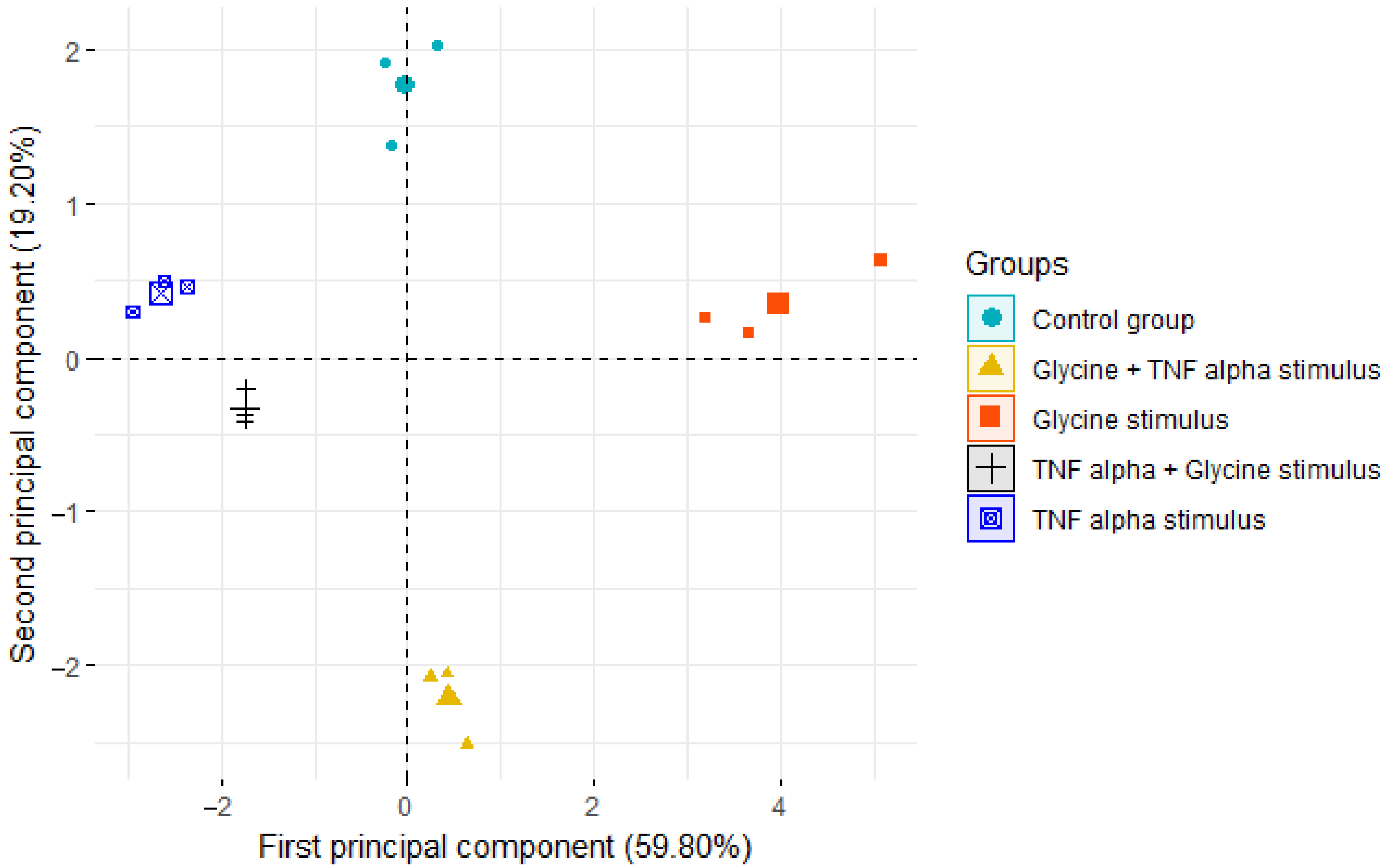
3.5. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calder, P.C.; Ahluwalia, N.; Albers, R.; Bosco, N.; Bourdet-Sicard, R.; Haller, D.; Holgate, S.T.; Jönsson, L.S.; Latulippe, M.E.; Marcos, A.; et al. A Consideration of Biomarkers to be Used for Evaluation of Inflammation in Human Nutritional Studies. Br. J. Nutr. 2013, 109, S1–S34. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Harford, K.A.; Reynolds, C.M.; McGillicuddy, F.; Roche, H.M. Fats, inflammation and insulin resistance: Insights to the role of macrophage and T-cell accumulation in adipose tissue. Proc. Nutr. Soc. 2011, 70, 408–417. [Google Scholar] [CrossRef]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic Switching of Adipose Tissue Macrophages With Obesity Is Generated by Spatiotemporal Differences in Macrophage Subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef]
- Ukita, M.; Yamaguchi, T.; Ohata, N.; Tamura, M. Sclerostin Enhances Adipocyte Differentiation in 3T3-L1 Cells. J. Cell. Biochem. 2016, 117, 1419–1428. [Google Scholar] [CrossRef]
- Hadrich, F.; Sayadi, S. La apiggetrina inhibe la adipogénesis en las células 3T3-L1 mediante la regulación negativa de PPARγ y CEBP-α. Lípidos Salud Dis. 2018, 17, 95. [Google Scholar] [CrossRef]
- McCarty, M.F.; Lerner, A. Perspective: Prospects for nutraceutical support of intestinal barrier function. Adv. Nutr. 2021, 12, 316–324. [Google Scholar] [CrossRef]
- Lewis, R.M.; Godfrey, K.M.; Jackson, A.A.; Cameron, I.T.; Hanson, M.A. Low Serine Hydroxymethyltransferase Activity in the Human Placenta Has Important Implications for Fetal Glycine Supply. J. Clin. Endocrinol. Metab. 2005, 90, 1594–1598. [Google Scholar] [CrossRef]
- Romero-Nava, R.; Alarcón-Aguilar, F.J.; Giacoman-Martínez, A.; Blancas-Flores, G.; Aguayo-Cerón, K.A.; Ballinas-Verdugo, M.A.; Sánchez-Muñoz, F.; Huang, F.; Villafaña-Rauda, S.; Almanza-Pérez, J.C. Glycine is a competitive antagonist of the TNF receptor mediating the expression of inflammatory cytokines in 3T3-L1 adipocytes. Inflamm. Res. 2021, 70, 605–618. [Google Scholar] [CrossRef]
- Matilla, B.; Mauriz, J.L.; Culebras, J.M.; González-Gallego, J.; González, P. Glycine: A cell-protecting anti-oxidant nutrient. Nutr. Hosp. 2002, 17, 2–9. [Google Scholar] [PubMed]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine metabolism in animals and humans: Implications for nutrition and health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Reyes López, Y.; Pérez-Torres, I.; Zúñiga-Muñoz, A.; Guarner Lans, V.; Díaz-Díaz, E.; Soria Castro, E.; Velázquez, E.R. Effect of glycine on adipocyte hypertrophy in a metabolic syndrome Rat Model. Curr. Drug Deliv. 2016, 13, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Alarcon-Aguilar, F.J.; Almanza-Perez, J.; Blancas, G.; Angeles, S.; Garcia-Macedo, R.; Roman, R.; Cruz, M. Glycine regulates the production of pro-inflammatory cytokines in lean and monosodium glutamate-obese mice. Eur. J. Pharmacol. 2008, 599, 152–158. [Google Scholar] [CrossRef]
- Almanza-Perez, J.C.; Alarcon-Aguilar, F.J.; Blancas-Flores, G.; Campos-Sepulveda, A.E.; Roman-Ramos, R.; Garcia-Macedo, R.; Cruz, M. Glycine regulates inflammatory markers modifying the energetic balance through PPAR and UCP-2. Biomed. Pharmacother. 2010, 64, 534–540. [Google Scholar] [CrossRef]
- Contreras-Nuñez, E.; Blancas-Flores, G.; Cruz, M.; Almanza-Perez, J.C.; Gomez-Zamudio, J.H.; Ventura-Gallegos, J.L.; Zentella-Dehesa, A.; Roberto-Lazzarini, R.-R.; Alarcon-Aguilar, F.J. Participation of the IKK-α/β complex in the inhibition of the TNF-α/NF-κB pathway by glycine: Possible involvement of a membrane receptor specific to adipocytes. Biomed. Pharmacother. 2018, 102, 120–131. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schioth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Sriram, K.; Insel, P.A. G Protein-Coupled Receptors as Targets for Approved Drugs: How Many Targets and How Many Drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar] [CrossRef]
- Watkins, L.R.; Orlandi, C. Orphan G protein coupled receptors in affective disorders. Genes 2020, 11, 694. [Google Scholar] [CrossRef]
- Fang, Y.; Kenakin, T.; Liu, C. Editorial: Orphan GPCRs As Emerging Drug Targets. Front. Pharmacol. 2015, 6, 295. [Google Scholar] [CrossRef]
- Ngo, T.; Coleman, J.L.J.; Smith, N.J. Using Constitutive Activity to Define Appropriate High-Throughput Screening Assays for Orphan G Protein-Coupled Receptors. Methods Mol. Biol. 2015, 1272, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.R.; Orlandi, C. In vitro profiling of orphan G protein coupled receptor (GPCR) constitutive activity. Br. J. Pharmacol. 2021, 178, 2963–2975. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Damian, M.; Marie, J.; Leyris, J.P.; Fehrentz, J.A.; Verdie, P.; Martinez, J.; Banères, J.-L.; Mary, S. High constitutive activity is an intrinsic feature of ghrelin receptor protein: A study with a functional monomeric GHS-R1a receptor reconstituted in lipid discs. J. Biol. Chem. 2012, 287, 3630–3641. [Google Scholar] [CrossRef]
- Corder, G.; Doolen, S.; Donahue, R.R.; Winter, M.K.; Jutras, B.L.; He, Y.; Hu, X.; Wieskopf, J.S.; Mogil, J.S.; Storm, D.R.; et al. Constitutive mu-opioid receptor activity leads to long-term endogenous analgesia and dependence. Science 2013, 341, 1394–1399. [Google Scholar] [CrossRef]
- Wacker, D.; Stevens, R.C.; Roth, B.L. How Ligands Illuminate GPCR Molecular Pharmacology. Cell 2017, 170, 414–427. [Google Scholar] [CrossRef]
- Leonard, S.; Kinsella, G.K.; Benetti, E.; Findlay, J.B.C. Regulating the effects of GPR21, a novel target for type 2 diabetes. Sci. Rep. 2016, 6, 27002. [Google Scholar] [CrossRef]
- Kumar, A.; Bharti, S.K.; Kumar, A. Therapeutic molecules against type 2 diabetes: What we have and what are we expecting? Pharmacol. Rep. 2017, 69, 959–970. [Google Scholar] [CrossRef]
- Romero-Nava, R.; García, N.; Aguayo-Cerón, K.A.; Sánchez Muñoz, F.; Huang, F.; Hong, E.; Villafaña, S. Modifications in GPR21 and GPR82 genes expression as a consequence of metabolic syndrome etiology. J. Recept. Signal Transduct Res. 2021, 41, 38–44. [Google Scholar] [CrossRef]
- Osborn, O.; Oh, D.Y.; McNelis, J.; Sanchez-Alavez, M.; Talukdar, S.; Lu, M.; Li, P.; Thiede, L.; Morinaga, H.; Kim, J.J.; et al. G protein-coupled receptor 21 deletion improves insulin sensitivity in diet-induced obese mice. J. Clin. Investig. 2012, 122, 2444–2453. [Google Scholar] [CrossRef]
- Chung, S.; Funakoshi, T.; Civelli, O. Orphan GPCR research. Br. J. Pharmacol. 2008, 153 (Suppl. 1), S339–S346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-L.; Wang, J.-J.; Liu, Y.; Lu, X.-B.; Kuang, Y.; Wan, Y.-H.; Chen, Y.; Yan, H.-M.; Fei, J.; Wang, Z.-G. GPR26-deficient mice display increased anxiety- and depression-like behaviors accompanied by reduced phosphorylated cyclic AMP responsive element-binding protein level in central amygdala. Neuroscience 2011, 196, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.G.; Nawoschik, S.P.; Sreekumar, K.; Uveges, A.J.; Tseng, E.; Zhang, L.; Johnson, J.; He, L.; Paulsen, J.E.; Bates, B.; et al. Tissue distribution and functional analyses of the constitutively active orphan G protein coupled receptors, GPR26 and GPR78. Biochim. Biophys. Acta 2007, 1770, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Wang, S.; Li, W.-D.; Li, D.; Zhao, H.; Price, R.A. Interacting Genetic Loci on Chromosomes 20 and 10 Influence Extreme Human Obesity. Am. J. Hum. Genet. 2003, 72, 115–124. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Romero-Nava, R.; Zhou, D.-S.; García, N.; Ruiz-Hernández, A.; Si, Y.-C.; Sánchez-Muñoz, F.; Huang, F.; Hong, E.; Villafaña, S. Evidence of alterations in the expression of orphan receptors GPR26 and GPR39 due to the etiology of the metabolic syndrome. J. Recept. Signal Transduct. Res. 2017, 37, 422–429. [Google Scholar] [CrossRef]
- McKee, K.K.; Tan, C.P.; Palyha, O.C.; Liub, J.; Feighner, S.D.; Hreniuk, D.L.; Smith, R.G.; Howard, A.D.; Van Der Ploeg, L.H. Cloning and Characterization of Two Human G Protein-Coupled Receptor Genes (GPR38 and GPR39) Related to the Growth Hormone Secretagogue and Neurotensin Receptors. Genomics 1997, 46, 426–434. [Google Scholar] [CrossRef]
- Lauwers, E.; Landuyt, B.; Arckens, L.; Schoofs, L.; Luyten, W. Obestatin does not activate orphan G protein-coupled receptor GPR39. Biochem. Biophys. Res. Commun. 2006, 351, 21–25. [Google Scholar] [CrossRef]
- Holst, B.; Egerod, K.L.; Schild, E.; Vickers, S.P.; Cheetham, S.; Gerlach, L.-O.; Storjohann, L.; Stidsen, C.E.; Jones, R.; Beck-Sickinger, A.G.; et al. GPR39 Signaling Is Stimulated by Zinc Ions But Not by Obestatin. Endocrinology 2007, 148, 13–20. [Google Scholar] [CrossRef]
- Laitakari, A.; Liu, L.; Frimurer, T.M.; Holst, B. The Zinc-Sensing Receptor GPR39 in Physiology and as a Pharmacological Target. Int. J. Mol. Sci. 2021, 22, 3872. [Google Scholar] [CrossRef]
- Sharir, H.; Hershfinkel, M. The extracellular zinc-sensing receptor mediates intercellular communication by inducing ATP release. Biochem. Biophys. Res. Commun. 2005, 332, 845–852. [Google Scholar] [CrossRef]
- Dong, X.; Tang, S.; Zhang, W.; Gao, W.; Chen, Y. GPR39 activates proliferation and differentiation of porcine intramuscular preadipocytes through targeting the PI3K/AKT cell signaling pathway. J. Recept. Signal Transduct. Res. 2016, 36, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Sekler, I.; Hershfinkel, M. The zinc sensing receptor, ZnR/GPR39, controls proliferation and differentiation of colonocytes and thereby tight junction formation in the colon. Cell Death Dis. 2014, 5, e1307. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Ntoupa, P.-S.A.; Spiliopoulou, C.A.; Stefanidou, M.E. Recent aspects of the effects of zinc on human health. Arch. Toxicol. 2020, 94, 1443–1460. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Sun, W.; Zhang, N.; Zhao, C.; Yan, X. The protective effects of the GPR39 agonist TC-G 1008 against TNF-α-induced inflammation in human fibroblast-like synoviocytes (FLSs). Eur. J. Pharmacol. 2019, 865, 172663. [Google Scholar] [CrossRef]
- Shan, W.; Qi, J.; Li, C.; Nie, X. Agonism of GPR39 displays protective effects against advanced glycation end-product (AGE)-induced degradation of extracellular matrix in human SW1353 cells. Arch. Biochem. Biophys. 2019, 677, 108164. [Google Scholar] [CrossRef]
- Muneoka, S.; Goto, M.; Kadoshima-Yamaoka, K.; Kamei, R.; Terakawa, M.; Tomimori, Y. G protein-coupled receptor 39 plays an anti-inflammatory role by enhancing IL-10 production from macrophages under inflammatory conditions. Eur. J. Pharmacol. 2018, 834, 240–245. [Google Scholar] [CrossRef]
- Southan, C.; Sharman, J.; Benson, H.E.; Faccenda, E.; Pawson, A.J.; Alexander, S.; Buneman, O.P.; Davenport, A.P.; McGrath, J.C.; Peters, J.A.; et al. The IUPHAR/BPS Guide to PHARMACOLOGY in 2016: Towards curated quantitative interactions between 1300 protein targets and 6000 ligands. Nucleic Acids Res. 2016, 44, D1054–D1068. [Google Scholar] [CrossRef]
- Engel, K.M.Y.; Schröck, K.; Teupser, D.; Holdt, L.M.; Tönjes, A.; Kern, M.; Dietrich, K.; Kovacs, P.; Krügel, U.; Scheidt, H.A.; et al. Reduced Food Intake and Body Weight in Mice Deficient for the G Protein-Coupled Receptor GPR82. PLoS ONE 2011, 6, e29400. [Google Scholar] [CrossRef]
- Romero-Nava, R.; Aguayo-Cerón, K.A.; Ruiz-Hernández, A.; Huang, F.; Hong, E.; Aguilera-Mendez, A.; Rauda, S.V. Silencing of GPR82 with Interference RNA Improved Metabolic Profiles in Rats with High Fructose Intake. J. Vasc. Res. 2020, 57, 1–7. [Google Scholar] [CrossRef]
- Prasad, B.M.; Hollins, B.; Lambert, N.A. Methods to Detect Cell Surface Expression and Constitutive Activity of GPR6. Methods Enzymol. 2010, 484, 179–195. [Google Scholar] [CrossRef]
- Isawi, I.H.; Morales, P.; Sotudeh, N.; Hurst, D.P.; Lynch, D.L.; Reggio, P.H. GPR6 Structural Insights: Homology Model Construction and Docking Studies. Molecules 2020, 25, 725. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.H.; Young, W.S.; Brownstein, M.J.; Bonner, T.I. Molecular cloning of a novel candidate G protein-coupled receptor from rat brain. FEBS Lett. 1994, 351, 375–379. [Google Scholar] [CrossRef]
- Morales, P.; Isawi, I.; Reggio, P.H. Towards a better understanding of the cannabinoid-related orphan receptors GPR3, GPR6, and GPR12. Drug Metab. Rev. 2018, 50, 74–93. [Google Scholar] [CrossRef] [PubMed]
- Laun, A.S.; Song, Z.-H. GPR3 and GPR6, novel molecular targets for cannabidiol. Biochem. Biophys. Res. Commun. 2017, 490, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Beeley, N.R.; Behan, D.P.; Chalmers, D.T.; Menzaghi, F.; Strah-Pleynet, S.; Arena Pharmaceuticals Inc. Small Molecule Modulators of G Protein-Coupled Receptor Six. U.S. Patent 6,420,563, 30 August 2001. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Sánchez-Solana, B.; Laborda, J.; Baladrón, V. Mouse Resistin Modulates Adipogenesis and Glucose Uptake in 3T3-L1 Preadipocytes Through the ROR1 Receptor. Mol. Endocrinol. 2012, 26, 110–127. [Google Scholar] [CrossRef]
- Zhang, J.V.; Jahr, H.; Luo, C.-W.; Klein, C.; Van Kolen, K.; Donck, L.V.; De, A.; Baart, E.; Li, J.; Moechars, D.; et al. Obestatin Induction of Early-Response Gene Expression in Gastrointestinal and Adipose Tissues and the Mediatory Role of G Protein-Coupled Receptor, GPR39. Mol. Endocrinol. 2008, 22, 1464–1475. [Google Scholar] [CrossRef]
- Gurriarán-Rodríguez, U.; Al-Massadi, O.; Crujeiras, A.B.; Mosteiro, C.S.; Amil-Diz, M.; Beiroa, D.; Nogueiras, R.; Seoane, L.M.; Gallego, R.; Pazos, Y.; et al. Preproghrelin expression is a key target for insulin action on adipogenesis. J. Endocrinol. 2011, 210, R1–R7. [Google Scholar] [CrossRef]
- Lee, J.-E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and Epigenomic Regulation of Adipogenesis. Mol. Cell. Biol. 2019, 39, e00601-18. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Li, B.; Zhang, D. Activation of G-protein-coupled bile acid receptor Gpbar1 (TGR5) inhibits degradation of type II collagen and aggrecan in human chondrocytes. Eur. J. Pharmacol. 2019, 856, 172387. [Google Scholar] [CrossRef] [PubMed]
- Spittler, A.; Reissner, C.M.; Oehler, R.; Gornikiewicz, A.; Gruenberger, T.; Manhart, N.; Brodowicz, T.; Mittlboeck, M.; Boltz-Nitulescu, G.; Roth, E. Immunomodulatory effects of glycine on LPS-treated monocytes: Reduced TNF-a production and accelerated IL-10 expression. FASEB J. 1999, 13, 563–571. [Google Scholar] [CrossRef]
- Garcia-Macedo, R.; Sanchez-Muñoz, F.; Almanza-Perez, J.C.; Duran-Reyes, G.; Alarcon-Aguilar, F.; Cruz, M. Glycine increases mRNA adiponectin and diminishes pro-inflammatory adipokines expression in 3T3-L1 cells. Eur. J. Pharmacol. 2008, 587, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Blancas-Flores, G.; Alarcón-Aguilar, F.J.; García-Macedo, R.; Almanza-Pérez, J.C.; Flores-Sáenz, J.L.; Román-Ramos, R.; Ventura-Gallegos, J.L.; Kumate, J.; Zentella-Dehesa, A.; Cruz, M. Glycine suppresses TNF-alpha-induced activation of NF-κB in differentiated 3T3-L1 adipocytes. Eur. J. Pharmacol. 2012, 689, 270–277. [Google Scholar] [CrossRef]
- Luka, Z.; Cerone, R.; Phillips, J.A., III; Mudd, H.S.; Wagner, C. Mutations in human glycine N-methyltransferase give insights into its role in methionine metabolism. Hum. Genet. 2002, 110, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Chantar, M.L.; Vázquez-Chantada, M.; Ariz, U.; Martínez, N.; Varela, M.; Luka, Z.; Capdevila, A.; Rodríguez, J.; Aransay, A.M.; Matthiesen, R.; et al. Loss of the glycine N-methyltransferase gene leads to steatosis and hepatocellular carcinoma in mice. Hepatology 2008, 47, 1191–1199. [Google Scholar] [CrossRef]
- Howard, A.; Hirst, B.H. The Glycine Transporter GLYT1 in Human Intestine: Expression and Function. Biol. Pharm. Bull. 2011, 34, 784–788. [Google Scholar] [CrossRef]
- van Bergenhenegouwen, J.; Braber, S.; Loonstra, R.; Buurman, N.; Rutten, L.; Knipping, K.; Savelkoul, P.J.; Harthoorn, L.F.; Jahnsen, F.L.; Garssen, J.; et al. Oral exposure to the free amino acid glycine inhibits the acute allergic response in a model of cow’s milk allergy in mice. Nutr. Res. 2018, 58, 95–105. [Google Scholar] [CrossRef]
- Hartog, A.; Cozijnsen, M.; de Vrij, G.; Garssen, J. Collagen Hydrolysate Inhibits Zymosan-Induced Inflammation. Exp. Biol. Med. 2013, 238, 798–802. [Google Scholar] [CrossRef]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious beneficial effect of nonessential amino acid, glycine: A review. Oxid. Med. Cell Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, A.; Reimann, F.; Habib, A.M.; O’malley, D.; Williams, L.; Simpson, A.K.; Gribble, F.M. The neurotransmitters glycine and GABA stimulate glucagon-like peptide-1 release from the GLUTag cell line. J. Physiol. 2005, 569, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Gannon, M.C.; Nuttall, J.A.; Nuttall, F.Q. The metabolic response to ingested glycine. Am. J. Clin. Nutr. 2002, 76, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Okekunle, A.P.; Li, Y.; Liu, L.; Du, S.; Wu, X.; Chen, Y.; Li, Y.; Qi, J.; Sun, C.; Feng, R. Abnormal circulating amino acid profiles in multiple metabolic disorders. Diabetes Res. Clin. Pract. 2017, 132, 45–58. [Google Scholar] [CrossRef]
- Nguyen, D.; Hsu Jean, W.; Jahoor, F.; Sekhar Rajagopal, V. Effect of increasing glutathione with cysteine and glycine supplementation on mitochondrial fuel oxidation, insulin sensitivity, and body composition in older HIV-infected patients. J. Clin. Endocrinol. Metab. 2014, 99, 169–177. [Google Scholar] [CrossRef]
- Chen, J.; Ma, X.; Yang, Y.; Dai, Z.; Wu, Z.; Wu, G. Glycine enhances expression of adiponectin and IL-10 in 3T3-L1 adipocytes without affecting adipogenesis and lipolysis. Amino Acids 2018, 50, 629–640. [Google Scholar] [CrossRef]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Forward Primer | Reverse Primer | Probe (Roche) | Accession Number |
|---|---|---|---|---|
| Housekeeping | ||||
| 36B4 | F: 5′-AAGCGCGTCCTGGCATTGTCT-3′ | R: 5′-CCGCAGGGGCAGCAGTGGT-3′ | 72 | NM_007475.4 |
| Genes of interest | ||||
| GPR21 | F: 5′-GAACTCCACCTGGGATGGTA-3′ | R: 5′-GTAGCCCAGTGCCAGAAGAC-3′ | 46 | NM_177383.4 |
| GPR26 | F: 5′-GCCAGAGCAAAGGGAGGT-3′ | R: 5′-AGGCAATGGTGGCAGTTATT-3′ | 10 | NM_173410.3 |
| GPR39 | F: 5′-CGGCGCAGTAACTCTTCC-3′ | R: 5′ -GGCCTCAGTCTGAAAAGTGC-3′ | 74 | NM_027677.2 |
| GPR82 | F: 5′-GGAACAGAAAATATGACCTGATTCAT-3′ | R: 5′- GAGGGCCTAGCACATAGCAA ′ | 36 | NM_175669.4 |
| GPR6 | F: 5′-ACATGCCAGCCTTTGGTG-3′ | R: 5′-GCACCACTGACACCTCAAGA-3′ | 40 | NM_199058.2 |
| TNF-α | F: 5′-TCTTCTCATTCCTGCTTGTGG-3′ | R: 5′-GGTCTGGGCCATAGAACTGA -3′ | 49 | NM_001278601.1 |
| IL-6 | F: 5′-ACAAAGCCAGAGTCCTTCAGA-3′ | R: 5′-TGGTCCTTAGCCACTCCTTC-3′ | 78 | NM_001314054.1 |
| AdipoQ | F: 5′-GGCTCTGTGCTCCTCCATCT-3′ | R: 5′-AGAGTCGTTGACGTTATCTGCA-3′ | 1 | NM_009605.5 |
| IL-10 | F: 5′-CCCTGGGTGAGAAGCTGAAG-3′ | R: 5′-GGGGAAGAACGCATCTGCTA-3′ | 30 | NM_010548.2 |
| Gene Expression | First Component Loading (Correlation) | Second Component Loading (Correlation) |
|---|---|---|
| GPR21 | 0.328 (0.760 **) | 0.431 (0.566 *) |
| GPR26 | 0.367 (0.850 **) | −0.190 (−0.250) |
| GPR39 | −0.058(−0.136) | −0.589 (−0.773 **) |
| GPR82 | 0.417 (0.966 **) | 0.018 (0.024) |
| GPR6 | 0.334 (0.775 **) | −0.466 (−0.612 *) |
| TNFα | −0.280 (−0.648 **) | −0.169 (−0.222) |
| AdipoQ | 0.359 (0.832 **) | 0.305 (0.400) |
| IL-6 | −0.357 (−0.829 **) | 0.010 (0.013) |
| IL-10 | 0.367 (0.851 **) | −0.302 (−0.394) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Rojas, R.A.; Aguayo-Cerón, K.A.; Vargas-De-León, C.; Cabrera-Becerra, S.E.; Almanza-Pérez, J.C.; Huang, F.; Villafaña, S.; Romero-Nava, R. Glycine Effect on the Expression Profile of Orphan Receptors GPR21, GPR26, GPR39, GPR82 and GPR6 in a Model of Inflammation in 3T3-L1 Cells. Life 2022, 12, 1687. https://doi.org/10.3390/life12111687
Gutiérrez-Rojas RA, Aguayo-Cerón KA, Vargas-De-León C, Cabrera-Becerra SE, Almanza-Pérez JC, Huang F, Villafaña S, Romero-Nava R. Glycine Effect on the Expression Profile of Orphan Receptors GPR21, GPR26, GPR39, GPR82 and GPR6 in a Model of Inflammation in 3T3-L1 Cells. Life. 2022; 12(11):1687. https://doi.org/10.3390/life12111687
Chicago/Turabian StyleGutiérrez-Rojas, Rocío Alejandra, Karla Aidee Aguayo-Cerón, Cruz Vargas-De-León, Sandra Edith Cabrera-Becerra, Julio Cesar Almanza-Pérez, Fengyang Huang, Santiago Villafaña, and Rodrigo Romero-Nava. 2022. "Glycine Effect on the Expression Profile of Orphan Receptors GPR21, GPR26, GPR39, GPR82 and GPR6 in a Model of Inflammation in 3T3-L1 Cells" Life 12, no. 11: 1687. https://doi.org/10.3390/life12111687
APA StyleGutiérrez-Rojas, R. A., Aguayo-Cerón, K. A., Vargas-De-León, C., Cabrera-Becerra, S. E., Almanza-Pérez, J. C., Huang, F., Villafaña, S., & Romero-Nava, R. (2022). Glycine Effect on the Expression Profile of Orphan Receptors GPR21, GPR26, GPR39, GPR82 and GPR6 in a Model of Inflammation in 3T3-L1 Cells. Life, 12(11), 1687. https://doi.org/10.3390/life12111687