
African Swine Fever Virus: A Review

Abstract
1. Introduction
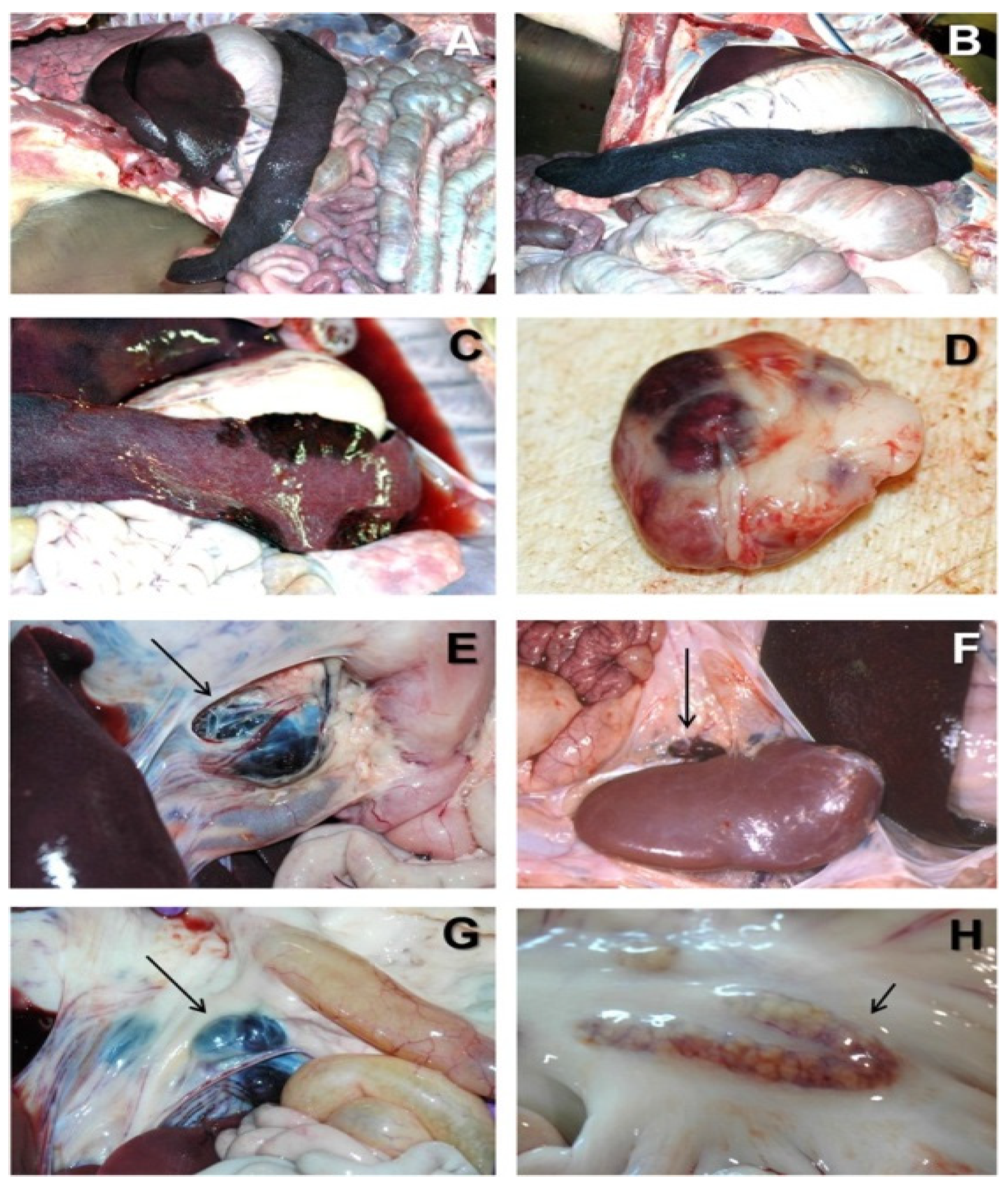
2. African Swine Fever
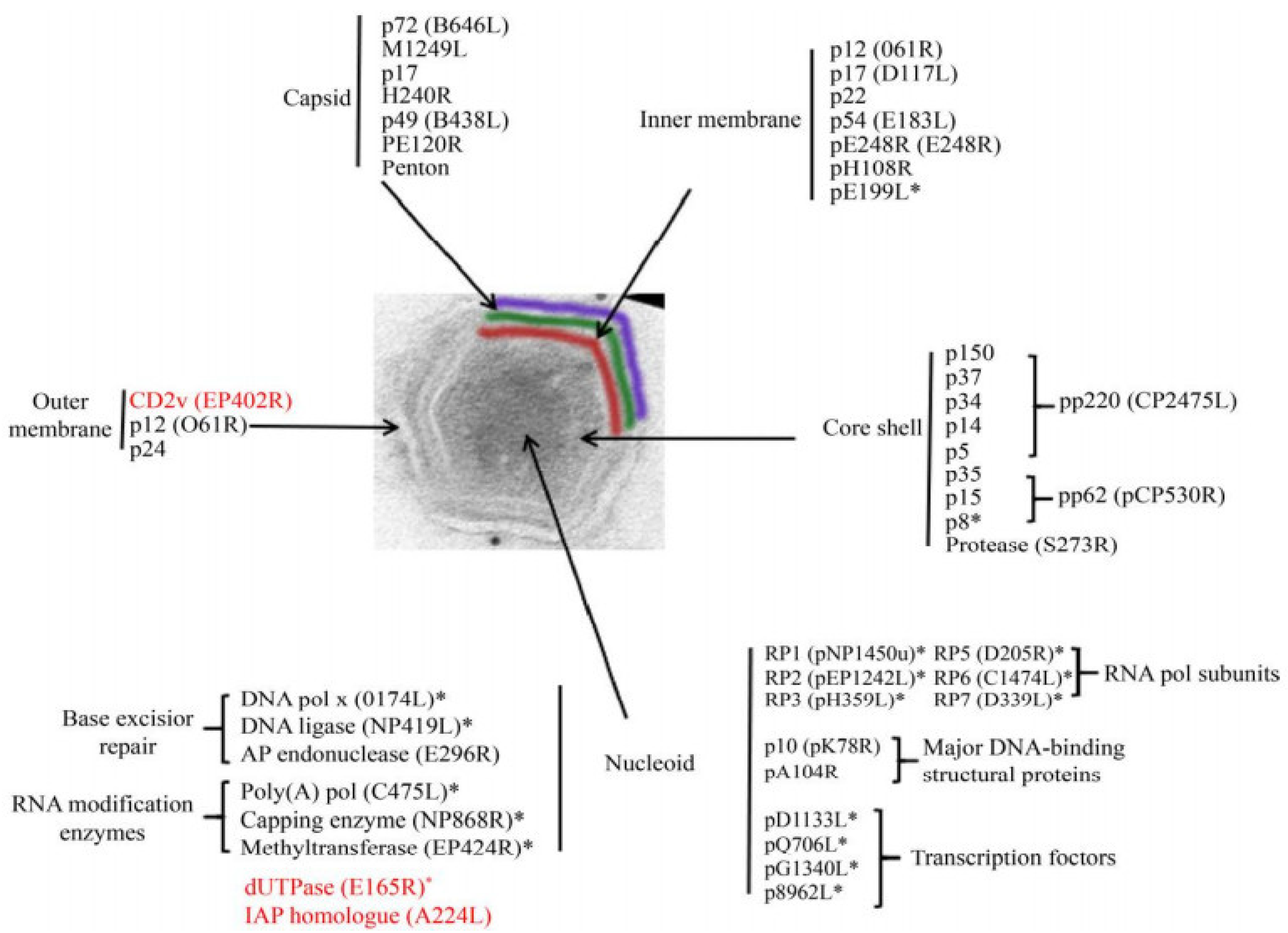
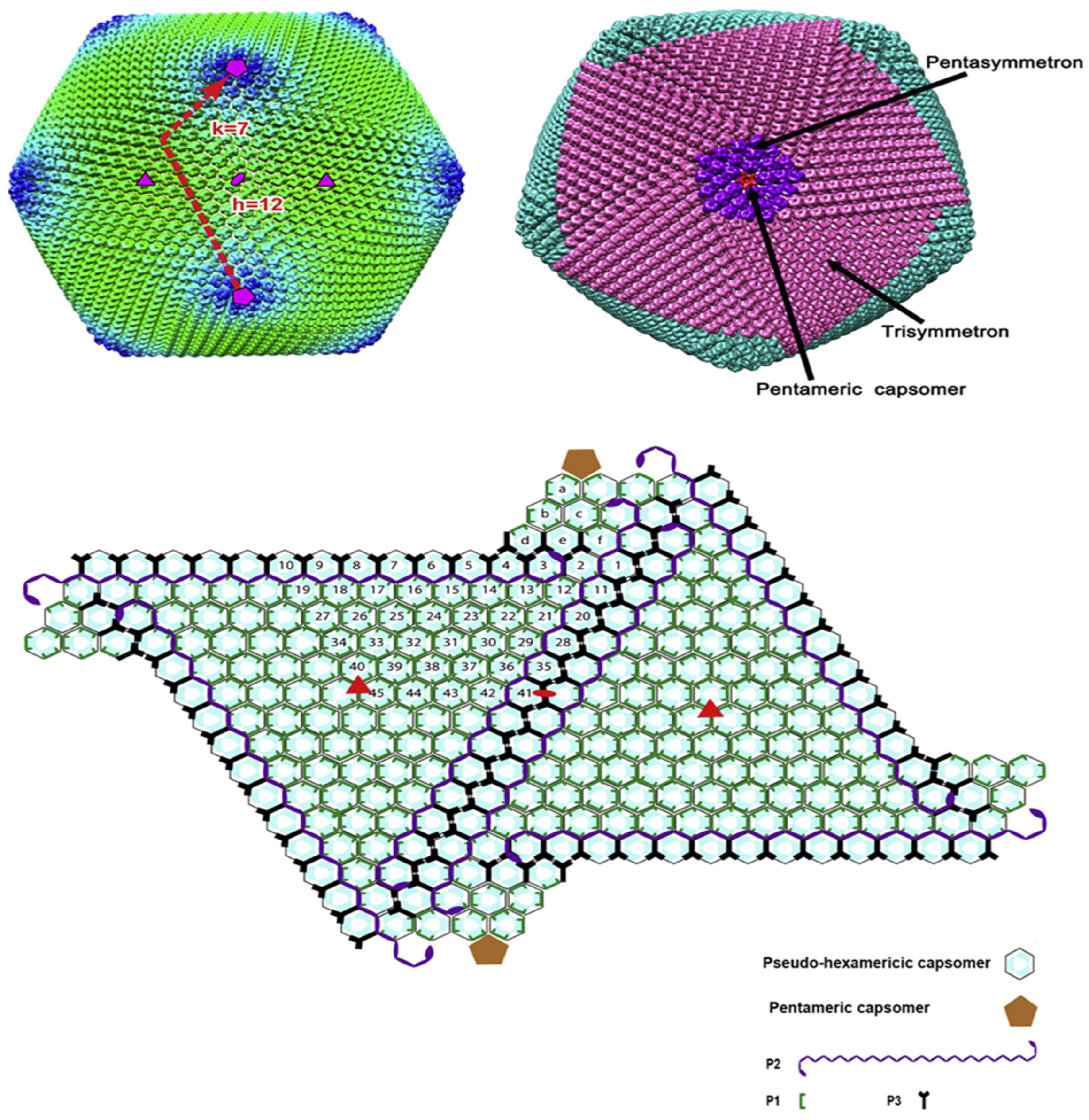
3. ASFV Structure
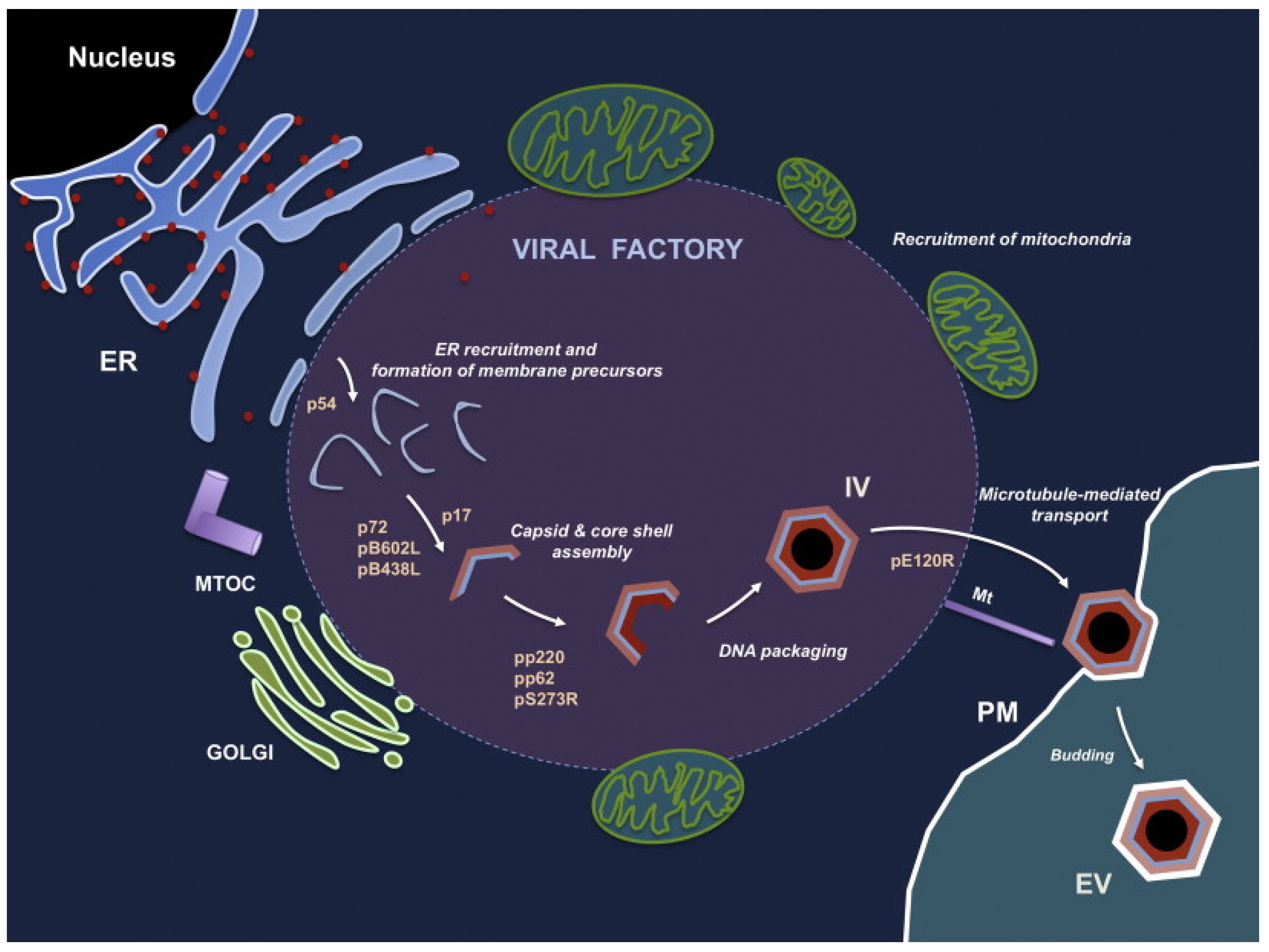
4. Virus Replication
5. Virulence Genes
6. Immunoescape
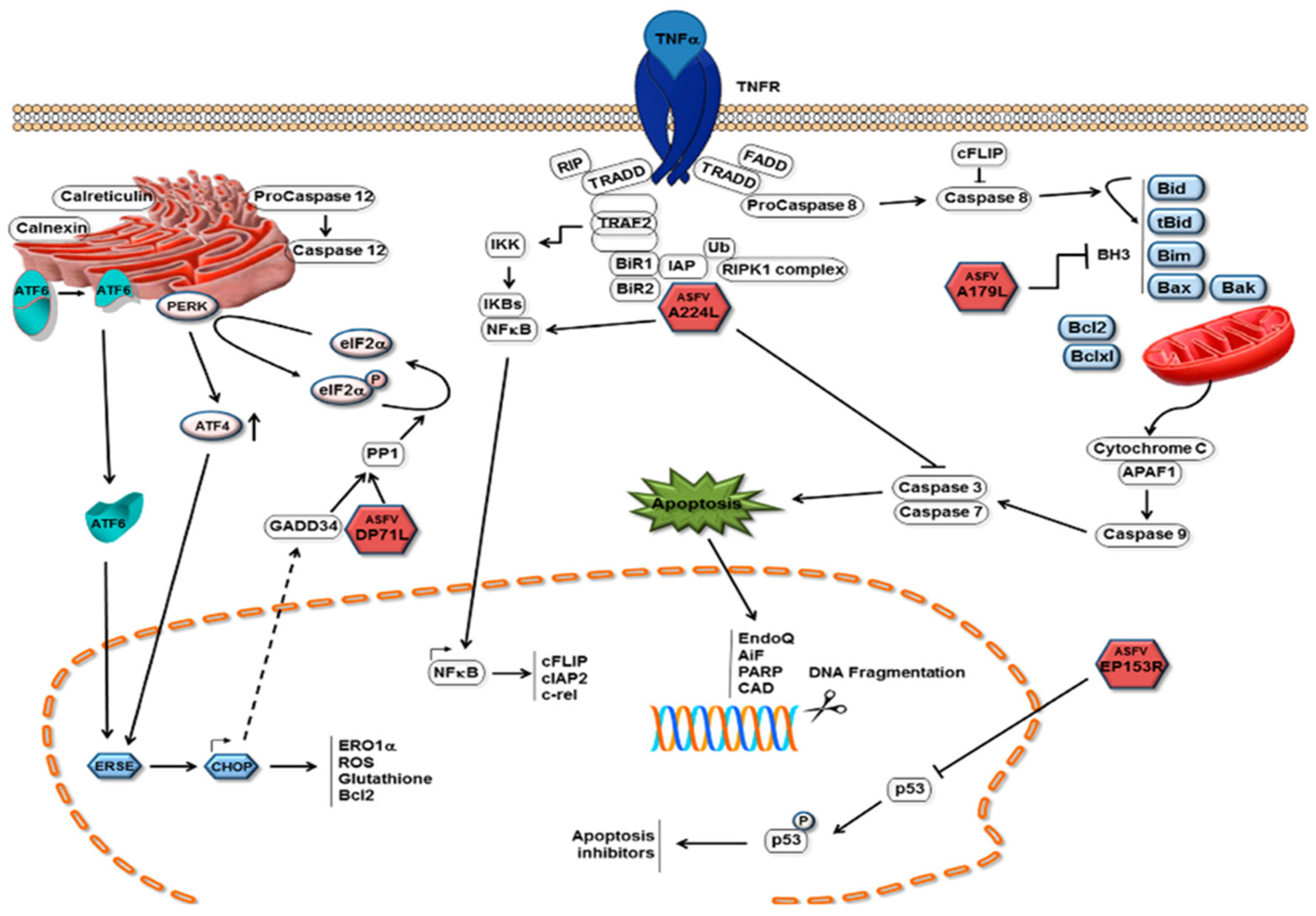
6.1. Apoptosis
6.2. Autophagy
7. Virus Inactivation
- (1)
- 1% formaldehyde;
- (2)
- Sodium hypochlorite (0.03% to 0.0075%);
- (3)
- 2% caustic soda solution (the strongest virucidal agent);
- (4)
- Glutaraldehyde, formic;
- (5)
- 1% sodium or calcium hydroxide (effective at virus inactivation in slurry at 4°C);
- (6)
- v Phenols—lysol, lysephoform, and creolin;
- (7)
- Chemical compounds based on lipid solvents [212];
- (8)
- Multi-constituent compounds such as Virkon (1:100), Lysoformin, and Desoform, Octyldodeceth-20 (OD-20) surfactants, active substances, organic acids, and glycosal;
- (9)
- Triton X-100, and NP-40 [217].
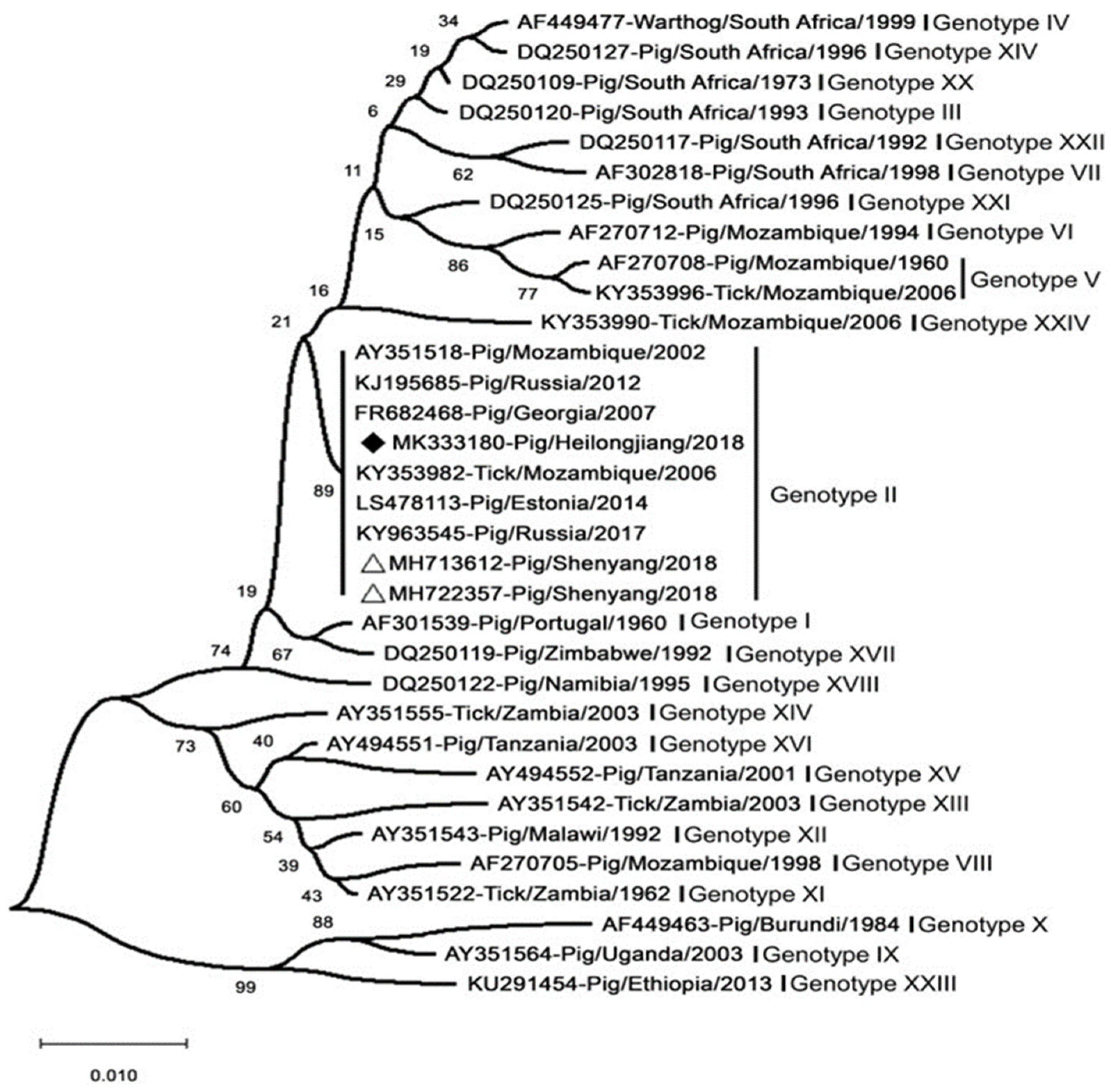
8. Virus Typing
9. Disease Dissemination and Control
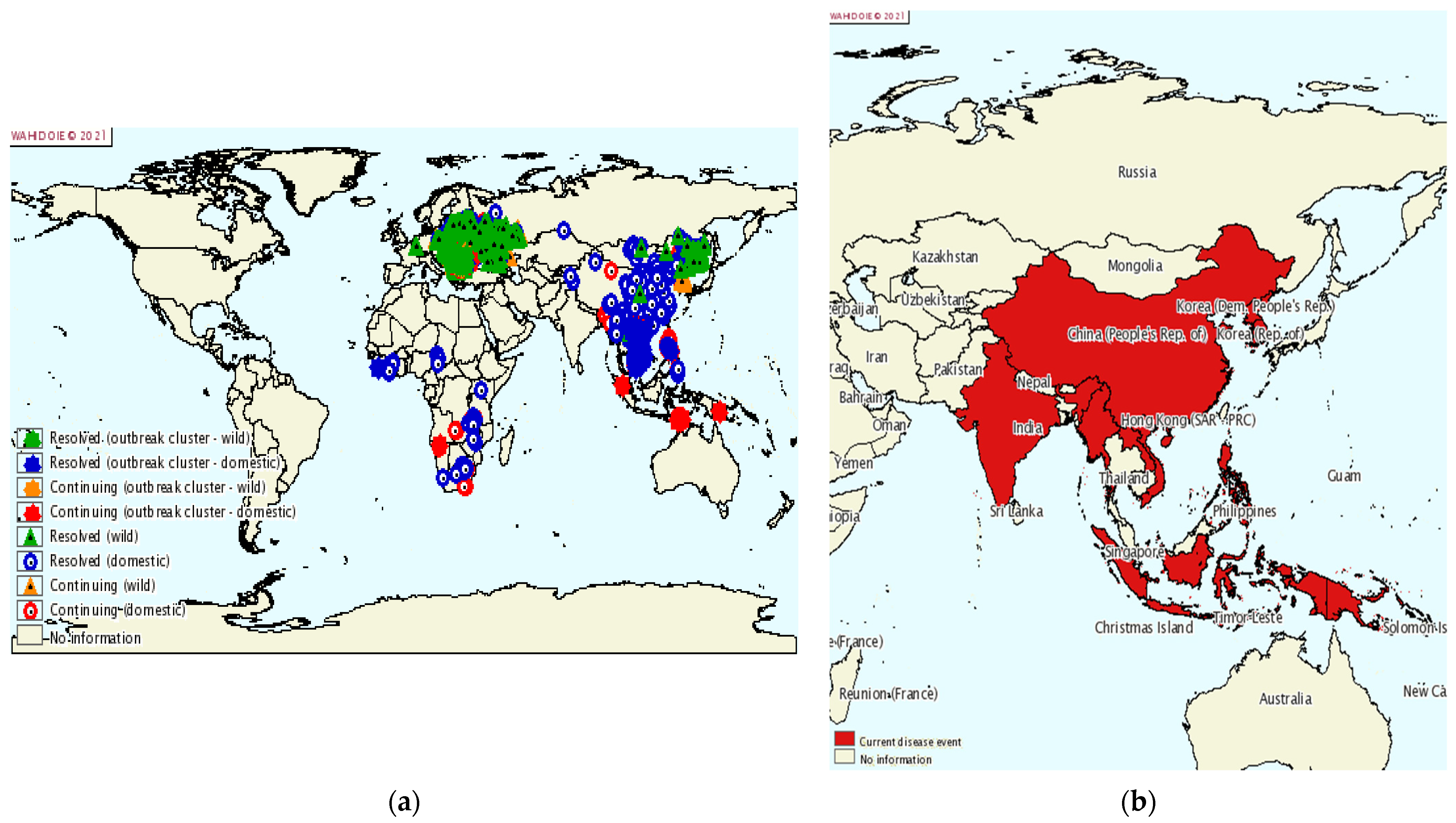
9.1. Dissemination
9.2. Control
9.2.1. Government
9.2.2. Breeding Farms
- (1)
- Keep materials that will eventually enter the piggery out of the piggery at first, until they have been disinfected in the external disinfection area.
- (2)
- The exit for the selling of pigs should be built on the outside of the enclosure.
- (3)
- No visiting vehicles should be allowed inside the enclosure.
- (4)
- Staff should be strictly bathed, and their clothing disinfected before entering the farm.
- (5)
- Rodent-proofing should be established along the fence to keep wild animals out.
- (6)
- Pens should be regularly disinfected.
- (7)
- The breeding density should not be too high.
- (8)
- Pigs should be regularly tested.
- (9)
- Emergency treatment should be initiated after an outbreak of illness.
9.2.3. Emergency Handling
10. Vaccination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Type | ASFV Target Protein (Strain) | Number of Immunizations; Dose, Adjuvant | Specific/Neutralizing Antibodies | T Cell Response | Challenge Strain; Dose | Clinical Outcome | Reference |
|---|---|---|---|---|---|---|---|
| Baculovirus-expressed proteins | CD2v (E75CV) | 3×; 0.5–1 × 107 HAU + Freund’s adjuvant | Yes; No | NA | E75; 4 × 102 | 100% protection, n = 3/3 | [247] |
| Baculovirus-expressed proteins | p30, p54, p54 + p30 (E75) | 3×; 100 μg + Freund’s adjuvant | Yes; Yes | NA | E75; 5 × 102 | 50% protection, n = 3/6 | [248] |
| Baculovirus-expressed proteins | p54/p30 chimera (E75) | 5×; 100 μg + Freund’s adjuvant | Yes; Yes | NA | E75; 5 × 102 | 100% protection, n = 2/2 | [249] |
| Baculovirus-expressed proteins | p54 + p30 + p72 + p22 (Pr4) | 4×; 200 μg + Freund’s adjuvant | Yes; Yes | NA | Pr4; 104 | Slight delay of clinical disease and viremia; no protection, (n = 0/6) | [250] |
| HEK cell-expressed proteins | p72, p54, p12 (Georgia 2007/1) | 2×; 200 μg/antigen + TS6 adjuvant | Yes; NA | Some | NA | NA | [251] |
| DNA (pCMV) | SLA-II/p54/p30 fusion (E75) | 3×; 600 μg | Yes; No | Yes | E75; 104 | No protection, (n = 0/4) | [252] |
| DNA (pCMV) | sHA/p54/p30 fusion (E75) | 3× and 4×; 600 μg | Yes; No | Yes | E75; 104 | No protection, (n = 0/6) | [253] |
| DNA (pCMV) | Ub/sHA/p54/p30 fusion (E75) | 2× and 4×; 600 μg | Not detectable | Yes | E75; 104 | Partial protection, (2 immunizations, n = 2/6; 4 immunizations, n = 1/6) | [253] |
| DNA expression library | 80 ORFs fragments fused with Ub (Ba71V) | 2×; 600 μg | Yes—after challenge; NA | Yes-after challenge | E75; 104 | 60% protection (n = 6/10) | [254] |
| BacMam | sHA/p54/p30 fusion (E75) | 3×; 107 PFU | No (only after challenge); No | Yes | E75; 2× sublethal challenge 102 | Partial protection (n = 4/6) | [255] |
| Adenovirus | p30 + p54 + pp62 + p72 (Georgia 2007/1) | 2×; 1010 or 1011 per Ad5-antigen + adjuvants | Yes; NA | Yes | NA | NA | [256] |
| Adenovirus | A151R + B119L + B602L + EP402RΔPRR + B438L + K205R + A104R (Georgia 2007/1) | 2×; 1011 per Ad5-antigen + adjuvant | Yes; NA | Yes | NA | NA | [156] |
| Vaccinia virus Ankara | p72, C-type Lectin, CD2v (Georgia 2007/1) | 2×; rVACV-ASFV 107 TCID50 | No; NA | Yes | NA | NA | [251] |
| Alphavirus RPs | p30, p54, p72, sHA/72 (Ba71V) | 3×; 2–4.5 × 107 RPs | Yes; NA | NA | NA | NA | [257] |
| DNA–Protein | Combinations of DNA and protein: p15, p30, p35, p54, p72, CD2v, CP312R, g5R (Georgia 2007/1; Ba71V) | 3×; 100 μg per DNA, 100 μg protein + ISA25 adjuvant | Yes; Yes | Some | NA | NA | [258] |
| DNA–Protein | Proteins: p15, p35, p54, p17; DNA: CD2v, p72, p54, p30, p17 (Georgia 2007/1; Ba71V) | 3×; 100 μg per DNA, 100 μg protein + ISA25 adjuvant | Yes; No | Some | Armenia 2007; 360 HAU | No protection; disease enhancement | [259] |
| DNA prime + vaccinia virus boost | 47 antigens (Georgia 2007/1) | Prime 2×: 10 μg pCMV-DNA + CpG oligo adjuvant;Boost 2×: 108 PFU rVACV-ASFV | Yes; No | Yes | Georgia 2007/1; 104 | No protection; reduced viral load, higher clinical scores | [260] |
| Vaccinia virus prime + protein boost | p72, C-type Lectin, CD2v (Georgia 2007/1) | Prime: rVACV-ASFV 107 TCID50; Boost: 200 μg/antigen + TS6 adjuvant | NA | Yes | NA | NA | [251] |
| Alphavirus RP prime + live attenuated ASFV boost | p30 (Ba71V) + OURT88/3 | Prime 2×: 2–4.5 × 107 RPs; Boost: 104 TCID50 OURT88/3 | Yes; Yes | NA | NA | NA | [257] |
| p30 referred to as p32; CD2v also referred to as HA = hemagglutinin; NA = not available. | |||||||
| Strain | Virulence | Challenge | Protection | References |
|---|---|---|---|---|
| NH/P68 | Low | Heterologous strain L60 | 100% | [261] |
| OUR T88/3 | Low | Heterologous strain Benin 97/1 | 85.70% | [262] |
| OUR T88/3 | Low | Homologous strain OURT88/1 | 50–100% | [263] |
| Lv17/WB/Rie1 | Low | Homologous strain HAD Latvian ASFV | 100% | [264] |
| Strain | Virulence | Deleted Genes | Challenge | Protection | References |
|---|---|---|---|---|---|
| OUR T88/3 | Low | DP71L and DP96R | Homologous strain OURT88/1 | 100% | [261] |
| Georgia 2007/1 | High | DP96R(UK) and B119L(9GL) | Homologous strain Georgia 2007/1 | 100% | [18] |
| Benin 97/1 | High | DP148R | Homologous strain Benin 97/1 | 100% | [164] |
| Georgia 2007/1 | High | MGF505/360(6) | Homologous strain Georgia 2007/1 | 100% | [97] |
| Georgia 2007/1 | High | MGF505/360 and B119L(9GL) | Homologous strain Georgia 2007/1 | 100% | [265] |
| Benin 97/1 | High | MGF505/530/360 | Homologous strain Benin 97/1 | 100% | [266] |
| BA71 | High | EP402R(CD2v) | Heterologous strain Georgia 2007/1 | 100% | [13] |
| Georgia 2010 | High | EP402R(CD2v) | Homologous strain Georgia 2010 | 100% | [16] |
| HLJ/2018 | High | MGF505/360(6) and EP402R(CD2v) | Homologous strain HLJ/2018 | 100% | [242] |
| Georgia 2010 | High | I177L | Homologous strain Georgia 2010 | 100% | [14] |
| Georgia 2007/1 | High | L83L | Do not verify | Do not verify | [267] |
| Georgia 2007/1 | High | B119L, DP71L and DP96R | Homologous strain Georgia 2007/1 | 0 | [268] |
| NH/P68 | Low | A276R | Heterologous strain virulent Arm07 | 0 | [269] |
| Genes | Strains | Genotype | Minimal Protective Dose | Route | Challenge | Gene Function | References |
|---|---|---|---|---|---|---|---|
| A137R | Georgia2007/1 | II | 102HAD50 | IM | Georgia2007/1 | Unknown | [152] |
| I226R | SY18 | II | 102HAD50 | IM | Georgia2007/1 | Unknown | [270] |
| L7L-L11L | SY18 | II | 103HAD50 | IM | SY18 | Unknown | [271] |
| MGF505/360 and EP402R | HLJ/18 | II | 103HAD50 105HAD50 | IM ON | HLJ/18 | Hemadsorbing and inhibition of type I | [242] |
| EP402R | Ba71V | I | 104HAD50 | IM | Ba71V E75 Georgia2007/1 | Interferon responses Hemadsorbing | [13] |
| I177L | Georgia2007/1 | II | 102HAD50 106HAD50 | IM ON | Georgia2007/1 Georgia2007/1 | Unknown | [15,16] |
11. Diagnosis
12. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Penrith, M.L.; Vosloo, W.; Jori, F.; Bastos, A.D. African swine fever virus eradication in Africa. Virus Res. 2013, 173, 228–246. [Google Scholar] [CrossRef]
- Boklund, A.; Cay, B.; Depner, K.; Földi, Z.; Guberti, V.; Masiulis, M.; Miteva, A.; More, S.; Olsevskis, E.; Šatrán, P.; et al. Epidemiological Analyses of African Swine Fever in the European Union (November 2017 until November 2018). EFSA J. 2018, 16, e05494. [Google Scholar]
- Dixon, L.K.; Islam, M.; Nash, R.; Reis, A.L. African Swine Fever Virus Evasion of Host Defences. Virus Res. 2019, 266, 25–33. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef]
- Pikalo, J.; Zani, L.; Hühr, J.; Beer, M.; Blome, S. Pathogenesis of African Swine Fever in Domestic Pigs and European Wild Boar—Lessons Learned from Recent Animal Trials. Virus Res. 2019, 271, 197614. [Google Scholar] [CrossRef]
- Sanchez-Vizcaino, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An Update on the Epidemiology and Pathology of African Swine Fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef]
- Salguero, F.J. Comparative Pathology and Pathogenesis of African Swine Fever Infection in Swine. Front. Vet. Sci. 2020, 7, 282. [Google Scholar] [CrossRef]
- Gomez-Villamandos, J.C.; Carrasco, L.; Bautista, M.J.; Sierra, M.A.; Quezada, M.; Hervas, J.; de Lara F Chacón, M.; Ruiz-Villamor, E.; Salguero, F.J.; Sonchez-Cordon, P.J.; et al. African Swine Fever and Classical Swine Fever: A Review of the Pathogenesis. Dtsch. Tierarztl. Wochenschr. 2003, 110, 165–169. [Google Scholar]
- Oura, C.A.; Powell, P.P.; Anderson, E.; Parkhouse, R.M. The Pathogenesis of African Swine Fever in the Resistant Bushpig. J. Gen. Virol. 1998, 79 Pt 6, 1439–1443. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Nurmoja, I.; Cano-Gomez, C.; Cvetkova, S.; Frant, M.; Wozniakowski, G.; Simon, A.; Perez, C.; Nieto, R.; et al. Dynamics of African Swine Fever Virus (ASFV) Infection in Domestic Pigs Infected with Virulent, Moderate Virulent and Attenuated Genotype Ii ASFV European Isolates. Transbound. Emerg. Dis. 2021, 68, 2826–2841. [Google Scholar] [CrossRef]
- Nguyen-Thi, T.; Pham-Thi-Ngoc, L.; Nguyen-Ngoc, Q.; Dang-Xuan, S.; Lee, H.S.; Nguyen-Viet, H.; Padungtod, P.; Nguyen-Thu, T.; Nguyen-Thi, T.; Tran-Cong, T.; et al. An Assessment of the Economic Impacts of the 2019 African Swine Fever Outbreaks in Vietnam. Front. Vet. Sci. 2021, 8, 686038. [Google Scholar] [CrossRef]
- Monteagudo, P.L.; Lacasta, A.; Lopez, E.; Bosch, L.; Collado, J.; Pina-Pedrero, S.; Correa-Fiz, F.; Accensi, F.; Navas, M.J.; Vidal, E.; et al. BA71ΔCD2: A New Recombinant Live Attenuated African Swine Fever Virus with Cross-Protective Capabilities. J. Virol. 2017, 91, e01058-17. [Google Scholar] [CrossRef]
- Lewis, T.; Zsak, L.; Burrage, T.G.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Rock, D.L. An African Swine Fever Virus Erv1-Alr Homologue, 9gl, Affects Virion Maturation and Viral Growth in Macrophages and Viral Virulence in Swine. J. Virol. 2000, 74, 1275–1285. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Gay, C.G.; Gladue, D.P. ASFV-G-∆I177l as an Effective Oral Nasal Vaccine against the Eurasia Strain of Africa Swine Fever. Viruses 2021, 13, 765. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177l Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef]
- Moore, D.M.; Zsak, L.; Neilan, J.G.; Lu, Z.; Rock, D.L. The African Swine Fever Virus Thymidine Kinase Gene Is Required for Efficient Replication in Swine Macrophages and for Virulence in Swine. J. Virol. 1998, 72, 10310–10315. [Google Scholar] [CrossRef]
- O’Donnell, V.; Risatti, G.R.; Holinka, L.G.; Krug, P.W.; Carlson, J.; Velazquez-Salinas, L.; Azzinaro, P.A.; Gladue, D.P.; Borca, M.V. Simultaneous Deletion of the 9gl and UK Genes from the African Swine Fever Virus Georgia 2007 Isolate Offers Increased Safety and Protection against Homologous Challenge. J. Virol. 2017, 91, e01760-16. [Google Scholar] [CrossRef]
- Tran, X.H.; Le, T.T.P.; Nguyen, Q.H.; Do, T.T.; Nguyen, V.D.; Gay, C.G.; Borca, M.V.; Gladue, D.P. African Swine Fever Virus Vaccine Candidate ASFV-G-Deltai177l Efficiently Protects European and Native Pig Breeds against Circulating Vietnamese Field Strain. Transbound. Emerg. Dis. 2022, 69, e497–e504. [Google Scholar] [CrossRef]
- Tran, X.H.; Phuong, L.T.T.; Huy, N.Q.; Thuy, D.T.; Nguyen, V.D.; Quang, P.H.; Ngon, Q.V.; Rai, A.; Gay, C.G.; Gladue, D.P.; et al. Evaluation of the Safety Profile of the ASFV Vaccine Candidate ASFV-G-Deltai177l. Viruses 2022, 14, 896. [Google Scholar] [CrossRef]
- More, S.; Miranda, M.A.; Bicout, D.; Bøtner, A.; Butterworth, A.; Calistri, P.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Michel, V.; et al. African Swine Fever in Wild Boar. EFSA J. 2018, 16, e05344. [Google Scholar]
- Pietschmann, J.; Mur, L.; Blome, S.; Beer, M.; Pérez-Sánchez, R.; Oleaga, A.; Sánchez-Vizcaíno, J.M. African Swine Fever Virus Transmission Cycles in Central Europe: Evaluation of Wild Boar-Soft Tick Contacts through Detection of Antibodies against Ornithodoros Erraticus Saliva Antigen. BMC Vet. Res. 2016, 12, 1. [Google Scholar] [CrossRef]
- Pereira de Oliveira, R.; Hutet, E.; Paboeuf, F.; Duhayon, M.; Boinas, F.; de Leon, A.P.; Filatov, S.; Vial, L.; le Potier, M.F. Comparative Vector Competence of the Afrotropical Soft Tick Ornithodoros Moubata and Palearctic Species, O. Erraticus and O. Verrucosus, for African Swine Fever Virus Strains Circulating in Eurasia. PLoS ONE 2019, 14, e0225657. [Google Scholar] [CrossRef]
- Greig, A. Pathogenesis of African Swine Fever in Pigs Naturally Exposed to the Disease. J. Comp. Pathol. 1972, 82, 73–79. [Google Scholar] [CrossRef]
- Carrasco, L.; Núñez, A.; Salguero, F.J.; Segundo, F.D.S.; Sánchez-Cordón, P.; Gómez-Villamandos, J.C.; Sierra, M.A. African Swine Fever: Expression of Interleukin-1 Alpha and Tumour Necrosis Factor-Alpha by Pulmonary Intravascular Macrophages. J. Comp. Pathol. 2002, 126, 194–201. [Google Scholar] [CrossRef]
- Wu, K.; Zhang, Y.; Zeng, S.; Liu, X.; Li, Y.; Li, X.; Chen, W.; Li, Z.; Qin, Y.; Chen, J.; et al. Development and Application of Raa Nucleic Acid Test Strip Assay and Double Raa Gel Electrophoresis Detection Methods for ASFV and Csfv. Front. Mol. Biosci. 2021, 8, 811824. [Google Scholar] [CrossRef]
- Konno, S.; Taylor, W.D.; Hess, W.R.; Heuschele, W.P. Spleen Pathology in African Swine Fever. Cornell Vet. 1972, 62, 486–506. [Google Scholar]
- Gómez-Villamandos, J.C.; Hervás, J.; Méndez, A.; Carrasco, L.; de las Mulas, J.M.; Villeda, C.J.; Wilkinson, P.J.; Sierra, M.A. Experimental African Swine Fever: Apoptosis of Lymphocytes and Virus Replication in Other Cells. J. Gen. Virol. 1995, 76 Pt 9, 2399–2405. [Google Scholar] [CrossRef]
- Wardley, R.C.; Wilkinson, P.J. The Association of African Swine Fever Virus with Blood Components of Infected Pigs. Arch. Virol. 1977, 55, 327–334. [Google Scholar] [CrossRef]
- Haresnape, J.M.; Wilkinson, P.J.; Mellor, P.S. Isolation of African Swine Fever Virus from Ticks of the Ornithodoros Moubata Complex (Ixodoidea: Argasidae) Collected within the African Swine Fever Enzootic Area of Malawi. Epidemiol. Infect. 1988, 101, 173–185. [Google Scholar] [CrossRef]
- Karalyan, Z.; Zakaryan, H.; Arzumanyan, H.; Sargsyan, K.; Voskanyan, H.; Hakobyan, L.; Abroyan, L.; Avetisyan, A.; Karalova, E. Pathology of Porcine Peripheral White Blood Cells during Infection with African Swine Fever Virus. BMC Vet. Res. 2012, 8, 18. [Google Scholar] [CrossRef]
- Ramiro-Ibáñez, F.; Ortega, A.; Ruiz-Gonzalvo, F.; Escribano, J.M.; Alonso, C. Modulation of Immune Cell Populations and Activation Markers in the Pathogenesis of African Swine Fever Virus Infection. Virus Res. 1997, 47, 31–40. [Google Scholar] [CrossRef]
- Salguero, F.J.; Ruiz-Villamor, E.; Bautista, M.J.; Sanchez-Cordon, P.J.; Carrasco, L.; Gomez-Villamandos, J.C. Changes in Macrophages in Spleen and Lymph Nodes during Acute African Swine Fever: Expression of Cytokines. Vet. Immunol. Immunopathol. 2002, 90, 11–22. [Google Scholar] [CrossRef]
- Carrasco, L.; de Lara, F.C.; de las Mulas, J.M.; Gomez-Villamandos, J.C.; Perez, J.; Wilkinson, P.J.; Sierra, M.A. Apoptosis in Lymph Nodes in Acute African Swine Fever. J. Comp. Pathol. 1996, 115, 415–428. [Google Scholar] [CrossRef]
- Oura, C.A.; Powell, P.P.; Parkhouse, R.M. Detection of African Swine Fever Virus in Infected Pig Tissues by Immunocytochemistry and in Sity Hybridisation. J. Virol. Methods 1998, 72, 205–217. [Google Scholar] [CrossRef]
- Pikalo, J.; Schoder, M.E.; Sehl, J.; Breithaupt, A.; Tignon, M.; Cay, A.B.; Gager, A.M.; Fischer, M.; Beer, M.; Blome, S. The African Swine Fever Virus Isolate Belgium 2018/1 Shows High Virulence in European Wild Boar. Transbound. Emerg. Dis. 2020, 67, 1654–1659. [Google Scholar] [CrossRef]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: ASFarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Dixon, L.K.; Sun, H.; Roberts, H. African Swine Fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef]
- Iyer, L.M.; Balaji, S.; Koonin, E.V.; Aravind, L. Evolutionary Genomics of Nucleo-Cytoplasmic Large DNA Viruses. Virus Res. 2006, 117, 156–184. [Google Scholar] [CrossRef]
- Colson, P.; de Lamballerie, X.; Fournous, G.; Raoult, D. Reclassification of Giant Viruses Composing a Fourth Domain of Life in the New Order Megavirales. Intervirology 2012, 55, 321–332. [Google Scholar] [CrossRef]
- Carrascosa, J.L.; Carazo, J.M.; Carrascosa, A.L.; García, N.; Santisteban, A.; Viñuela, E. General Morphology and Capsid Fine Structure of African Swine Fever Virus Particles. Virology 1984, 132, 160–172. [Google Scholar] [CrossRef]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.L.; Wang, M.; Gao, Y.; Li, F.; Wang, J.F.; Bu, Z.; Rao, Z.G.; et al. Architecture of African Swine Fever Virus and Implications for Viral Assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef]
- Alejo, A.; Andrés, G.; Viñuela, E.; Salas, M.L. The African Swine Fever Virus Prenyltransferase Is an Integral Membrane Trans-Geranylgeranyl-Diphosphate Synthase. J. Biol. Chem. 1999, 274, 18033–18039. [Google Scholar] [CrossRef]
- Dixon, L.K.; Chapman, D.A.; Netherton, C.L.; Upton, C. African Swine Fever Virus Replication and Genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Andres, G.; Garcia-Escudero, R.; Salas, M.L.; Rodriguez, J.M. Repression of African Swine Fever Virus Polyprotein Pp220-Encoding Gene Leads to the Assembly of Icosahedral Core-Less Particles. J. Virol. 2002, 76, 2654–2666. [Google Scholar] [CrossRef]
- Yáñez, R.J.; Rodríguez, J.M.; Nogal, M.L.; Yuste, L.; Enríquez, C.; Rodriguez, J.F.; Viñuela, E. Analysis of the Complete Nucleotide Sequence of African Swine Fever Virus. Virology 1995, 208, 249–278. [Google Scholar] [CrossRef]
- Salas, M.L.; Andrés, G. African Swine Fever Virus Morphogenesis. Virus Res. 2013, 173, 29–41. [Google Scholar] [CrossRef]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G. A Proteomic Atlas of the African Swine Fever Virus Particle. J. Virol. 2018, 92, e01293-18. [Google Scholar]
- Andres, G.; Alejo, A.; Salas, J.; Salas, M.L. African Swine Fever Virus Polyproteins Pp220 and Pp62 Assemble into the Core Shell. J. Virol. 2002, 76, 12473–12482. [Google Scholar] [CrossRef]
- Andrés, G.; Alejo, A.; Simón-Mateo, C.; Salas, M.L. African Swine Fever Virus Protease, a New Viral Member of the Sumo-1-Specific Protease Family. J. Biol. Chem. 2001, 276, 780–787. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.; Yang, M.; Zhang, G.; Wang, Z.; Guo, K.; Gao, Y.; Jiao, P.; Sun, J.; Chen, C.; et al. Crystal Structure of African Swine Fever Virus Ps273r Protease and Implications for Inhibitor Design. J. Virol. 2020, 94, e02125-19. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; García-Escudero, R.; Salas, M.L.; Andrés, G. African Swine Fever Virus Structural Protein P54 Is Essential for the Recruitment of Envelope Precursors to Assembly Sites. J. Virol. 2004, 78, 4299–4313. [Google Scholar] [CrossRef]
- Suárez, C.; Gutiérrez-Berzal, J.; Andrés, G.; Salas, M.L.; Rodríguez, J.M. African Swine Fever Virus Protein P17 Is Essential for the Progression of Viral Membrane Precursors toward Icosahedral Intermediates. J. Virol. 2010, 84, 7484–7499. [Google Scholar] [CrossRef]
- Liu, S.; Luo, Y.; Wang, Y.; Li, S.; Zhao, Z.; Bi, Y.; Sun, J.; Peng, R.; Song, H.; Zhu, D.; et al. Cryo-Em Structure of the African Swine Fever Virus. Cell Host Microbe 2019, 26, 836–843.e3. [Google Scholar] [CrossRef]
- García-Escudero, R.; Andrés, G.; Almazán, F.; Viñuela, E. Inducible Gene Expression from African Swine Fever Virus Recombinants: Analysis of the Major Capsid Protein P72. J. Virol. 1998, 72, 3185–3195. [Google Scholar] [CrossRef]
- Penrith, M.L.; Bastos, A.D.; Etter, E.M.C.; Beltrán-Alcrudo, D. Epidemiology of African Swine Fever in Africa Today: Sylvatic Cycle Versus Socio-Economic Imperatives. Transbound. Emerg. Dis. 2019, 66, 672–686. [Google Scholar] [CrossRef]
- Epifano, C.; Krijnse-Locker, J.; Salas, M.L.; Salas, J.; Rodríguez, J.M. Generation of Filamentous Instead of Icosahedral Particles by Repression of African Swine Fever Virus Structural Protein Pb438l. J. Virol. 2006, 80, 11456–11466. [Google Scholar] [CrossRef]
- Andrés, G.; García-Escudero, R.; Viñuela, E.; Salas, M.L.; Rodríguez, J.M. African Swine Fever Virus Structural Protein Pe120r Is Essential for Virus Transport from Assembly Sites to Plasma Membrane but Not for Infectivity. J. Virol. 2001, 75, 6758–6768. [Google Scholar] [CrossRef]
- Wen, X.; He, X.; Zhang, X.; Zhang, X.; Liu, L.; Guan, Y.; Zhang, Y.; Bu, Z. Genome Sequences Derived from Pig and Dried Blood Pig Feed Samples Provide Important Insights into the Transmission of African Swine Fever Virus in China in 2018. Emerg. Microbes Infect. 2019, 8, 303–306. [Google Scholar] [CrossRef]
- Galindo, I.; Vinuela, E.; Carrascosa, A.L. Protein Cell Receptors Mediate the Saturable Interaction of African Swine Fever Virus Attachment Protein P12 with the Surface of Permissive Cells. Virus Res. 1997, 49, 193–204. [Google Scholar] [CrossRef]
- Burmakina, G.; Malogolovkin, A.; Tulman, E.R.; Xu, W.; Delhon, G.; Kolbasov, D.; Rock, D.L. Identification of T-Cell Epitopes in African Swine Fever Virus Cd2v and C-Type Lectin Proteins. J. Gen. Virol. 2019, 100, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.; Garcia-Barreno, B.; Nogal, M.L.; Vinuela, E.; Enjuanes, L. Monoclonal Antibodies Specific for African Swine Fever Virus Proteins. J. Virol. 1985, 54, 199–206. [Google Scholar] [CrossRef]
- Alonso, C.; Galindo, I.; Cuesta-Geijo, M.A.; Cabezas, M.; Hernaez, B.; Munoz-Moreno, R. African Swine Fever Virus-Cell Interactions: From Virus Entry to Cell Survival. Virus Res. 2013, 173, 42–57. [Google Scholar] [CrossRef]
- Meier, O.; Boucke, K.; Hammer, S.V.; Keller, S.; Stidwill, R.P.; Hemmi, S.; Greber, U.F. Adenovirus Triggers Macropinocytosis and Endosomal Leakage Together with Its Clathrin-Mediated Uptake. J. Cell Biol. 2002, 158, 1119–1131. [Google Scholar] [CrossRef]
- Galindo, I.; Cuesta-Geijo, M.A.; Hlavova, K.; Munoz-Moreno, R.; Barrado-Gil, L.; Dominguez, J.; Alonso, C. African Swine Fever Virus Infects Macrophages, the Natural Host Cells, Via Clathrin- and Cholesterol-Dependent Endocytosis. Virus Res. 2015, 200, 45–55. [Google Scholar] [CrossRef]
- Sanchez, E.G.; Perez-Nunez, D.; Revilla, Y. Mechanisms of Entry and Endosomal Pathway of African Swine Fever Virus. Vaccines 2017, 5, 42. [Google Scholar] [CrossRef]
- Lithgow, P.; Takamatsu, H.; Werling, D.; Dixon, L.; Chapman, D. Correlation of Cell Surface Marker Expression with African Swine Fever Virus Infection. Vet. Microbiol. 2014, 168, 413–419. [Google Scholar] [CrossRef]
- Almazán, F.; Rodríguez, J.M.; Andrés, G.; Pérez, R.; Viñuela, E.; Rodriguez, J.F. Transcriptional Analysis of Multigene Family 110 of African Swine Fever Virus. J. Virol. 1992, 66, 6655–6667. [Google Scholar] [CrossRef]
- Hernaez, B.; Guerra, M.; Salas, M.L.; Andres, G. African Swine Fever Virus Undergoes Outer Envelope Disruption, Capsid Disassembly and Inner Envelope Fusion before Core Release from Multivesicular Endosomes. PLoS Pathog. 2016, 12, e1005595. [Google Scholar] [CrossRef]
- Sanchez, E.G.; Quintas, A.; Perez-Nunez, D.; Nogal, M.; Barroso, S.; Carrascosa, A.L.; Revilla, Y. African Swine Fever Virus Uses Macropinocytosis to Enter Host Cells. PLoS Pathog. 2012, 8, e1002754. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, B.; Alonso, C. Dynamin- and Clathrin-Dependent Endocytosis in African Swine Fever Virus Entry. J. Virol. 2010, 84, 2100–2109. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, C.; Antonio, A.; de Lima, M.C.P.; Valdeira, M.L. Cholesterol Affects African Swine Fever Virus Infection. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1998, 1393, 19–25. [Google Scholar] [CrossRef]
- Sanchez-Torres, C.; Gomez-Puertas, P.; Gomez-del-Moral, M.; Alonso, F.; Escribano, J.M.; Ezquerra, A.; Dominguez, J. Expression of Porcine Cd163 on Monocytes/Macrophages Correlates with Permissiveness to African Swine Fever Infection. Arch. Virol. 2003, 148, 2307–2323. [Google Scholar] [CrossRef]
- Salas, M.L.; Kuznar, J.; Vinuela, E. Polyadenylation, Methylation, and Capping of the Rna Synthesized in Vitro by African Swine Fever Virus. Virology 1981, 113, 484–491. [Google Scholar] [CrossRef]
- Rojo, G.; Garcia-Beato, R.; Vinuela, E.; Salas, M.L.; Salas, J. Replication of African Swine Fever Virus DNA in Infected Cells. Virology 1999, 257, 524–536. [Google Scholar] [CrossRef]
- Heath, C.M.; Windsor, M.; Wileman, T. Aggresomes Resemble Sites Specialized for Virus Assembly. J. Cell Biol. 2001, 153, 449–456. [Google Scholar] [CrossRef]
- Andrés, G.; García-Escudero, R.; Simón-Mateo, C.; Viñuela, E. African Swine Fever Virus Is Enveloped by a Two-Membraned Collapsed Cisterna Derived from the Endoplasmic Reticulum. J. Virol. 1998, 72, 8988–9001. [Google Scholar] [CrossRef]
- Almendral, J.M.; Almazan, F.; Blasco, R.; Vinuela, E. Multigene Families in African Swine Fever Virus: Family 110. J. Virol. 1990, 64, 2064–2072. [Google Scholar] [CrossRef]
- Salas, M.L.; Rey-Campos, J.; Almendral, J.M.; Talavera, A.; Vinuela, E. Transcription and Translation Maps of African Swine Fever Virus. Virology 1986, 152, 228–240. [Google Scholar] [CrossRef]
- Brookes, S.M.; Dixon, L.K.; Parkhouse, R.M. Assembly of African Swine Fever Virus: Quantitative Ultrastructural Analysis in Vitro and in Vivo. Virology 1996, 224, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rouiller, I.; Brookes, S.M.; Hyatt, A.D.; Windsor, M.; Wileman, T. African Swine Fever Virus Is Wrapped by the Endoplasmic Reticulum. J. Virol. 1998, 72, 2373–2387. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, C.; Brookes, S.M.; Wileman, T. Biochemical Requirements of Virus Wrapping by the Endoplasmic Reticulum: Involvement of Atp and Endoplasmic Reticulum Calcium Store during Envelopment of African Swine Fever Virus. J. Virol. 2000, 74, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.; Salas, M.L.; Rodriguez, J.M. African Swine Fever Virus Polyprotein Pp62 Is Essential for Viral Core Development. J. Virol. 2010, 84, 176–187. [Google Scholar] [CrossRef]
- Jouvenet, N.; Monaghan, P.; Way, M.; Wileman, T. Transport of African Swine Fever Virus from Assembly Sites to the Plasma Membrane Is Dependent on Microtubules and Conventional Kinesin. J. Virol. 2004, 78, 7990–8001. [Google Scholar] [CrossRef]
- Schulz, K.; Staubach, C.; Blome, S. African and Classical Swine Fever: Similarities, Differences and Epidemiological Consequences. Vet. Res. 2017, 48, 84. [Google Scholar] [CrossRef]
- Mighell, E.; Ward, M.P. African Swine Fever Spread across Asia, 2018–2019. Transbound. Emerg. Dis. 2021, 68, 2722–2732. [Google Scholar] [CrossRef]
- Nurmoja, I.; Petrov, A.; Breidenstein, C.; Zani, L.; Forth, J.H.; Beer, M.; Kristian, M.; Viltrop, A.; Blome, S. Biological Characterization of African Swine Fever Virus Genotype Ii Strains from North-Eastern Estonia in European Wild Boar. Transbound. Emerg. Dis. 2017, 64, 2034–2041. [Google Scholar] [CrossRef]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernandez-Pinero, J.; Sanchez-Vizcaino, J.M. Gaps in African Swine Fever: Analysis and Priorities. Transbound. Emerg. Dis. 2018, 65 (Suppl. 1), 235–247. [Google Scholar] [CrossRef]
- Gallardo, C.; Sánchez, E.G.; Pérez-Núñez, D.; Nogal, M.; de León, P.; Carrascosa, Á.L.; Nieto, R.; Soler, A.; Arias, M.L.; Revilla, Y. African Swine Fever Virus (ASFV) Protection Mediated by Nh/P68 and Nh/P68 Recombinant Live-Attenuated Viruses. Vaccine 2018, 36, 2694–2704. [Google Scholar] [CrossRef]
- Zsak, L.; Caler, E.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Rock, D.L. A Nonessential African Swine Fever Virus Gene Uk Is a Significant Virulence Determinant in Domestic Swine. J. Virol. 1998, 72, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.A.; Darby, A.C.; da Silva, M.; Upton, C.; Radford, A.D.; Dixon, L.K. Genomic Analysis of Highly Virulent Georgia 2007/1 Isolate of African Swine Fever Virus. Emerg. Infect. Dis. 2011, 17, 599–605. [Google Scholar] [CrossRef]
- Zsak, L.; Lu, Z.; Kutish, G.F.; Neilan, J.G.; Rock, D.L. An African Swine Fever Virus Virulence-Associated Gene Nl-S with Similarity to the Herpes Simplex Virus Icp34.5 Gene. J. Virol. 1996, 70, 8865–8871. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.; Abrams, C.; Hernáez, B.; Alcázar, A.; Escribano, J.M.; Dixon, L.; Alonso, C. The Myd116 African Swine Fever Virus Homologue Interacts with the Catalytic Subunit of Protein Phosphatase 1 and Activates Its Phosphatase Activity. J. Virol. 2007, 81, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Keßler, C.; Forth, J.H.; Keil, G.M.; Mettenleiter, T.C.; Blome, S.; Karger, A. The Intracellular Proteome of African Swine Fever Virus. Sci. Rep. 2018, 8, 14714. [Google Scholar] [CrossRef] [PubMed]
- Zsak, L.; Neilan, J.G. Regulation of Apoptosis in African Swine Fever Virus–Infected Macrophages. Sci. World J. 2002, 2, 1186–1195. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of Mgf360 and Mgf505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef]
- Sanford, B.; Holinka, L.G.; O’Donnell, V.; Krug, P.W.; Carlson, J.; Alfano, M.; Carrillo, C.; Wu, P.; Lowe, A.; Risatti, G.R.; et al. Deletion of the Thymidine Kinase Gene Induces Complete Attenuation of the Georgia Isolate of African Swine Fever Virus. Virus Res. 2016, 213, 165–171. [Google Scholar] [CrossRef]
- Masembe, C.; Phan, M.V.T.; Robertson, D.L.; Cotton, M. Increased Resolution of African Swine Fever Virus Genome Patterns Based on Profile Hmms of Protein Domains. Virus Evol. 2020, 6, veaa044. [Google Scholar] [CrossRef]
- Sánchez, E.G.; Quintas, A.; Nogal, M.; Castelló, A.; Revilla, Y. African Swine Fever Virus Controls the Host Transcription and Cellular Machinery of Protein Synthesis. Virus Res. 2013, 173, 58–75. [Google Scholar] [CrossRef]
- Cackett, G.; Matelska, D.; Sykora, M.; Portugal, R.; Malecki, M.; Bahler, J.; Dixon, L.; Werner, F. The African Swine Fever Virus Transcriptome. J. Virol. 2020, 94, e00119-20. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Medina, E.; Vuono, E.A.; Pruitt, S.; Rai, A.; Espinoza, N.; Valladares, A.; Silva, E.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. Deletion of African Swine Fever Virus Histone-Like Protein, A104r from the Georgia Isolate Drastically Reduces Virus Virulence in Domestic Pigs. Viruses 2022, 14, 1112. [Google Scholar] [CrossRef] [PubMed]
- Keita, D.; Heath, L.; Albina, E. Control of African Swine Fever Virus Replication by Small Interfering Rna Targeting the A151r and Vp72 Genes. Antivir. Ther. 2010, 15, 727–736. [Google Scholar] [CrossRef]
- Shi, J.; Liu, W.; Zhang, M.; Sun, J.; Xu, X. The A179l Gene of African Swine Fever Virus Suppresses Virus-Induced Apoptosis but Enhances Necroptosis. Viruses 2021, 13, 2490. [Google Scholar] [CrossRef] [PubMed]
- Granja, A.G.; Perkins, N.D.; Revilla, Y. A238l Inhibits Nf-Atc2, Nf-Kappa B, and C-Jun Activation through a Novel Mechanism Involving Protein Kinase C-Theta-Mediated up-Regulation of the Amino-Terminal Transactivation Domain of P300. J. Immunol. 2008, 180, 2429–2442, Erratum in J. Immunol. 2015, 194, 2032. [Google Scholar] [CrossRef]
- Li, D.; Yang, W.; Li, L.; Li, P.; Ma, Z.; Zhang, J.; Qi, X.; Ren, J.; Ru, Y.; Niu, Q.; et al. African Swine Fever Virus Mgf-505-7r Negatively Regulates Cgas-Sting-Mediated Signaling Pathway. J. Immunol. 2021, 206, 1844–1857. [Google Scholar] [CrossRef]
- Gao, Q.; Yang, Y.L.; Quan, W.P.; Zheng, J.C.; Luo, Y.Z.; Wang, H.; Chen, X.N.; Huang, Z.; Chen, X.J.; Xu, R.D.; et al. The African Swine Fever Virus with Mgf360 and Mgf505 Deleted Reduces the Apoptosis of Porcine Alveolar Macrophages by Inhibiting the Nf-Kappa B Signaling Pathway and Interleukin-1 Beta. Vaccines 2021, 9, 1371. [Google Scholar] [CrossRef]
- Farlow, J.; Donduashvili, M.; Kokhreidze, M.; Kotorashvili, A.; Vepkhvadze, N.G.; Kotaria, N.; Gulbani, A. Intra-Epidemic Genome Variation in Highly Pathogenic African Swine Fever Virus (ASFV) from the Country of Georgia. Virol. J. 2018, 15, 190. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Krug, P.W.; Gladue, D.P.; Carlson, J.; Sanford, B.; Alfano, M.; Kramer, E.; Lu, Z.Q.; Arzt, J.; et al. African Swine Fever Virus Georgia 2007 with a Deletion of Virulence-Associated Gene 9gl (B119l), When Administered at Low Doses, Leads to Virus Attenuation in Swine and Induces an Effective Protection against Homologous Challenge. J. Virol. 2015, 89, 8556–8566. [Google Scholar] [CrossRef]
- Barrado-Gil, L.; Galindo, I.; Martinez-Alonso, D.; Viedma, S.; Alonso, C. The Ubiquitin-Proteasome System Is Required for African Swine Fever Replication. PLoS ONE 2017, 12, e0189741. [Google Scholar] [CrossRef]
- Kinyanyi, D.; Obiero, G.; Obiero, G.F.O.; Amwayi, P.; Mwaniki, S.; Wamalwa, M. In Silico Structural and Functional Prediction of African Swine Fever Virus Protein-B263r Reveals Features of a Tata-Binding Protein. PeerJ 2018, 6, e4396. [Google Scholar] [CrossRef] [PubMed]
- Alejo, A.; Yanez, R.J.; Rodriguez, J.M.; Vinuela, E.; Salas, M.L. African Swine Fever Virus Trans-Prenyltransferase. J. Biol. Chem. 1997, 272, 9417–9423. [Google Scholar] [CrossRef] [PubMed]
- Onzere, C.K.; Bastos, A.D.; Okoth, E.A.; Lichoti, J.K.; Bochere, E.N.; Owido, M.G.; Ndambuki, G.; Bronsvoort, M.; Bishop, R.P. Multi-Locus Sequence Typing of African Swine Fever Viruses from Endemic Regions of Kenya and Eastern Uganda (2011–2013) Reveals Rapid B602l Central Variable Region Evolution. Virus Genes 2018, 54, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ma, B.; Qian, N.; Zhang, F.; Tan, X.; Lei, J.; Xiang, Y. Structure of the African Swine Fever Virus Major Capsid Protein P72. Cell Res. 2019, 29, 953–955. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Shao, N.; Chen, J.W.; Qi, W.B.; Li, Y.; Liu, P.; Chen, Y.J.; Bian, S.Y.; Zhang, Y.; Tao, S.C. Multiplex and Visual Detection of African Swine Fever Virus (ASFV) Based on Hive-Chip and Direct Loop-Mediated Isothermal Amplification. Anal. Chim. Acta 2020, 1140, 30–40. [Google Scholar] [CrossRef]
- Angulo, A.; Alcami, A.; Vinuela, E. Virus-Host Interactions in African Swine Fever: The Attachment to Cellular Receptors. Arch. Virol. Suppl. 1993, 7, 169–183. [Google Scholar]
- Ramirez-Medina, E.; Vuono, E.A.; Rai, A.; Pruitt, S.; Silva, E.; Velazquez-Salinas, L.; Zhu, J.; Borca, M.V.; Gladue, D.P. The C962r Orf of African Swine Fever Strain Georgia Is Non-Essential and Not Required for Virulence in Swine. Viruses 2020, 12, 676. [Google Scholar] [CrossRef]
- Hubner, A.; Petersen, B.; Keil, G.M.; Niemann, H.; Mettenleiter, T.C.; Fuchs, W. Efficient Inhibition of African Swine Fever Virus Replication by Crispr/Cas9 Targeting of the Viral P30 Gene (Cp204l). Sci. Rep. 2018, 8, 1449. [Google Scholar] [CrossRef]
- Chen, L.Y.; Zhang, X.H.; Shao, G.M.; Shao, Y.Y.; Hu, Z.Z.; Feng, K.Y.; Xie, Z.; Li, H.X.; Chen, W.G.; Lin, W.C.; et al. Construction and Evaluation of Recombinant Pseudorabies Virus Expressing African Swine Fever Virus Antigen Genes. Front. Vet. Sci. 2022, 9, 832255. [Google Scholar] [CrossRef]
- Yanez, R.J.; Rodriguez, J.M.; Boursnell, M.; Rodriguez, J.F.; Vinuela, E. Two Putative African Swine Fever Virus Helicases Similar to Yeast ‘Deah’ Pre-Mrna Processing Proteins and Vaccinia Virus Atpases D11l and D6r. Gene 1993, 134, 161–174. [Google Scholar] [CrossRef]
- Muturi, E.; Meng, F.; Liu, H.; Jiang, M.; Wei, H.; Yang, H. Comprehensive Analysis of G-Quadruplexes in African Swine Fever Virus Genome Reveals Potential Antiviral Targets by G-Quadruplex Stabilizers. Front. Microbiol. 2021, 12, 798431. [Google Scholar] [CrossRef] [PubMed]
- Quintas, A.; Perez-Nunez, D.; Sanchez, E.G.; Nogal, M.L.; Hentze, M.W.; Castello, A.; Revilla, Y. Characterization of the African Swine Fever Virus Decapping Enzyme during Infection. J. Virol. 2017, 91, e00990-17. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, M.; García-Escudero, R.; Alejo, A.; Viñuela, E.; Salas, M.L.; Salas, J. African Swine Fever Virus Dutpase Is a Highly Specific Enzyme Required for Efficient Replication in Swine Macrophages. J. Virol. 1999, 73, 8934–8943. [Google Scholar] [CrossRef]
- Petrovan, V.; Murgia, M.V.; Wu, P.; Lowe, A.D.; Jia, W.; Rowland, R.R.R. Epitope Mapping of African Swine Fever Virus (ASFV) Structural Protein, P54. Virus Res. 2020, 279, 197871. [Google Scholar] [CrossRef]
- Kollnberger, S.D.; Gutierrez-Castaneda, B.; Foster-Cuevas, M.; Corteyn, A.; Parkhouse, R.M.E. Identification of the Principal Serological Immunodeterminants of African Swine Fever Virus by Screening a Virus Cdna Library with Antibody. J. Gen. Virol. 2002, 83, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, T.; Alejo, A.; Rodriguez, J.M.; Hernaez, B.; Guerra, M.; Fraile-Ramos, A.; Andres, G. African Swine Fever Virus Protein Pe199l Mediates Virus Entry by Enabling Membrane Fusion and Core Penetration. Mbio 2020, 11, e00789-20. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Almazan, F.; Bustos, M.J.; Vinuela, E.; Carrascosa, A.L. African Swine Fever Virus Ep153r Open Reading Frame Encodes a Glycoprotein Involved in the Hemadsorption of Infected Cells. Virology 2000, 266, 340–351. [Google Scholar] [CrossRef]
- Rodriguez, J.M.; Yanez, R.J.; Almazan, F.; Vinuela, E.; Rodriguez, J.F. African Swine Fever Virus Encodes a Cd2 Homolog Responsible for the Adhesion of Erythrocytes to Infected Cells. J. Virol. 1993, 67, 5312–5320. [Google Scholar] [CrossRef]
- Rodriguez, J.M.; Salas, M.L.; Viñuela, E. Genes Homologous to Ubiquitin-Conjugating Proteins and Eukaryotic Transcription Factor Sii in African Swine Fever Virus. Virology 1992, 186, 40–52. [Google Scholar] [CrossRef]
- Nunes-Correia, I.; Rodriguez, J.M.; Eulalio, A.; Carvalho, A.L.; Citovsky, V.; Simoes, S.; Faro, C.; Salas, M.L.; de Lima, M.C.P. African Swine Fever Virus P10 Protein Exhibits Nuclear Import Capacity and Accumulates in the Nucleus during Viral Infection. Vet. Microbiol. 2008, 130, 47–59. [Google Scholar] [CrossRef]
- Bosch-Camos, L.; Lopez, E.; Collado, J.; Navas, M.J.; Blanco-Fuertes, M.; Pina-Pedrero, S.; Accensi, F.; Salas, M.L.; Mundt, E.; Nikolin, V.; et al. M448r and Mgf505-7r: Two African Swine Fever Virus Antigens Commonly Recognized by ASFV-Specific T-Cells and with Protective Potential. Vaccines 2021, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, P.; Liu, H.; Feng, T.; Yang, W.; Ru, Y.; Li, P.; Qi, X.; Shi, Z.; Zheng, H. A Qp509l/Qp383r-Deleted African Swine Fever Virus Is Highly Attenuated in Swine but Does Not Confer Protection against Parental Virus Challenge. J. Virol. 2022, 96, e01500-21. [Google Scholar] [CrossRef] [PubMed]
- Yanez, R.J.; Rodriguez, J.M.; Rodriguez, J.F.; Salas, M.L.; Vinuela, E. African Swine Fever Virus Thymidylate Kinase Gene: Sequence and Transcriptional Mapping. J. Gen. Virol. 1993, 74 Pt 8, 1633–1638. [Google Scholar] [CrossRef]
- Reis, A.L.; Parkhouse, R.M.E.; Penedos, A.R.; Martins, C.; Leitao, A. Systematic Analysis of Longitudinal Serological Responses of Pigs Infected Experimentally with African Swine Fever Virus. J. Gen. Virol. 2007, 88 Pt 9, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.; Martins, C.; Ferreira, F.; Leitao, A. African Swine Fever Virus Orf P1192r Codes for a Functional Type Ii DNA Topoisomerase. Virology 2015, 474, 82–93. [Google Scholar] [CrossRef]
- Mazur-Panasiuk, N.; Walczak, M.; Juszkiewicz, M.; Wozniakowski, G. The Spillover of African Swine Fever in Western Poland Revealed Its Estimated Origin on the Basis of O174l, K145r, Mgf 505-5r and Igr I73r/I329l Genomic Sequences. Viruses 2020, 12, 1094. [Google Scholar] [CrossRef]
- Song, J.; Li, K.; Li, T.; Zhao, G.; Zhou, S.; Li, H.; Li, J.; Weng, C. Screening of Prrsv- and ASFV-Encoded Proteins Involved in the Inflammatory Response Using a Porcine Igluc Reporter. J. Virol. Methods 2020, 285, 113958. [Google Scholar] [CrossRef]
- Redrejo-Rodriguez, M.; Garcia-Escudero, R.; Yanez-Munoz, R.J.; Salas, M.L.; Salas, J. African Swine Fever Virus Protein Pe296r Is a DNA Repair Apurinic/Apyrimidinic Endonuclease Required for Virus Growth in Swine Macrophages. J. Virol. 2006, 80, 4847–4857. [Google Scholar] [CrossRef]
- Yáñez, R.J.; Boursnell, M.; Nogal, M.L.; Yuste, L.; Viñuela, E. African Swine Fever Virus Encodes Two Genes Which Share Significant Homology with the Two Largest Subunits of DNA-Dependent Rna Polymerases. Nucleic Acids Res. 1993, 21, 2423–2427. [Google Scholar] [CrossRef]
- Pastorino, L.; Cusano, R.; Bruno, W.; Lantieri, F.; Origone, P.; Barile, M.; Gliori, S.; Shepherd, G.A.; Sturm, R.A.; Bianchi-Scarra, G. Novel Mc1r Variants in Ligurian Melanoma Patients and Controls. Hum. Mutat. 2004, 24, 103. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.A.; Pruitt, S.; Rai, A.; Espinoza, N.; Velazquez-Salinas, L.; Gladue, D.P.; Borca, M.V. Evaluation of an ASFV Rna Helicase Gene A859l for Virus Replication and Swine Virulence. Viruses 2022, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Teramo, A.; Gasparini, V.R.; Huuhtanen, J.; Kim, D.; Theodoropoulos, J.; Schiavoni, G.; Barila, G.; Vicenzetto, C.; Calabretto, G.; et al. Identification of Novel Stat5b Mutations and Characterization of Tcrbeta Signatures in Cd4+ T-Cell Large Granular Lymphocyte Leukemia. Blood Cancer J. 2022, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.B.; Frouco, G.; Martins, C.; Ferreira, F. The Qp509l and Q706l Superfamily Ii Rna Helicases of African Swine Fever Virus Are Required for Viral Replication, Having Non-Redundant Activities. Emerg. Microbes Infect. 2019, 8, 291–302. [Google Scholar] [CrossRef]
- Rodriguez, J.M.; Salas, M.L.; Vinuela, E. Intermediate Class of Mrnas in African Swine Fever Virus. J. Virol. 1996, 70, 8584–8589. [Google Scholar] [CrossRef] [PubMed]
- Eaton, H.E.; Kobayashi, T.; Dermody, T.S.; Johnston, R.N.; Jais, P.H.; Shmulevitz, M. African Swine Fever Virus Np868r Capping Enzyme Promotes Reovirus Rescue during Reverse Genetics by Promoting Reovirus Protein Expression, Virion Assembly, and Rna Incorporation into Infectious Virions. J. Virol. 2017, 91, e02416-16. [Google Scholar] [CrossRef]
- Dodantenna, N.; Ranathunga, L.; Chathuranga, W.A.G.; Weerawardhana, A.; Cha, J.W.; Subasinghe, A.; Gamage, N.; Haluwana, D.K.; Kim, Y.; Jheong, W.; et al. African Swine Fever Virus Ep364r and C129r Target Cyclic Gmp-Amp to Inhibit the Cgas-Sting Signaling Pathway. J. Virol. 2022, e01022-22. [Google Scholar] [CrossRef]
- Leitão, A.; Malur, A.; Cartaxeiro, C.; Vasco, G.; Cruz, B.; Cornelis, P.; Martins, C.L. Bacterial Lipoprotein Based Expression Vectors as Tools for the Characterisation of African Swine Fever Virus (ASFV) Antigens. Arch. Virol. 2000, 145, 1639–1657. [Google Scholar] [CrossRef]
- Moustaq, L.; Smaczynska-de Rooij, I.I.; Palmer, S.E.; Marklew, C.J.; Ayscough, K.R. Insights into Dynamin-Associated Disorders through Analysis of Equivalent Mutations in the Yeast Dynamin Vps1. J. Microbial. Cell 2016, 3, 147–158. [Google Scholar] [CrossRef]
- Freitas, F.B.; Frouco, G.; Martins, C.; Ferreira, F. African Swine Fever Virus Encodes for an E2-Ubiquitin Conjugating Enzyme That Is Mono- and Di-Ubiquitinated and Required for Viral Replication Cycle. Sci. Rep. 2018, 8, 3471. [Google Scholar] [CrossRef]
- Zhang, F.; Moon, A.; Childs, K.; Goodbourn, S.; Dixon, L.K. The African Swine Fever Virus Dp71l Protein Recruits the Protein Phosphatase 1 Catalytic Subunit to Dephosphorylate eIF2α and Inhibits Chop Induction but Is Dispensable for These Activities during Virus Infection. J. Virol. 2010, 84, 10681–10689. [Google Scholar] [CrossRef]
- Vuono, E.A.; Ramirez-Medina, E.; Pruitt, S.; Rai, A.; Espinoza, N.; Velazquez-Salinas, L.; Gladue, D.P.; Borca, M.V. Evaluation of the Function of the ASFV Kp177r Gene, Encoding for Structural Protein P22, in the Process of Virus Replication and in Swine Virulence. Viruses 2021, 13, 986. [Google Scholar] [CrossRef] [PubMed]
- Gladue, D.P.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V. Deletion of the A137r Gene from the Pandemic Strain of African Swine Fever Virus Attenuates the Strain and Offers Protection against the Virulent Pandemic Virus. J. Virol. 2021, 95, e01139-21. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Li, K.; Qiao, S.; Chen, X.X.; Deng, R.; Zhang, G. Development and Evaluation of Duplex Taqman Real-Time Pcr Assay for Detection and Differentiation of Wide-Type and Mgf505-2r Gene-Deleted African Swine Fever Viruses. BMC Vet. Res. 2020, 16, 428. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zhang, L.H.; Lin, Y.; Cai, Y.F.; Zou, Y.W.; Hao, Z.Y.; Luo, Z.H.; Wang, N.D.; Deng, Z.B.; Yang, Y.; et al. Development and Preliminary Testing of a Probe-Based Duplex Real-Time Pcr Assay for the Detection of African Swine Fever Virus. Mol. Cell. Probes 2021, 59, 101764. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Castaneda, B.; Reis, A.L.; Corteyn, A.; Parkhouse, R.M.; Kollnberger, S. Expression, Cellular Localization and Antibody Responses of the African Swine Fever Virus Genes B602l and K205r. Arch. Virol. 2008, 153, 2303–2306. [Google Scholar] [CrossRef]
- Lokhandwala, S.; Waghela, S.D.; Bray, J.; Sangewar, N.; Charendoff, C.; Martin, C.L.; Hassan, W.S.; Koynarski, T.; Gabbert, L.; Burrage, T.G.; et al. Adenovirus-Vectored Novel African Swine Fever Virus Antigens Elicit Robust Immune Responses in Swine. PLoS ONE 2017, 12, e0177007. [Google Scholar] [CrossRef]
- Zhao, G.; Li, T.; Liu, X.; Zhang, T.; Zhang, Z.; Kang, L.; Song, J.; Zhou, S.; Chen, X.; Wang, X.; et al. African Swine Fever Virus Cysteine Protease Ps273r Inhibits Pyroptosis by Noncanonically Cleaving Gasdermin D. J. Biol. Chem. 2022, 298, 101480. [Google Scholar] [CrossRef]
- Vuono, E.; Ramirez-Medina, E.; Silva, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Valladares, A.; Velazquez-Salinas, L.; Gladue, D.P.; Borca, M.V. Deletion of the H108r Gene Reduces Virulence of the Pandemic Eurasia Strain of African Swine Fever Virus with Surviving Animals Being Protected against Virulent Challenge. J. Virol. 2022, 96, e00545-22. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, Z.; Feng, T.; Ma, Z.; Xue, Q.; Wu, P.; Li, P.; Li, S.; Yang, F.; Cao, W.; et al. African Swine Fever Virus E120r Protein Inhibits Interferon Beta Production by Interacting with Irf3 to Block Its Activation. J. Virol. 2021, 95, e00824-21. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, W.; Wen, Y.; Niu, Q.; Yang, J.; Guan, G.; Yin, H.; Zheng, H.; Li, D.; Liu, Z. The E248r Protein of African Swine Fever Virus Inhibits the Cgas-Sting-Mediated Innate Immunity. Sheng Wu Gong Cheng Xue Bao 2022, 38, 1837–1846. [Google Scholar]
- Powell, P.P.; Dixon, L.K.; Parkhouse, R.M. An Ikappab Homolog Encoded by African Swine Fever Virus Provides a Novel Mechanism for Downregulation of Proinflammatory Cytokine Responses in Host Macrophages. J. Virol. 1996, 70, 8527–8533. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Abrams, C.C.; Bowick, G.; Goatley, L.C.; Kay-Jackson, P.C.; Chapman, D.; Liverani, E.; Nix, R.; Silk, R.; Zhang, F. African Swine Fever Virus Proteins Involved in Evading Host Defence Systems. Vet. Immunol. Immunopathol. 2004, 100, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Ventura, S.; Parkhouse, R.M. Identification and Utility of Innate Immune System Evasion Mechanisms of ASFV. Virus Res. 2013, 173, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.L.; Goatley, L.C.; Jabbar, T.; Sanchez-Cordon, P.J.; Netherton, C.L.; Chapman, D.A.G.; Dixon, L.K. Deletion of the African Swine Fever Virus Gene Dp148r Does Not Reduce Virus Replication in Culture but Reduces Virus Virulence in Pigs and Induces High Levels of Protection against Challenge. J. Virol. 2017, 91, e01428-17. [Google Scholar] [CrossRef] [PubMed]
- Whittall, J.T.; Parkhouse, R.M. Changes in Swine Macrophage Phenotype after Infection with African Swine Fever Virus: Cytokine Production and Responsiveness to Interferon-Gamma and Lipopolysaccharide. Immunology 1997, 91, 444–449. [Google Scholar] [CrossRef]
- Borca, M.V.; Carrillo, C.; Zsak, L.; Laegreid, W.W.; Kutish, G.F.; Neilan, J.G.; Burrage, T.G.; Rock, D.L. Deletion of a Cd2-Like Gene, 8-Dr, from African Swine Fever Virus Affects Viral Infection in Domestic Swine. J. Virol. 1998, 72, 2881–2889. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Wu, Y.; Chen, H.; Zhang, S.; Li, J.; Xin, T.; Jia, H.; Hou, S.; Jiang, Y.; et al. Inhibition of Cgas-Sting-Tbk1 Signaling Pathway by Dp96r of ASFV China 2018/1. Biochem. Biophys. Res. Commun. 2018, 506, 437–443. [Google Scholar] [CrossRef]
- García-Belmonte, R.; Pérez-Núñez, D.; Pittau, M.; Richt, J.A.; Revilla, Y. African Swine Fever Virus Armenia/07 Virulent Strain Controls Interferon Beta Production through the Cgas-Sting Pathway. J. Virol. 2019, 93, e02298-18. [Google Scholar] [CrossRef]
- Gómez-Villamandos, J.C.; Bautista, M.J.; Sánchez-Cordón, P.J.; Carrasco, L. Pathology of African Swine Fever: The Role of Monocyte-Macrophage. Virus Res. 2013, 173, 140–149. [Google Scholar] [CrossRef]
- Zakaryan, H.; Cholakyans, V.; Simonyan, L.; Misakyan, A.; Karalova, E.; Chavushyan, A.; Karalyan, Z. A Study of Lymphoid Organs and Serum Proinflammatory Cytokines in Pigs Infected with African Swine Fever Virus Genotype Ii. Arch. Virol. 2015, 160, 1407–1414. [Google Scholar] [CrossRef]
- Reis, A.L.; Goatley, L.C.; Jabbar, T.; Lopez, E.; Rathakrishnan, A.; Dixon, L.K. Deletion of the Gene for the Type I Interferon Inhibitor I329l from the Attenuated African Swine Fever Virus Ourt88/3 Strain Reduces Protection Induced in Pigs. Vaccines 2020, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, V.L.; Almeida, S.C.P.; Soares, H.R.; Crespo, A.; Marshall-Clarke, S.; Parkhouse, R.M.E. A Novel Tlr3 Inhibitor Encoded by African Swine Fever Virus (ASFV). Arch. Virol. 2011, 156, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Nogal, M.L.; de Buitrago, G.G.; Rodriguez, C.; Cubelos, B.; Carrascosa, A.L.; Salas, M.L.; Revilla, Y. African Swine Fever Virus Iap Homologue Inhibits Caspase Activation and Promotes Cell Survival in Mammalian Cells. J. Virol. 2001, 75, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Granja, A.G.; Nogal, M.L.; Hurtado, C.; Salas, J.; Salas, M.L.; Carrascosa, A.L.; Revilla, Y. Modulation of P53 Cellular Function and Cell Death by African Swine Fever Virus. J. Virol. 2004, 78, 7165–7174. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, B.; Diaz-Gil, G.; Garcia-Gallo, M.; Quetglas, J.I.; Rodriguez-Crespo, I.; Dixon, L.; Escribano, J.M.; Alonso, C. The African Swine Fever Virus Dynein-Binding Protein P54 Induces Infected Cell Apoptosis. FEBS Lett. 2004, 569, 224–228. [Google Scholar] [CrossRef]
- Barber, C.; Netherton, C.; Goatley, L.; Moon, A.; Goodbourn, S.; Dixon, L. Identification of Residues within the African Swine Fever Virus Dp71l Protein Required for Dephosphorylation of Translation Initiation Factor Eif2 Alpha and Inhibiting Activation of Pro-Apoptotic Chop. Virology 2017, 504, 107–113. [Google Scholar] [CrossRef]
- Wang, C.Y.; Mayo, M.W.; Korneluk, R.G.; Goeddel, D.V.; Baldwin, A.S., Jr. Nf-Kappab Antiapoptosis: Induction of Traf1 and Traf2 and C-Iap1 and C-Iap2 to Suppress Caspase-8 Activation. Science 1998, 281, 1680–1683. [Google Scholar] [CrossRef]
- Dixon, L.K.; Sánchez-Cordón, P.J.; Galindo, I.; Alonso, C. Investigations of Pro- and Anti-Apoptotic Factors Affecting African Swine Fever Virus Replication and Pathogenesis. Viruses 2017, 9, 241. [Google Scholar] [CrossRef]
- Salguero, F.J.; Sanchez-Cordon, P.J.; Sierra, M.A.; Jover, A.; Nunez, A.; Gomez-Villamandos, J.C. Apoptosis of Thymocytes in Experimental African Swine Fever Virus Infection. Histol. Histopathol. 2004, 19, 77–84. [Google Scholar]
- Banjara, S.; Caria, S.; Dixon, L.K.; Hinds, M.G.; Kvansakula, M. Structural Insight into African Swine Fever Virus A179l-Mediated Inhibition of Apoptosis. J. Virol. 2017, 91, e02228-16. [Google Scholar] [CrossRef]
- Salguero, F.J.; Sánchez-Cordón, P.J.; Núñez, A.; de Marco, M.F.; Gómez-Villamandos, J.C. Proinflammatory Cytokines Induce Lymphocyte Apoptosis in Acute African Swine Fever Infection. J. Comp. Pathol. 2005, 132, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Hernáez, B.; Muñoz-Moreno, R.; Cuesta-Geijo, M.A.; Dalmau-Mena, I.; Alonso, C. The Atf6 Branch of Unfolded Protein Response and Apoptosis Are Activated to Promote African Swine Fever Virus Infection. Cell Death Dis. 2012, 3, e341. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, A.L.; Bustos, M.J.; Nogal, M.L.; de Buitrago, G.G.; Revilla, Y. Apoptosis Induced in an Early Step of African Swine Fever Virus Entry into Vero Cells Does Not Require Virus Replication. Virology 2002, 294, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Netherton, C.L.; Parsley, J.C.; Wileman, T. African Swine Fever Virus Inhibits Induction of the Stress-Induced Proapoptotic Transcription Factor Chop/Gadd153. J. Virol. 2004, 78, 10825–10828. [Google Scholar] [CrossRef] [PubMed]
- Banjara, S.; Shimmon, G.L.; Dixon, L.K.; Netherton, C.L.; Hinds, M.G.; Kvansakul, M. Crystal Structure of African Swine Fever Virus A179l with the Autophagy Regulator Beclin. Viruses 2019, 11, 789. [Google Scholar] [CrossRef]
- Neilan, J.G.; Lu, Z.; Afonso, C.L.; Kutish, G.F.; Sussman, M.D.; Rock, D.L. An African Swine Fever Virus Gene with Similarity to the Proto-Oncogene Bcl-2 and the Epstein-Barr Virus Gene Bhrf1. J. Virol. 1993, 67, 4391–4394. [Google Scholar] [CrossRef]
- Rodríguez, C.I.; Nogal, M.L.; Carrascosa, A.L.; Salas, M.L.; Fresno, M.; Revilla, Y. African Swine Fever Virus Iap-Like Protein Induces the Activation of Nuclear Factor Kappa B. J. Virol. 2002, 76, 3936–3942. [Google Scholar] [CrossRef]
- Hurtado, C.; Granja, A.G.; Bustos, M.J.; Nogal, M.L.; de Buitrago, G.G.; de Yebenes, V.G.; Salas, M.L.; Revilla, Y.; Carrascosa, A.L. The C-Type Lectin Homologue Gene (Ep153r) of African Swine Fever Virus Inhibits Apoptosis Both in Virus Infection and in Heterologous Expression. Virology 2004, 326, 160–170. [Google Scholar] [CrossRef]
- Li, J.; Song, J.; Kang, L.; Huang, L.; Zhou, S.; Hu, L.; Zheng, J.; Li, C.; Zhang, X.; He, X.; et al. Pmgf505-7r Determines Pathogenicity of African Swine Fever Virus Infection by Inhibiting Il-1beta and Type I Ifn Production. PLoS Pathog. 2021, 17, e1009733. [Google Scholar] [CrossRef]
- Yang, B.; Shen, C.C.; Zhang, D.J.; Zhang, T.; Shi, X.J.; Yang, J.K.; Hao, Y.; Zhao, D.S.; Cui, H.M.; Yuan, X.G.; et al. Mechanism of Interaction between Virus and Host Is Inferred from the Changes of Gene Expression in Macrophages Infected with African Swine Fever Virus Cn/Gs/2018 Strain. Virol. J. 2021, 18, 170. [Google Scholar] [CrossRef]
- Mitter, S.K.; Song, C.J.; Qi, X.P.; Mao, H.Y.; Rao, H.; Akin, D.; Lewin, A.; Grant, M.; Dunn, W.; Ding, J.D.; et al. Dysregulated Autophagy in the Rpe Is Associated with Increased Susceptibility to Oxidative Stress and Amd. Autophagy 2014, 10, 1989–2005. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, K. Subversion of the Cellular Autophagy Pathway by Viruses. Autophagy Infect. Immun. 2009, 335, 323–333. [Google Scholar]
- Jackson, W.T.; Giddings, T.H.; Taylor, M.P.; Mulinyawe, S.; Rabinovitch, M.; Kopito, R.R.; Kirkegaard, K. Subversion of Cellular Autophagosomal Machinery by RNA Viruses. PLoS Biol. 2005, 3, e156. [Google Scholar] [CrossRef]
- Pei, J.; Zhao, M.; Ye, Z.; Gou, H.; Wang, J.; Yi, L.; Dong, X.; Liu, W.; Luo, Y.; Liao, M.; et al. Autophagy Enhances the Replication of Classical Swine Fever Virus in Vitro. Autophagy 2014, 10, 93–110. [Google Scholar] [CrossRef]
- Dash, S.; Chava, S.; Aydin, Y.; Chandra, P.K.; Ferraris, P.; Chen, W.N.; Balart, L.A.; Wu, T.; Garry, R.F. Hepatitis C Virus Infection Induces Autophagy as a Prosurvival Mechanism to Alleviate Hepatic Er-Stress Response. Viruses 2016, 8, 150. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Cui, L.P.; Zhu, E.P.; Zhou, W.D.; Wang, Q.X.; Wu, X.P.; Wu, B.C.; Huang, Y.F.; Liu, H.J. Muscovy Duck Reovirus Sigma Ns Protein Triggers Autophagy Enhancing Virus Replication. Virol. J. 2017, 14, 53. [Google Scholar] [CrossRef]
- Hernaez, B.; Cabezas, M.; Munoz-Moreno, R.; Galindo, I.; Cuesta-Geijo, M.A.; Alonso, C. A179l, a New Viral Bcl2 Homolog Targeting Beclin 1 Autophagy Related Protein. Curr. Mol. Med. 2013, 13, 305–316. [Google Scholar] [CrossRef]
- Revilla, Y.; Cebrian, A.; Baixeras, E.; Martinez, C.; Vinuela, E.; Salas, M.L. Inhibition of Apoptosis by the African Swine Fever Virus Bcl-2 Homologue: Role of the Bh1 Domain. Virology 1997, 228, 400–404. [Google Scholar] [CrossRef]
- Shimmon, G.L.; Hui, J.Y.K.; Wileman, T.E.; Netherton, C.L. Autophagy Impairment by African Swine Fever Virus. J. Gen. Virol. 2021, 102, 001637. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, X.H.; Nie, Y.; Li, H.X.; Chen, W.G.; Lin, W.C.; Chen, F.; Xie, Q.M. African Swine Fever Virus Protein E199l Promotes Cell Autophagy through the Interaction of Pycr2. Virol. Sin. 2021, 36, 196–206. [Google Scholar] [CrossRef]
- Davies, K.; Goatley, L.C.; Guinat, C.; Netherton, C.L.; Gubbins, S.; Dixon, L.K.; Reis, A.L. Survival of African Swine Fever Virus in Excretions from Pigs Experimentally Infected with the Georgia 2007/1 Isolate. Transbound. Emerg. Dis. 2017, 64, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Panasiuk, N.; Woźniakowski, G. Natural Inactivation of African Swine Fever Virus in Tissues: Influence of Temperature and Environmental Conditions on Virus Survival. Vet. Microbiol. 2020, 242, 108609. [Google Scholar] [CrossRef] [PubMed]
- Farez, S.; Morley, R.S. Potential Animal Health Hazards of Pork and Pork Products. Rev. Sci. Tech. 1997, 16, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Petrini, S.; Feliziani, F.; Casciari, C.; Giammarioli, M.; Torresi, C.; de Mia, G.M. Survival of African Swine Fever Virus (ASFV) in Various Traditional Italian Dry-Cured Meat Products. Prev. Vet. Med. 2019, 162, 126–130. [Google Scholar] [CrossRef]
- Zani, L.; Masiulis, M.; Bušauskas, P.; Dietze, K.; Pridotkas, G.; Globig, A.; Blome, S.; Mettenleiter, T.; Depner, K.; Karvelienė, B. African Swine Fever Virus Survival in Buried Wild Boar Carcasses. Transbound. Emerg. Dis. 2020, 67, 2086–2092. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African Swine Fever—A Review of Current Knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Olesen, A.S.; Hansen, M.F.; Rasmussen, T.B.; Belsham, G.J.; Bødker, R.; Bøtner, A. Survival and Localization of African Swine Fever Virus in Stable Flies (Stomoxys Calcitrans) after Feeding on Viremic Blood Using a Membrane Feeder. Vet. Microbiol. 2018, 222, 25–29. [Google Scholar] [CrossRef]
- Karalyan, Z.; Avetisyan, A.; Avagyan, H.; Ghazaryan, H.; Vardanyan, T.; Manukyan, A.; Semerjyan, A.; Voskanyan, H. Presence and Survival of African Swine Fever Virus in Leeches. Vet. Microbiol. 2019, 237, 108421. [Google Scholar] [CrossRef]
- Penrith, M.L.; Guberti, V.; Depner, K.; Lubroth, J. Preparation of African Swine Fever Contingency Plans, 8th ed.; FAO: Rome, Italy, 2009. [Google Scholar]
- Gallina, L.; Scagliarini, A. Virucidal Efficacy of Common Disinfectants against Orf Virus. Vet. Rec. 2010, 166, 725. [Google Scholar] [CrossRef]
- World Organisation for Animal Health—OIE. Terrestrial Animal Health Code, General Recommendations on Disinfection and Disinsectisation; World Organisation for Animal Health (OIE): Paris, France, 2011; Volume 1, p. 169. [Google Scholar]
- Juszkiewicz, M.; Walczak, M.; Woźniakowski, G. Characteristics of Selected Active Substances Used in Disinfectants and Their Virucidal Activity against ASFV. J. Vet. Res. 2019, 63, 17–25. [Google Scholar] [CrossRef]
- Shirai, J.; Kanno, T.; Tsuchiya, Y.; Mitsubayashi, S.; Seki, R. Effects of Chlorine, Iodine, and Quaternary Ammonium Compound Disinfectants on Several Exotic Disease Viruses. J. Vet. Med. Sci. 2000, 62, 85–92. [Google Scholar] [CrossRef]
- Fischer, M.; Mohnke, M.; Probst, C.; Pikalo, J.; Conraths, F.J.; Beer, M.; Blome, S. Stability of African Swine Fever Virus on Heat-Treated Field Crops. Transbound. Emerg. Dis. 2020, 67, 2318–2323. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, Y.; Wang, W.; Sun, Y.; Zhang, J.; Fatima, M.; Jia, X.; Qiu, H.J. Efficient Inactivation of African Swine Fever Virus by Ozonized Water. Vet. Microbiol. 2020, 247, 108796. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health—OIE. Technical Disease Card for African Swine Fever; World Organisation for Animal Health (OIE): Paris, France, 2009. [Google Scholar]
- Franco-Martinez, L.; Beer, M.; Martinez-Subiela, S.; Garcia-Manzanilla, E.; Blome, S.; Carrau, T. Impact of ASFV Detergent Inactivation on Biomarkers in Serum and Saliva Samples. Pathogens 2022, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Garigliany, M.; Desmecht, D.; Tignon, M.; Cassart, D.; Lesenfant, C.; Paternostre, J.; Volpe, R.; Cay, A.B.; van den Berg, T.; Linden, A. Phylogeographic Analysis of African Swine Fever Virus, Western Europe, 2018. Emerg. Infect. Dis. 2019, 25, 184–186. [Google Scholar] [CrossRef]
- Bastos, A.D.; Penrith, M.L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.; Thomson, R.G. Genotyping Field Strains of African Swine Fever Virus by Partial P72 Gene Characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Malogolovkin, A.; Burmakina, G.; Titov, I.; Sereda, A.; Gogin, A.; Baryshnikova, E.; Kolbasov, D. Comparative Analysis of African Swine Fever Virus Genotypes and Serogroups. Emerg. Infect. Dis. 2015, 21, 312–315. [Google Scholar] [CrossRef]
- Frączyk, M.; Woźniakowski, G.; Kowalczyk, A.; Bocian, Ł.; Kozak, E.; Niemczuk, K.; Pejsak, Z. Evolution of African Swine Fever Virus Genes Related to Evasion of Host Immune Response. Vet. Microbiol. 2016, 193, 133–144. [Google Scholar] [CrossRef]
- Forth, J.H.; Tignon, M.; Cay, A.B.; Forth, L.F.; Höper, D.; Blome, S.; Beer, M. Comparative Analysis of Whole-Genome Sequence of African Swine Fever Virus Belgium 2018/1. Emerg. Infect Dis. 2019, 25, 1249–1252. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, R.; Zhang, X.; Li, F.; Wang, J.; Zhang, J.; Liu, X.; Wang, L.; Zhang, J.; Wu, X.; et al. Replication and Virulence in Pigs of the First African Swine Fever Virus Isolated in China. Emerg. Microbes Infect. 2019, 8, 438–447. [Google Scholar] [CrossRef]
- Mulumba-Mfumu, L.K.; Saegerman, C.; Dixon, L.K.; Madimba, K.C.; Kazadi, E.; Mukalakata, N.T.; Oura, C.A.L.; Chenais, E.; Masembe, C.; Ståhl, K.; et al. African Swine Fever: Update on Eastern, Central and Southern Africa. Transbound. Emerg. Dis. 2019, 66, 1462–1480. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Liu, J.; Wang, L.; Fan, S.; Li, Z.; Li, Y.; Yi, L.; Ding, H.; Zhao, M.; Chen, J. Current State of Global African Swine Fever Vaccine Development under the Prevalence and Transmission of ASF in China. Vaccines 2020, 8, 531. [Google Scholar] [CrossRef] [PubMed]
- MARA. Information on the Epidemic of ASF. Available online: http://www.moa.gov.cn/ztzl/fzzwfk/yqxx/ (accessed on 21 August 2020).
- Blome, S.; Gabriel, C.; Beer, M. Pathogenesis of African Swine Fever in Domestic Pigs and European Wild Boar. Virus Res. 2013, 173, 122–130. [Google Scholar] [CrossRef] [PubMed]
- OIE Official Website. Available online: https://www.oie.int/animal-health-in-the-world/information-on-aquatic-and-terrestrial-animal-diseases/ (accessed on 20 January 2021).
- Relun, A.; Grosbois, V.; Alexandrov, T.; Sánchez-Vizcaíno, J.M.; Waret-Szkuta, A.; Molia, S.; Etter, E.M.; Martínez-López, B. Prediction of Pig Trade Movements in Different European Production Systems Using Exponential Random Graph Models. Front. Vet. Sci. 2017, 4, 27. [Google Scholar] [CrossRef]
- Guinat, C.; Relun, A.; Wall, B.; Morris, A.; Dixon, L.; Pfeiffer, D.U. Exploring Pig Trade Patterns to Inform the Design of Risk-Based Disease Surveillance and Control Strategies. Sci. Rep. 2016, 6, 28429. [Google Scholar] [CrossRef]
- Wu, X.; Fan, X.; Xu, T.; Li, J. Emergency Preparedness and Response to African Swine Fever in the People’s Republic of China. Rev. Sci. Tech. 2020, 39, 591–598. [Google Scholar] [CrossRef]
- Liu, R.; Sun, Y.; Chai, Y.; Li, S.; Li, S.; Wang, L.; Su, J.; Yu, S.; Yan, J.; Gao, F.; et al. The Structural Basis of African Swine Fever Virus Pa104r Binding to DNA and Its Inhibition by Stilbene Derivatives. Proc. Natl. Acad. Sci. USA 2020, 117, 11000–11009. [Google Scholar] [CrossRef]
- Liu, J.; Liu, B.; Shan, B.; Wei, S.; An, T.; Shen, G.; Chen, Z. Prevalence of African Swine Fever in China, 2018–2019. J. Med. Virol. 2020, 92, 1023–1034. [Google Scholar] [CrossRef]
- Tao, D.; Sun, D.; Liu, Y.; Wei, S.; Yang, Z.; An, T.; Shan, F.; Chen, Z.; Liu, J. One Year of African Swine Fever Outbreak in China. Acta Trop. 2020, 211, 105602. [Google Scholar] [CrossRef]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Martínez-López, B. African Swine Fever: An Epidemiological Update. Transbound. Emerg. Dis. 2012, 59 (Suppl. 1), 27–35. [Google Scholar] [CrossRef]
- Olesen, A.S.; Lohse, L.; Boklund, A.; Halasa, T.; Belsham, G.J.; Rasmussen, T.B.; Bøtner, A. Short Time Window for Transmissibility of African Swine Fever Virus from a Contaminated Environment. Transbound. Emerg. Dis. 2018, 65, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Camós, L.; López, E.; Rodriguez, F. African Swine Fever Vaccines: A Promising Work Still in Progress. Porc. Health Manag. 2020, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.S.; Hess, W.R. Antibody Response to Inactivated Preparations of African Swine Fever Virus in Pigs. Am. J. Vet. Res. 1967, 28, 475–481. [Google Scholar] [PubMed]
- Revilla, Y.; Pérez-Núñez, D.; Richt, J.A. African Swine Fever Virus Biology and Vaccine Approaches. Adv. Virus Res. 2018, 100, 41–74. [Google Scholar] [PubMed]
- Cadenas-Fernández, E.; Sánchez-Vizcaíno, J.M.; Kosowska, A.; Rivera, B.; Mayoral-Alegre, F.; Rodríguez-Bertos, A.; Yao, J.; Bray, J.; Lokhandwala, S.; Mwangi, W.; et al. Adenovirus-Vectored African Swine Fever Virus Antigens Cocktail Is Not Protective against Virulent Arm07 Isolate in Eurasian Wild Boar. Pathogens 2020, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, G.; Giudici, S.D.; Oggiano, A. Infection, Modulation and Responses of Antigen-Presenting Cells to African Swine Fever Viruses. Virus Res. 2018, 258, 73–80. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A Seven-Gene-Deleted African Swine Fever Virus Is Safe and Effective as a Live Attenuated Vaccine in Pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef]
- Sang, H.; Miller, G.; Lokhandwala, S.; Sangewar, N.; Waghela, S.D.; Bishop, R.P.; Mwangi, W. Progress toward Development of Effective and Safe African Swine Fever Virus Vaccines. Front. Vet. Sci. 2020, 7, 84. [Google Scholar] [CrossRef]
- Sánchez, E.G.; Pérez-Núñez, D.; Revilla, Y. Development of Vaccines against African Swine Fever Virus. Virus Res. 2019, 265, 150–155. [Google Scholar] [CrossRef]
- Wang, T.; Sun, Y.; Huang, S.J.; Qiu, H.J. Multifaceted Immune Responses to African Swine Fever Virus: Implications for Vaccine Development. Vet. Microbiol. 2020, 249, 108832. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gonzalvo, F.; Rodriguez, F.; Escribano, J.M. Functional and Immunological Properties of the Baculovirus-Expressed Hemagglutinin of African Swine Fever Virus. Virology 1996, 218, 285–289. [Google Scholar] [CrossRef]
- Gómez-Puertas, P.; Rodríguez, F.; Oviedo, J.M.; Brun, A.; Alonso, C.; Escribano, J.M. The African Swine Fever Virus Proteins P54 and P30 Are Involved in Two Distinct Steps of Virus Attachment and Both Contribute to the Antibody-Mediated Protective Immune Response. Virology 1998, 243, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Barderas, M.G.; Rodriguez, F.; Gomez-Puertas, P.; Aviles, M.; Beitia, F.; Alonso, C.; Escribano, J.M. Antigenic and Immunogenic Properties of a Chimera of Two Immunodominant African Swine Fever Virus Proteins. Arch. Virol. 2001, 146, 1681–1691. [Google Scholar] [CrossRef]
- Neilan, J.G.; Zsak, L.; Lu, Z.; Burrage, T.G.; Kutish, G.F.; Rock, D.L. Neutralizing Antibodies to African Swine Fever Virus Proteins P30, P54, and P72 Are Not Sufficient for Antibody-Mediated Protection. Virology 2004, 319, 337–342. [Google Scholar] [CrossRef]
- Lopera-Madrid, J.; Osorio, J.E.; He, Y.Q.; Xiang, Z.S.; Adams, L.G.; Laughlin, R.C.; Mwangi, W.; Subramanya, S.; Neilan, J.; Brake, D.; et al. Safety and Immunogenicity of Mammalian Cell Derived and Modified Vaccinia Ankara Vectored African Swine Fever Subunit Antigens in Swine. Vet. Immunol. Immunopathol. 2017, 185, 20–33. [Google Scholar] [CrossRef]
- Argilaguet, J.M.; Perez-Martin, E.; Gallardo, C.; Salguero, F.J.; Borrego, B.; Lacasta, A.; Accensi, F.; Diaz, I.; Nofrarias, M.; Pujols, J.; et al. Enhancing DNA Immunization by Targeting ASFV Antigens to Sla-Ii Bearing Cells. Vaccine 2011, 29, 5379–5385. [Google Scholar] [CrossRef]
- Argilaguet, J.M.; Perez-Martin, E.; Nofrarias, M.; Gallardo, C.; Accensi, F.; Lacasta, A.; Mora, M.; Ballester, M.; Galindo-Cardiel, I.; Lopez-Soria, S.; et al. DNA Vaccination Partially Protects against African Swine Fever Virus Lethal Challenge in the Absence of Antibodies. PLoS ONE 2012, 7, e40942. [Google Scholar] [CrossRef]
- Lacasta, A.; Ballester, M.; Monteagudo, P.L.; Rodriguez, J.M.; Salas, M.L.; Accensi, F.; Pina-Pedrero, S.; Bensaid, A.; Argilaguet, J.; Lopez-Soria, S.; et al. Expression Library Immunization Can Confer Protection against Lethal Challenge with African Swine Fever Virus. J. Virol. 2014, 88, 13322–13332. [Google Scholar] [CrossRef]
- Argilaguet, J.M.; Perez-Martin, E.; Lopez, S.; Goethe, M.; Escribano, J.M.; Giesow, K.; Keil, G.M.; Rodriguez, F. Bacmam Immunization Partially Protects Pigs against Sublethal Challenge with African Swine Fever Virus. Antivir. Res. 2013, 98, 61–65. [Google Scholar] [CrossRef]
- Lokhandwala, S.; Waghela, S.D.; Bray, J.; Martin, C.L.; Sangewar, N.; Charendoff, C.; Shetti, R.; Ashley, C.; Chen, C.H.; Berghman, L.R.; et al. Induction of Robust Immune Responses in Swine by Using a Cocktail of Adenovirus-Vectored African Swine Fever Virus Antigens. Clin. Vaccine Immunol. 2016, 23, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.V.; Mogler, M.; Certoma, A.; Green, D.; Monaghan, P.; Williams, D.T.; Rowland, R.R.R.; Gaudreault, N.N. Evaluation of an African Swine Fever (ASF) Vaccine Strategy Incorporating Priming with an Alphavirus-Expressed Antigen Followed by Boosting with Attenuated ASF Virus. Arch. Virol. 2019, 164, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nunez, D.; Sunwoo, S.Y.; Sanchez, E.G.; Haley, N.; Garcia-Belmonte, R.; Nogal, M.; Morozov, I.; Madden, D.; Gaudreault, N.N.; Mur, L.; et al. Evaluation of a Viral DNA-Protein Immunization Strategy against African Swine Fever in Domestic Pigs. Vet. Immunol. Immunopathol. 2019, 208, 34–43. [Google Scholar] [CrossRef]
- Sunwoo, S.Y.; Perez-Nunez, D.; Morozov, I.; Sanchez, E.G.; Gaudreault, N.N.; Trujillo, J.D.; Mur, L.; Nogal, M.; Madden, D.; Urbaniak, K.; et al. DNA-Protein Vaccination Strategy Does Not Protect from Challenge with African Swine Fever Virus Armenia 2007 Strain. Vaccines 2019, 7, 12. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Chapman, D.; Hansen, D.T.; Robida, M.D.; Loskutov, A.; Craciunescu, F.; Borovkov, A.; Kibler, K.; Goatley, L.; King, K.; et al. Immunization of Pigs by DNA Prime and Recombinant Vaccinia Virus Boost to Identify and Rank African Swine Fever Virus Immunogenic and Protective Proteins. J. Virol. 2018, 92, e02219-17. [Google Scholar] [CrossRef] [PubMed]
- Leitao, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.M.E.; Portugal, F.C.; Vigario, J.D.; Martins, C.L.V. The Non-Haemadsorbing African Swine Fever Virus Isolate ASFV/Nh/P68 Provides a Model for Defining the Protective Anti-Virus Immune Response. J. Gen. Virol. 2001, 82 Pt 3, 513–523. [Google Scholar] [CrossRef] [PubMed]
- King, K.; Chapman, D.; Argilaguet, J.M.; Fishbourne, E.; Hutet, E.; Cariolet, R.; Hutchings, G.; Oura, C.A.L.; Netherton, C.L.; Moffat, K.; et al. Protection of European Domestic Pigs from Virulent African Isolates of African Swine Fever Virus by Experimental Immunisation. Vaccine 2011, 29, 4593–4600. [Google Scholar] [CrossRef]
- Sanchez-Cordon, P.J.; Chapman, D.; Jabbar, T.; Reis, A.L.; Goatley, L.; Netherton, C.L.; Taylor, G.; Montoya, M.; Dixon, L. Different Routes and Doses Influence Protection in Pigs Immunised with the Naturally Attenuated African Swine Fever Virus Isolate Ourt88/3. Antivir. Res. 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Rodze, I.; Nieto, R.; Cano-Gomez, C.; Fernandez-Pinero, J.; Arias, M. Attenuated and Non-Haemadsorbing (Non-Had) Genotype Ii African Swine Fever Virus (ASFV) Isolated in Europe, Latvia 2017. Transbound. Emerg. Dis. 2019, 66, 1399–1404. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Sanford, B.; Krug, P.W.; Carlson, J.; Pacheco, J.M.; Reese, B.; Risatti, G.R.; Gladue, D.P.; Borca, M.V. African Swine Fever Virus Georgia Isolate Harboring Deletions of 9gl and Mgf360/505 Genes Is Highly Attenuated in Swine but Does Not Confer Protection against Parental Virus Challenge. Virus Res. 2016, 221, 8–14. [Google Scholar] [CrossRef]
- Reis, A.L.; Abrams, C.C.; Goatley, L.C.; Netherton, C.; Chapman, D.G.; Sanchez-Cordon, P.; Dixon, L.K. Deletion of African Swine Fever Virus Interferon Inhibitors from the Genome of a Virulent Isolate Reduces Virulence in Domestic Pigs and Induces a Protective Response. Vaccine 2016, 34, 4698–4705. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; O’Donnell, V.; Holinka, L.G.; Ramirez-Medina, E.; Clark, B.A.; Vuono, E.A.; Berggren, K.; Alfano, M.; Carey, L.B.; Richt, J.A.; et al. The L83l Orf of African Swine Fever Virus Strain Georgia Encodes for a Non-Essential Gene That Interacts with the Host Protein Il-1beta. Virus Res. 2018, 249, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Medina, E.; Vuono, E.; O’Donnell, V.; Holinka, L.G.; Silva, E.; Rai, A.; Pruitt, S.; Carrillo, C.; Gladue, D.P.; Borca, M.V. Differential Effect of the Deletion of African Swine Fever Virus Virulence-Associated Genes in the Induction of Attenuation of the Highly Virulent Georgia Strain. Viruses 2019, 11, 599. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Nurmoja, I.; Soler, A.; Delicad, V.; Simón, A.; Martin, E.; Perez, C.; Nieto, R.; Arias, M. Evolution in Europe of African Swine Fever Genotype Ii Viruses from Highly to Moderately Virulent. Vet. Microbiol. 2018, 219, 70–79. [Google Scholar]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African Swine Fever Virus Bearing an I226r Gene Deletion Elicits Robust Immunity in Pigs to African Swine Fever. J. Virol. 2021, 95, e01199-21. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Chen, T.; Yang, J.; Yue, H.; Wang, L.; Zhou, X.; Qi, Y.; Han, X.; Ke, J.; et al. Deletion of the L7l-L11l Genes Attenuates ASFV and Induces Protection against Homologous Challenge. Viruses 2021, 13, 255. [Google Scholar]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernández-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simón, A.; et al. Assessment of African Swine Fever Diagnostic Techniques as a Response to the Epidemic Outbreaks in Eastern European Union Countries: How to Improve Surveillance and Control Programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef]
- Oura, C.A.; Edwards, L.; Batten, C.A. Virological Diagnosis of African Swine Fever—Comparative Study of Available Tests. Virus Res. 2013, 173, 150–158. [Google Scholar] [CrossRef]
- Mur, L.; Gallardo, C.; Soler, A.; Zimmermman, J.; Pelayo, V.; Nieto, R.; Sánchez-Vizcaíno, J.M.; Arias, M. Potential Use of Oral Fluid Samples for Serological Diagnosis of African Swine Fever. Vet. Microbiol. 2013, 165, 135–139. [Google Scholar] [CrossRef]
- Gallardo, C.; Fernández-Pinero, J.; Arias, M. African Swine Fever (ASF) Diagnosis, an Essential Tool in the Epidemiological Investigation. Virus Res. 2019, 271, 197676. [Google Scholar]
- Sánchez-Vizcaíno, F.; Muniesa, A.; Singleton, D.A.; Jones, P.H.; Noble, P.J.; Gaskell, R.M.; Dawson, S.; Radford, A.D. Use of Vaccines and Factors Associated with Their Uptake Variability in Dogs, Cats and Rabbits Attending a Large Sentinel Network of Veterinary Practices across Great Britain. Epidemiol. Infect. 2018, 146, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Schoder, M.E.; Tignon, M.; Linden, A.; Vervaeke, M.; Cay, A.B. Evaluation of Seven Commercial African Swine Fever Virus Detection Kits and Three Taq Polymerases on 300 Well-Characterized Field Samples. J. Virol. Methods 2020, 280, 113874. [Google Scholar] [CrossRef] [PubMed]
- Malogolovkin, A.; Sereda, A. African Swine Fever Virus Hemadsorption Inhibition Assay. Methods Mol. Biol. 2022, 2503, 159–167. [Google Scholar] [PubMed]
- Sereda, A.D.; Balyshev, V.M. Antigenic Diversity of African Swine Fever Viruses. Vopr. Virusol. 2011, 56, 38–42. [Google Scholar]







| Symptom | Peracute ASF | Acute ASF | Subacute ASF | Chronic ASF |
|---|---|---|---|---|
| Fever | High | High | Moderate | Irregular or absent |
| Thrombocytopenia | Absent | Absent or slight (late) | Transient | Absent |
| Skin | Erythema | Erythema | Erythema | Necrotic areas |
| Lymph nodes | – | Gastrohepatic and renal with marbled aspect | The majority of lymph nodes resemble a blood clot | Swollen |
| Spleen | (-) | Hyperemic splenomegaly | Partial hyperemic splenomegaly or focal infarct | Enlarged with normal color |
| Kidney | (-) | Petechial hemorrhages, mainly in cortex | Petechial hemorrhages in cortex, medulla and pelvis; perirenal oedema | (-) |
| Lung | (-) | Severe alveolar oedema | (-) | Pleuritis and pneumonia |
| Gall bladder | (-) | Petechial hemorrhages | Wall oedema | – |
| Heart | – | Hemorrhages in epicardium and endocardium | Hemorrhages in epicardium and endocardium; hydropericardium | Fibrinous pericarditis |
| Tonsils | – | – | – | Necrotic foci |
| Reproductive alteration | – | – | Abortion | Abortion |
| Cell Types | The Percent (%) of Cells | ANOVA | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 1 dpi | 2 dpi | 3 dpi | 4 dpi | 5 dpi | 6 dpi | 7 dpi | p | |
| Lymphoblasts | 0 | 0 | 0.5 ± 0.1 | 3.2 ± 0.9 ** | 3.6 ± 0.9 ** | 18.9 ± 4.8 ** | 10.3 ± 2.7 ** | 2.8 ± 0.7 ** | <0.001 |
| Small lymphocytes | 34.2 ± 7.6 | 18.6 ± 4.3 * | 25.7 ± 7.0 | 12.6 ± 3.5 * | 11.2 ± 2.9 * | 8.5 ± 2.6 * | 8.0 ± 2.0 * | 3.3 ± 1.0 * | <0.001 |
| Medium lymphocytes | 13.2 ± 3.5 | 10.4 ± 2.2 * | 10.6 ± 2.3 * | 3.8 ± 0.8 * | 7.1 ± 1.8 | 7.2 ± 2.1 | 8.4 ± 2.2 | 3.7 ± 0.8 * | <0.001 |
| Large lymphocytes | 8.5 ± 3.1 | 11.0 ± 2.1 | 6.4 ± 1.2 | 6.3 ± 1.9 | 5.3 ± 1.1 | 5.4 ± 1.2 * | 6.5 ± 1.9 | 6.5 ± 1.2 | <0.001 |
| Reactive lymphocytes | 0 | 6.1 ± 2.0 * * | 3.7 ± 0.8 ** | 3.8 ± 0.9 ** | 8.0 ± 2.0 ** | 4.5 ± 1.1 ** | 4.7 ± 1.1 ** | 14.8 ± 2.9 ** | <0.001 |
| Atypical lymphocytes | 0 | 4.3 ± 1.1 ** | 8.0 ± 1.8 ** | 7.0 ± 1.9 ** | 4.4 ± 1.0 ** | 5.4 ± 1.2 ** | 10.3 ± 2.8 ** | 14.8 ± 3.3 ** | <0.001 |
| Monoblasts | 0.5 ± 0.1 | 3.0 ± 0.4 ** | 3.7 ± 0.6 ** | 3.8 ± 1.1 ** | 7.1 ± 2.1 ** | 2.7 ± 0.8 | 2.8 ± 0.5 * | 0.2 ± 0.1 | <0.001 |
| Monocytes | 7.7 ± 2.8 | 7.3 ± 2.1 | 8.5 ± 1.3 | 10.8 ± 2.2 | 9.7 ± 2.9 | 9.0 ± 2.3 | 7.5 ± 1.6 | 7.4 ± 1.4 | <0.001 |
| Metamyelocytes | 0 | 1.2 ± 0.3 ** | 3.7 ± 0.9 ** | 3.8 ± 1.0 ** | 6.2 ± 1.4 ** | 6.3 ± 1.5 ** | 7.5 ± 1.9 ** | 7.3 ± 2.0 ** | <0.001 |
| Band neutrophils | 7.7 ± 3.0 | 12.8 ± 3.4 ** | 14.9 ± 2.5 ** | 20.3 ± 3.4 ** | 12.4 ± 3.0 | 4.5 ± 1.1 * | 5.6 ± 1.2 | 5.6 ± 1.8 | <0.001 |
| Segmented neutrophils | 23.1 ± 5.5 | 19.5 ± 4.0 * | 5.3 ± 0.9 * | 8.9 ± 1.5 * | 3.5 ± 0.9 * | 3.6 ± 0.9 * | 3.7 ± 0.8 * | 1.9 ± 0.3 * | <0.001 |
| Eosinophils | 4.9 ± 1.1 | 4.9 ± 1.3 | 1.6 ± 0.2 * | 3.8 ± 0.8 | 1.8 ± 0.5 * | 3.6 ± 0.7 | 2.8 ± 0.6 | 1.9 ± 0.2 * | <0.001 |
| Basophils | 0.2 ± 0.01 | 0.3 ± 0.02 | 0.4 ± 0.02 | 0.1 ± 0.02 | 0.3 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.1 | <0.001 |
| Plasmocytes | 0 | 0 | 1.6 ± 0.3 ** | 3.2 ± 0.5 ** | 2.7 ± 0.7 ** | 3.6 ± 0.9 ** | 4.7 ± 0.9 ** | 4.6 ± 1.1 ** | <0.001 |
| Dead cells | 0 | 0.6 ± 0.1 | 5.3 ± 1.0 ** | 8.9 ± 1.6 ** | 16.8 ± 3.4 ** | 16.2 ± 4.0 ** | 16.8 ± 3.8 ** | 24.8 ± 4.8 ** | <0.001 |
| Cellular Factors | Inhibitors | Reference |
|---|---|---|
| Na+/H+ channels | EIPA | [70] |
| Actin | Cytochalasin D and B | [71] |
| Latrunculin A | ||
| Myosin II | Blebbistatin | [71] |
| EGFR | 324,674 | [71] |
| PI3K | LY294002Worthmanin | [66] |
| Rac1 | NSC23766, Rac1-N17 | [72] |
| Pak1 | IPA-3, Pak1-AID | [70] |
| Tyrosin kinases | Genistein | [71] |
| Dynamin-2 | Dynasore | [66] |
| Clathrin | Clorpromazine | [71] |
| Microtubules | Nocodazol | [71] |
| Vacuolar acidification | Cloroquine, NH4Cl, | [66] |
| Bafilomycin A | ||
| Cholesterol | MβCD | [73] |
| Rab-7 | Rab-7-T22N | [74] |
| Rab-7 siRNA | ||
| CD163 | CD163siRNA | [70] |
| CD45 | CD45siRNA | [73] |
| CD203a | CD203asiRNA | [74] |
| CD163 | CD163siRNA | [70] |
| Gene Name | Functional Group | Reference |
|---|---|---|
| A104R | Genome Organization | [102] |
| A151R | Structural/viral morphology | [103] |
| A179L | Immune evasion | [104] |
| A238L | Immune evasion | [105] |
| A489R | MGF 505 | [106] |
| A505R | MGF 505 | [107] |
| A506R | MGF 505 | [107] |
| A528R | MGF 505 | [106] |
| A542R | MGF 505 | [108] |
| B119L | Other/enzyme | [109] |
| B125R | Transcription/RNA modification | [110] |
| B263R | Transcription/RNA modification | [111] |
| B318L | Other/enzyme | [112] |
| B438L | Structural/viral morphology | [42] |
| B602L | Structural/viral morphology | [113] |
| B646L | Structural/viral morphology | [114] |
| B962L | NA metabolism/DNA replication/repair | [115] |
| C257L | TR/PSP | [116] |
| C962R | NA metabolism/DNA replication/repair | [117] |
| CP204L | Other/enzyme | [118] |
| CP2475L | Structural/viral morphology | [45] |
| CP530R | Structural/viral morphology | [119] |
| D1133L | Transcription/RNA modification | [120] |
| D117L | Structural/viral morphology | [121] |
| D250R | Transcription/RNA modification | [122] |
| D339L | Transcription/RNA modification | [120] |
| E165R | NA metabolism/DNA replication/repair | [123] |
| E183L | Structural/viral morphology | [124] |
| E184L | TR/PSP | [125] |
| E199L | Structural/viral morphology | [126] |
| EP153R | Immune evasion | [127] |
| EP402R | Immune evasion | [128] |
| I196L | TR/PSP | [129] |
| K78R | Structural/viral morphology | [130] |
| L57L | Uncharacterized | [42] |
| M448R | Other/enzyme | [131] |
| O61R | Structural/viral morphology | [116] |
| QP383R | Other/enzyme | [132] |
| A240L | Thymidylate kinase | [133] |
| K196R | Thymidine kinase | [134] |
| F334L | Ribonucleotide reductase (small subunit) | [44] |
| F778R | Ribonucleotide reductase (large subunit) | [44] |
| G1211R | DNA polymerase family B | [44] |
| P1192R | DNA topoisomerase type II | [135] |
| E301R | Proliferating cell nuclear antigen (PCNA)-like | [44] |
| O174L | DNA polymerase X-like | [136] |
| NP419L | DNA ligase | [137] |
| E296R | AP endonuclease class II | [138] |
| EP1242L | RNA polymerase subunit 2 | [139] |
| C147L | RNA polymerase subunit 6 | [108] |
| NP1450L | RNA polymerase subunit 1 | [139] |
| H359L | RNA polymerase subunit 3 | [44] |
| D205R | RNA polymerase subunit 5 | [44] |
| CP80R | RNA polymerase subunit 10 | [44] |
| C315R | TFIIB-like | [140] |
| A859L | Helicase superfamily II | [141] |
| F1055L | Helicase superfamily II | [44] |
| D1133L | Helicase superfamily II | [120] |
| Q706L | Helicase superfamily II | [142] |
| QP509L | Helicase superfamily II | [143] |
| I243L | Transcription factor SII | [144] |
| NP868R | Guanylyl transferase | [145] |
| C475L | PolyA polymerase large subunit | [44] |
| EP424R | FTS J-like methyl transferase domain | [44] |
| EP364R | ERCC4 nuclease domain | [146] |
| D345L | Lambda-like exonuclease | [44] |
| B385R | VV A2L-like transcription factor | [44] |
| G1340L | VV A8L-like transcription factor | [147] |
| B175L | VV VLTF2-like late transcription factor, FCS-like finger | [44] |
| C962R | DNA primase | [117] |
| R298L | Serine protein kinase | [148] |
| I215L | Ubiquitin conjugating enzyme | [149] |
| D250R | Nudix hydrolase | [122] |
| A224L | IAP apoptosis inhibitor | [90] |
| DP71L | Similar to HSV ICP34.5 neurovirulence factor | [150] |
| KP177R | P22 | [151] |
| A137R | P11.5 | [152] |
| A78R | P10 | [44] |
| B646L | P72 major capsid protein; involved in virus entry | [153] |
| B438L | P49; required for formation of vertices in icosahedral capsid | [154] |
| B602L | Chaperone; involved in folding of capsid; not incorporated into virions | [155] |
| B119L | ERV 1-like; involved in redox metabolism | [156] |
| S273R | SUMO-1-like protease; involved in polyprotein cleavage | [157] |
| CP2475L | pp220 polyprotein precursor of p150, p37, p14, and p34; required for packaging of nucleoprotein core | [45] |
| H108R | J5R; transmembrane domain | [158] |
| E120R | P14.5; DNA-binding; required for movement of virions to plasma membrane | [159] |
| E248R | E248R (k2R); possible component of redox pathway required disulphide bond formation | [160] |
| MGF 110-4L (XP124L) | XP124L; multigene family 110 member | [44] |
| Immune Response | Viral Genes | Immune Elements and Mechanisms | Impact on Virulence | Reference |
|---|---|---|---|---|
| Type I interferon response | A276R | Dampening type I IFN response by regulating IRF3 | ND | [163] |
| Inflammatory response | A528R | Promoting the expression of ULK1 to degrade STING | Attenuated | [163] |
| MGF360-12L | Interacting with nuclear transport proteins importin α (KPNA2, KPNA3, and KPNA4) to disrupt NF-κB nuclear translocation | ND | [171] | |
| I329L | Inhibiting the crucial adaptor protein TRIF | ND | [172] | |
| DP96R | Degradation of TBK1 | Attenuated | [164] | |
| L83L | No reduction in virulence | No reduction in virulence | [167] | |
| A238L | Inhibiting the activation of the NF-κB pathway | No reduction in virulence | [161] | |
| Apoptosis | 226L, A151R, NP419L, QP383R | ND | ND | [137] |
| A179L | Bind to pro-apoptotic proteins (Bid, Bim, Bak, and Bax) to inhibit apoptosis | ND | [104] | |
| A224L | Activating NF-κB pathway to promote anti-apoptotic genes expression, e.g., IAP and Bcl-2 family proteins | No reduction in virulence | [173] | |
| EP153R | Inhibiting the expression of caspase-3 | ND | [174] | |
| E183L | Interacting with DLC8 to activate caspase-3 and caspase-9 | ND | [175] | |
| DP71L | Recruit host phosphatase 1 (PP1) and remove the phosphorylation of eIF-2α to restore cellular protein synthesis to block CHOP activation suppressing apoptosis | No reduction in virulence | [176] | |
| A238L | Inhibiting CaN to decrease apoptosis; NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1 and c-IAP2 to suppress caspase-8 activation | No reduction in virulence | [177] |
| Action | Resistance |
|---|---|
| Temperature | Highly resistant to low temperatures. Heat-inactivated by 56 °C/70 min; 60 °C/20 min. |
| pH | Inactivated by pH < 3.9 or >11.5 in serum-free medium. Serum increases the resistance of the virus, e.g., at pH 13.4 resistance lasts up to 21 h without serum, and 7 days with serum. |
| Chemicals/disinfectants | Susceptible to ether and chloroform. Inactivated by 8/1000 sodium hydroxide (30 min), hypochlorites as 2.3% chlorine (3 min), 3/1000 formalin (30 min), 3% orthophenylphenol (30 min) and iodine compounds. |
| Survival | Remains viable for long periods in blood, feces, and tissues, especially infected uncooked or undercooked pork products. Can multiply in vectors (Ornithodoros sp.). |
| Aldehydes | Mutually Bind to Proteins, Inhibit Transport Mechanisms |
|---|---|
| Halogens (hypochlorite, iodophors, ClO2) | Penetrates the membrane and oxidizes proteins, interrupts the cell’s oxidative phosphorylation |
| Peroxides | Penetrates the membrane and oxidizes lipids, proteins, and DNA |
| Phenolics | Poisons the protoplasm and damages the cellular membrane |
| Quaternary ammonium compounds (QUATs) | Damages the cellular membrane and disrupts the membrane, cytoplasmic potential, and pH gradient |
| Serogroup | ASFV Reference Strain | ASFV Strains and Isolates | |
|---|---|---|---|
| Highly/Moderately Virulent | Low Virulent Attenuated/Avirulent | ||
| 1 | Lisbon-57 | Lisbon-57 (L-57), Kimakia, Katanga-78, Katanga-105, Katanga-115, Madeira-65, Diamant | LS, L-50, LF-97, Kimakia-155, Diamant-160, Katanga-139 (Kc-139), Katanga-149 (Kc-149), Katanga-160 (Kc-160), LK-111, Katanga-350 |
| 2 | Congo-49 | Congo-49 (K-49), Yamba-74, Le Bry-73, Sylva | NVL-1, Mfuati-79, Ndjassi-77, KK-202, KK-262/C |
| 3 | Mozambique-78 | Mozambique-78 (M-78), MK-101 | MK-200, MK-210 |
| 4 | France-32 | France-32 (F-32), Cuba-71, Brazil-80, Cuba-80, Malta-78, Sao-Tome, and Principe-79 (STP-1), DNOPA-Luanda, Odessa-77 | FK-32/135 |
| 5 | TSP-80 | TSP-80 | TSP-80/300 |
| 6 | TS-7 | TS-7 | TS-7/150, TS-7/230 |
| 7 | Uganda | Uganda | UK-50, UK-80 |
| 8 | Rhodesia, Stavropol 01/08 | Rhodesia, Stavropol 01/08 | St-CV1/20 |
| 9 | Davis | Davis | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; et al. African Swine Fever Virus: A Review. Life 2022, 12, 1255. https://doi.org/10.3390/life12081255
Li Z, Chen W, Qiu Z, Li Y, Fan J, Wu K, Li X, Zhao M, Ding H, Fan S, et al. African Swine Fever Virus: A Review. Life. 2022; 12(8):1255. https://doi.org/10.3390/life12081255
Chicago/Turabian StyleLi, Zhaoyao, Wenxian Chen, Zilong Qiu, Yuwan Li, Jindai Fan, Keke Wu, Xiaowen Li, Mingqiu Zhao, Hongxing Ding, Shuangqi Fan, and et al. 2022. "African Swine Fever Virus: A Review" Life 12, no. 8: 1255. https://doi.org/10.3390/life12081255
APA StyleLi, Z., Chen, W., Qiu, Z., Li, Y., Fan, J., Wu, K., Li, X., Zhao, M., Ding, H., Fan, S., & Chen, J. (2022). African Swine Fever Virus: A Review. Life, 12(8), 1255. https://doi.org/10.3390/life12081255